溫室黃瓜連作土壤真菌多樣性變化分析
2022-04-08 07:39:20孫玉祿高曉梅劉曉輝桓明輝池景良
中國土壤與肥料 2022年2期
李 楊,敖 靜,孫玉祿,高曉梅,劉曉輝,桓明輝,池景良
(遼寧省微生物科學研究院,遼寧省微生物菌種選育重點實驗室,遼寧 朝陽 122000)
黃瓜(Cucumis Sativas L.)是我國設施園藝生產的主要蔬菜作物,栽培面積占蔬菜總面積的60%,隨著設施栽種面積的逐年增加,為追求高產超量施用農藥和化肥,造成土壤理化條件改變、健康狀況惡化、病蟲害增多、產量下降、連作障礙嚴重,嚴重影響設施農業的可持續發展,同時食品安全也威脅著消費者的健康,因此黃瓜連作障礙問題越來越多地引發人們關注[1]。
土壤微生物是農業生態系統中的重要組成部分,土壤真菌在土壤中的數量僅次于細菌,承擔著土壤分解者的角色,在物質循環和能量循環過程中發揮著重要作用,可以將復雜難降解的有機質轉化為植物可以利用的營養元素[2-5]。有研究表明,作物連作栽培定向地影響土壤微生物的生長繁殖,細菌和有益微生物數量減少,病原真菌數量增加[6],導致土壤根際微生態系統失衡,是土壤連作障礙的主要原因[7-8]。
傳統研究方法應用于微生物群落的多樣性已取得較多的科研成果,但由于土壤中絕大部分微生物是不可培養的,傳統的分離鑒定方法只能區分上述種類中的0.1%~10%,導致對環境微生物群落多樣性的認識受到限制[9-11]。隨著分子生物學技術的突破,新一代高通量測序技術為研究微生物豐度、組成、群落結構、遺傳進化、代謝途徑等提供大量的信息,為大規模、快速、準確、全面檢測土壤微生物多樣性提供了技術保障[12-13],使微生物和連作障礙之間的關系再次成為研究的熱點。本試驗采用Illumina MiSeq 高通量測序技術,以日光溫室黃瓜連作不同年限土壤為研究對象,分析溫室黃瓜不同連作年限根際土壤樣品真菌種群結構和多樣性的變化,獲取了真菌在門、綱、目、科、屬不同分類水平上的相對豐度、群落結構和遺傳信息,探索了連作模式下土壤真菌中有益和有害菌群消長和演替的響應,闡明了連作年限-土壤微生物群落結構-環境因子之間的相互關系,為克服連作障礙提供理論依據,助力設施農業可持續發展。
1 材料與方法
1.1 試驗地概況及試驗設計
本試驗樣品來自遼寧省微生物科學研究院喀左平房子鎮設施蔬菜科研基地,位于119°38′39′E、40°58′44′′N,海拔310 m。試驗設4 個處理,根據種植年限分別命名為連作1 年(F1)、3 年(F3)、5 年(F5)和7 年(F7),每個處理設3 次重復,編號分別為F1A、F1B、F1C;F3A、F3B、F3C;F5A、F5B、F5C;F7A、F7B、F7C,共12 個樣品,田間管理措施相同。
1.2 樣品采集
在黃瓜種植后期,去除表層砂石和腐葉,每個處理在深度為20 ~30 cm 處采集3 份土壤,在每個區采用五點取樣法取樣,混勻成1 份裝進自封袋放入冰盒帶回實驗室。一部分鮮土放入超低溫冰箱,用于微生物多樣性研究;另一部分鮮土放入-4℃冰箱保存,用于測定可培養微生物數量和酶活性;還有一部分土壤自然風干后過1 mm 篩,用于測定土壤基本理化性質。
1.3 土壤基本理化性質測定
采用ThermoA211 酸度計按水土比5∶1 測定土壤pH;采用ORIONTDS 型電導率儀測定EC 值;采用堿解擴散法測定堿解氮;采用鹽酸-硫酸混合液浸提,北京吉天全自動流動注射分析儀iFIA7 測定土壤有效磷;采用醋酸銨浸提-火焰光度法測定速效鉀。
1.4 土壤微生物總DNA 的提取
采用Omega soil DNA kit 試劑盒,稱取0.5 g 鮮土,根據說明按步驟提取12 個土壤樣品中的微生物基因組DNA,0.8%的瓊脂糖凝膠電泳檢測DNA質量并通過紫外分光光度計對DNA 進行定量。
1.5 真菌ITS1 區PCR 擴增及測序
采 用 特 異 性 引 物ITS5F:5’-GGAAGTAAAA GTCGTAACAAGG-3’、ITS1R:5’-GCTGCGTTC TTCATCGATGC-3’對 真 菌ITS rRNA 的ITS1 區 進行PCR 擴增。PCR 反應體系為Q5 DNA 聚合酶0.25 μL,5×擴增緩沖液5 μL,5×GC 緩沖液5 μL,10 mmol/L dNTP 0.5 μL,模板DNA 1 μL,10 μmol/L正向、反向引物各1 μL,水11.25 μL。PCR 擴增條件為:98℃預變性2 min;98℃15 s,50℃30 s,72℃30 s,循環25 ~27 次;72℃延伸5 min。PCR產物采用2%瓊脂糖凝膠電泳檢測,用Axy Prep DNA 凝膠回收試劑盒回收目的片段,樣品送上海派森諾生物科技有限公司進行Illumina MiSeq 高通量測序。
1.6 序列獲取與分析
為獲得更為精準的生物學信息,首先使用Fast QC 進行原始數據的質量控制,對測序原始數據進行拼接、過濾,獲得最終用于分析的序列。使用Usearch 在0.97 的相似度下進行去冗余、聚類和操作分類單元OTU 表格生成。采用Graphpad Prism 8 one-way ANOVA 方法進行差異顯著性分析。利用Mothur[14]計算真菌群落的α 多樣性,并對Shannon 指數和Chao1 指數進行分析。基于Bray_Curtis距離矩陣,采用主坐標分析(PCoA)進行真菌群落的群落結構和聚類分析。利用FUNGuild 數據庫分析真菌的功能類別,進行功能預測和分類[15]。選擇Pearson 分析土壤真菌門相對豐度與土壤理化性質的相關性,采用Mantel 檢驗分析真菌整體群落與土壤理化性質之間的相關性。
2 結果與分析
2.1 溫室黃瓜連作不同年限根際土壤真菌相對豐度和群落組成的變化
通過Illumina Miseq 高通量測序分析,從12個樣品中共獲得786148 條優化序列,每樣本為46932 ~91804 條優化序列(平均65512 條優化序列),213009495 優化堿基數,優化序列長度50 ~485 bp,平均長度270 bp(表1),共檢測到14 個門,42 個綱,90 個目,197 個科,318 個屬和1205 個OTU。連作不同年限后真菌群落組成在門和綱分類水平上發生變化(圖1、2)。在門水平上,子囊菌門(Ascomycota)為優勢菌門,相對豐度占55.74% ~77.78%,其次是未被歸類的真菌(8.04% ~23.38%)、擔子菌門(Basidiomycota)(3.59% ~19.63%) 和Mortierellomycota(0.20% ~6.49%), 壺 菌 門(Chytridiomycota)(0.08% ~1.82%)、Aphelidiomycota(0.01%~1.57%)和Rozellomycota(0.51%~1.12%)相對豐度占比很少,且在F1 中并未發現Rozellomycota 真菌門(圖1a)。日光溫室黃瓜連作不同年限后真菌在門水平上的群落組成變化明顯,子囊菌門相對豐度在F3 樣品中無明顯變化,在F5 和F7 樣品中相對豐度分別下降9.94%和28.24%,而擔子菌門相對豐度在F3 樣品中顯著降低后又逐年上升。
在綱水平上,糞殼菌綱(Sordariomycetes,28.01% ~68.01%)、c_unclassified_k_Fungi(8.04% ~23.38%)、散囊菌綱(Eurotiomycetes,1.75% ~22.81%)、 傘 菌 綱(Agaricomycetes,1.13%~13.64%)和座囊菌綱(Dothideomycetes,2.27%~10.35%)的相對豐度較高,其次是c_unclassified_p_Ascomycota(0.18% ~6.15%)、 銀耳綱(Tremellomycetes,1.05% ~4.40%)、被孢霉 綱(Mortierellomycetes,0.20% ~6.12%)、盤菌綱(Pezizomycetes,0.13%~5.0%)、酵母菌綱(Saccharomycetes,0.09% ~1.88%) 等( 圖1b)。糞殼菌綱相對豐度變化顯著,連作3 年樣本中相對豐度最高,達68.01%,之后持續下降,到連作7年時剩余28.01%。
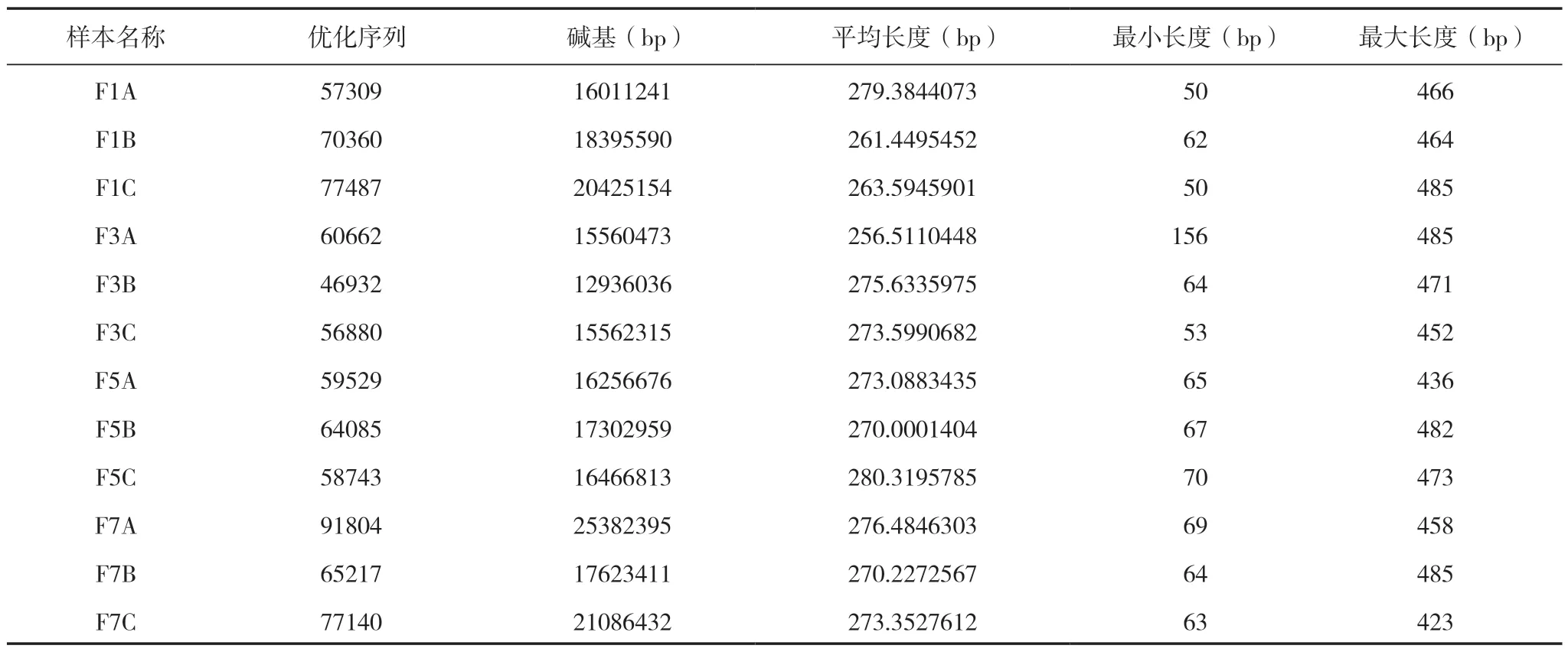
表1 真菌ITS1 Illumina MiSeq 高通量測序結果
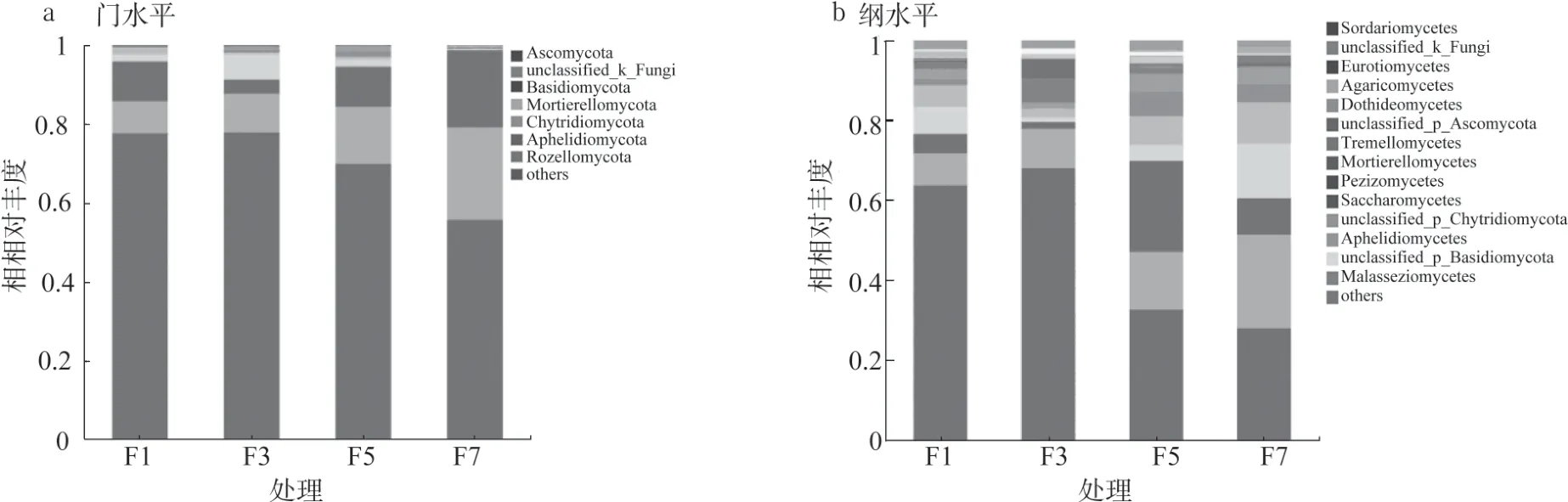
圖1 黃瓜連作條件下土壤真菌在門和綱分類水平上的群落組成
LEfSe 分析顯示,在屬分類水平上,設施黃瓜連作1 年后起重要作用的屬有Ramophialophora、錐蓋傘屬(Conocybe)、裂殼菌屬(Schizothecium)、柄孢殼菌屬(Podospora)和微囊菌屬(Microascus),連作3 年后起重要作用的屬有糞盤菌屬(Ascobolus)、蠟蚧菌屬(Lecanicillium)和被孢霉屬(Mortierella),連作5 年后起重要作用的屬有Pseudopithomyces、擬青霉屬(Simplicillium)和嗜熱真菌屬(Thermomyces),連作7 年后起重要作用的屬有曲霉屬(Aspergillus)和栓菌屬(Trametes)(圖2)。
2.2 不同連作年限對溫室黃瓜根際土壤真菌群落多樣性的影響
Chao1 指數主要是計算群落豐度,用于估計群落樣品中包含的物種總數,Chao1 指數越大,微生物群落物種數越大。Shannon 指數主要用于計算群落多樣性,該指數受群落中物種豐富度和均勻度的共同影響,Shannon 指數越大,微生物群落的多樣性越高。日光溫室黃瓜長期連作增加了土壤中真菌的多樣性,Shannon 指數隨著連作年限的增加逐漸增高,但差異不顯著(圖3a);Chao1 指數在連作3年達到最高,5 年后明顯降低(圖3b),且差異顯著(P<0.05)。真菌群落的多樣性為F7>F5>F3>F1,而真菌總數為F3>F1>F7>F5。

圖2 Lefse 分析連作不同年限真菌組成差異
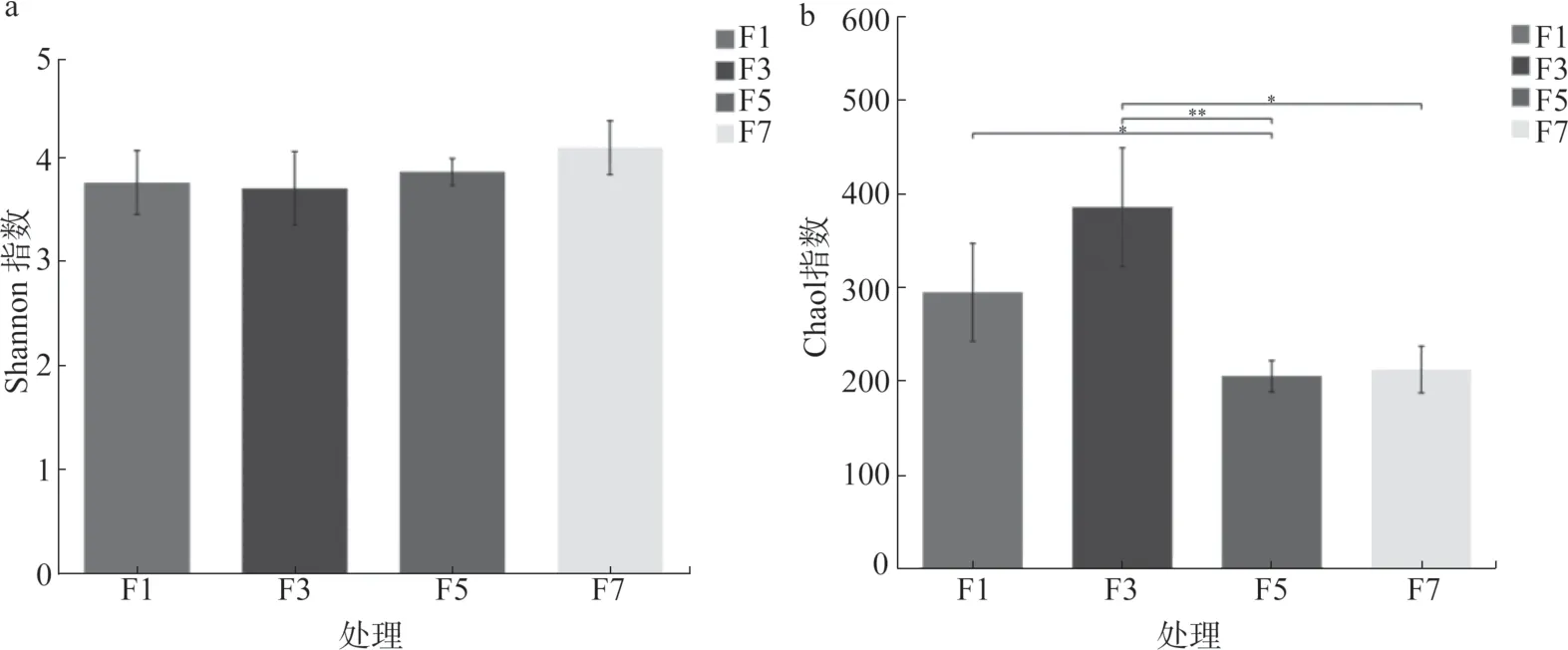
圖3 設施黃瓜不同連作年限土壤真菌多樣性指數組間差異檢驗
對黃瓜連作不同年限的真菌群落整體進行主坐標分析(PCoA),結果(圖4)顯示黃瓜連作年限可以影響真菌群落結構。不同連作年限黃瓜根際真菌群落結構均差異極顯著(P<0.01),在PCoA 上4種連作年限土壤樣品分開,相同年限土壤樣品聚在一起。基于Bray-Curtis 的ANOSIM 進一步分析表明,在不同連作年限之間的真菌群落結構的差異明顯高于組內差異(圖5)。
2.3 溫室黃瓜不同連作年限對土壤功能真菌的影響
采用FUNGuild 對不同連作年限真菌群落進行功能預測,4 組樣品共檢測到1205 個OTU,其中479 個OTU 被劃分為8 個真菌功能類群,約占總OTU 數量的39.75%。從圖6 中可以看出,腐生菌(Saprotroph)是主要功能類群,包括240 個OTU,占總OTU 數量的19.92%。然后為植物病原菌/腐生/共生過渡型菌(Pathotroph-Saprotroph-Symbiotroph),包括75 個OTU,約占6.22%;植物病原菌(Pathotroph), 包 括50 個OTU, 約 占4.15%;腐生/共生過渡型菌(Saprotroph-Symbiotroph),包括40 個OTU,約占3.32%;植物病原菌/腐生菌過渡型(Pathotroph-Saprotroph),包括36 個OTU,約占2.99%。共生菌(Symbiotroph)僅有19 個OTU,占1.58%;植物病原菌/共生過渡型菌(Pathotroph-Symbiotroph),12 個OTU,約占1.0%;植物病原/腐生/共生過渡型菌(Pathogen-Saprotroph-Symbiotroph)只有7 個OTU,所占比例為0.58%。本研究中仍有60.25%的OTU 不能進行功能分類,需要進一步進行分析。
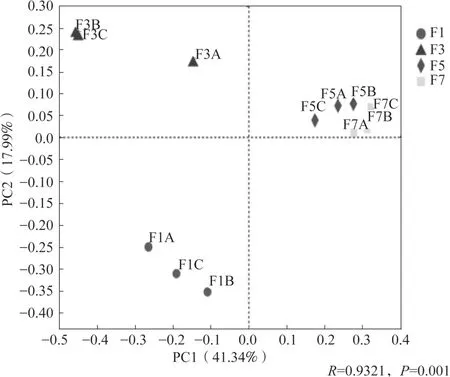
圖4 基于屬水平的Bray-Curtis 距離矩陣不同連作年限土壤真菌群落的主坐標分析
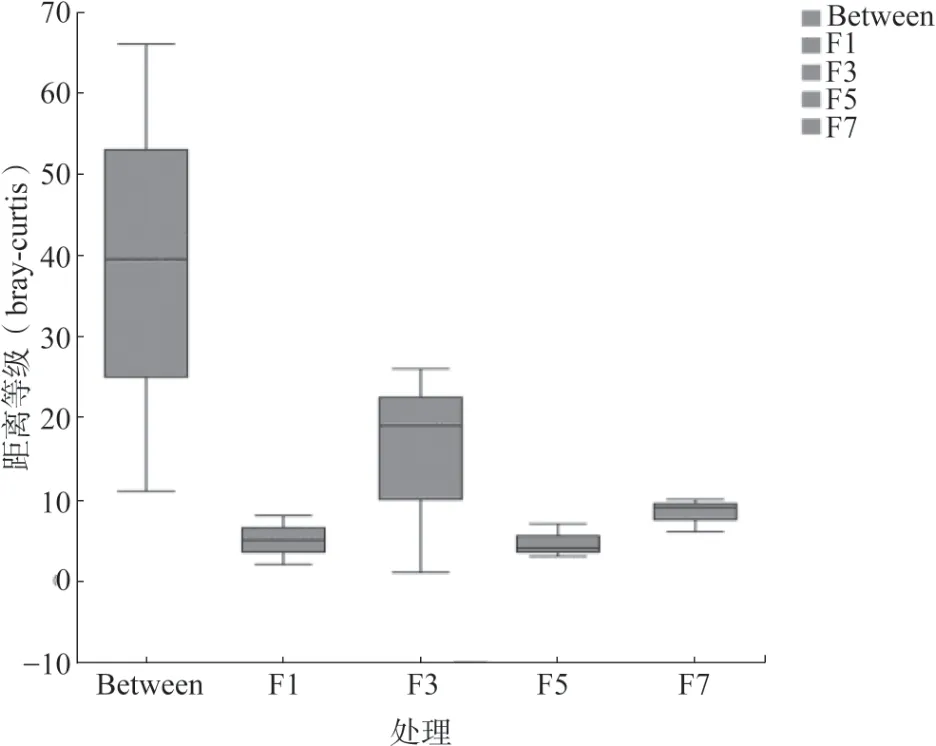
圖5 基于Bray-Curtis 距離矩陣ANOSIM 差異分析
從圖6 可以看出,設施黃瓜連作不同年限根際土壤中真菌功能類群的相對豐度不同。其中,連作3年(F3)根際土壤中的腐生/共生過渡型菌(6.76%)和植物病原菌/腐生/共生過渡型菌(44.75%)的相對豐度明顯高于F1、F5 和F7(0.94%~2.42%、12.15%~14.76%),連作5 年土壤中植物病原菌/腐生/共生過渡型菌(0.91%)、植物病原菌/共生過渡型菌(18.22%)和植物病原菌(8.70%)的相對豐度最高,而共生菌(1.69%)和腐生菌(35.45%)的相對豐度在連作7 年樣品中最高。但是仍有很大一部分真菌不能進行功能歸類(Unassigned)。
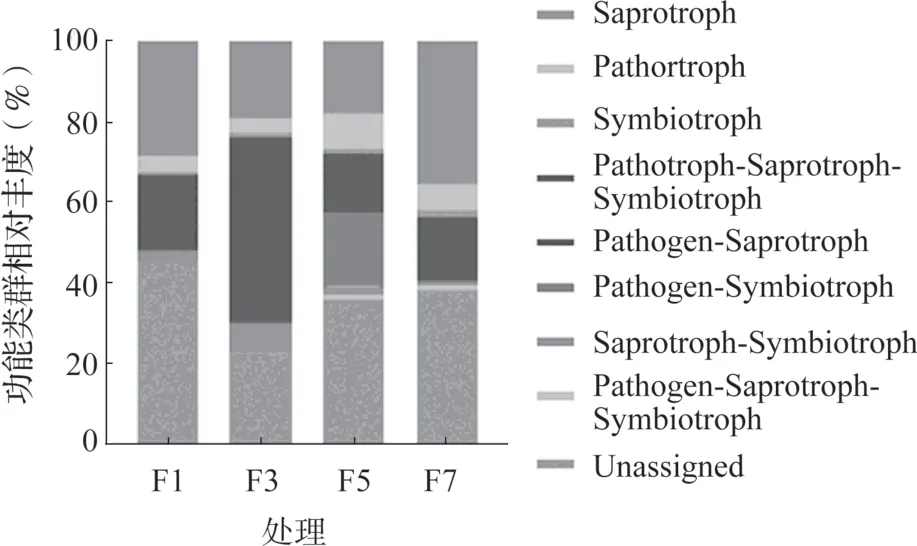
圖6 不同連作年限黃瓜根際真菌菌群功能組成與豐度
2.4 土壤真菌群落與土壤理化性質間的相關分析
土壤環境因子pH、堿解氮(AN)、脲酶(SUR)等與土壤真菌群落結構之間存在密切關系。如圖7所示,土壤pH 與Aphelidiomycota(R=-0.651)和毛霉門(Mucoromycota)(R=-0.704)呈顯著負相關關系(P<0.05)。土 壤AN 和SUR 與Mortierellomycota(R=0.741,R=0.713)呈極顯著正相關關系(P<0.01),與梳霉門(Kickxellomycota)(R=0.65,R=0.591)呈顯著正相關關系(P<0.05),與擔子菌門(Basidiomycota)(R=-0.762,R=-0.762)呈極顯著負相關關系(P<0.01);土壤AN 與油壺菌門(Olpidiomycota)(R=0.708)呈極顯著正相關關系(P<0.01),SUR 與捕蟲霉門(Zoopagomycota)(R=0.78)呈極顯著正相關關系(P<0.01),與油壺菌門(Olpidiomycota)(R=0.658)和壺菌門(Chytridiomycota)(R=0.58)呈顯著正相關關系(P<0.05)。 土 壤 有 效 磷(AP) 與 隱 真 菌 門(Rozellomycota)(R=0.599)呈顯著正相關關系(P<0.05),與蛙糞霉門(Basidiobolomycota)(R=-0.640)和芽枝霉門(Blastocladiomycota)(R=-0.592)呈顯著負相關關系(P<0.05)。土壤速效鉀(AK)與總豐度前14 的真菌門均無顯著相關性。環境因子聚類分析表明,AN和SUR 聚為一類,AP 和AK 聚為一類。
采用Mantel 檢驗設施黃瓜連作不同年限土壤真菌群落結構和土壤理化性質之間的相關性(表2),可以看出,土壤真菌群落與pH、AN、SUR 和AP具有顯著(P<0.05)或極顯著(P<0.01)相關性,對真菌菌群影響表現為SUR>AP>AN>pH。
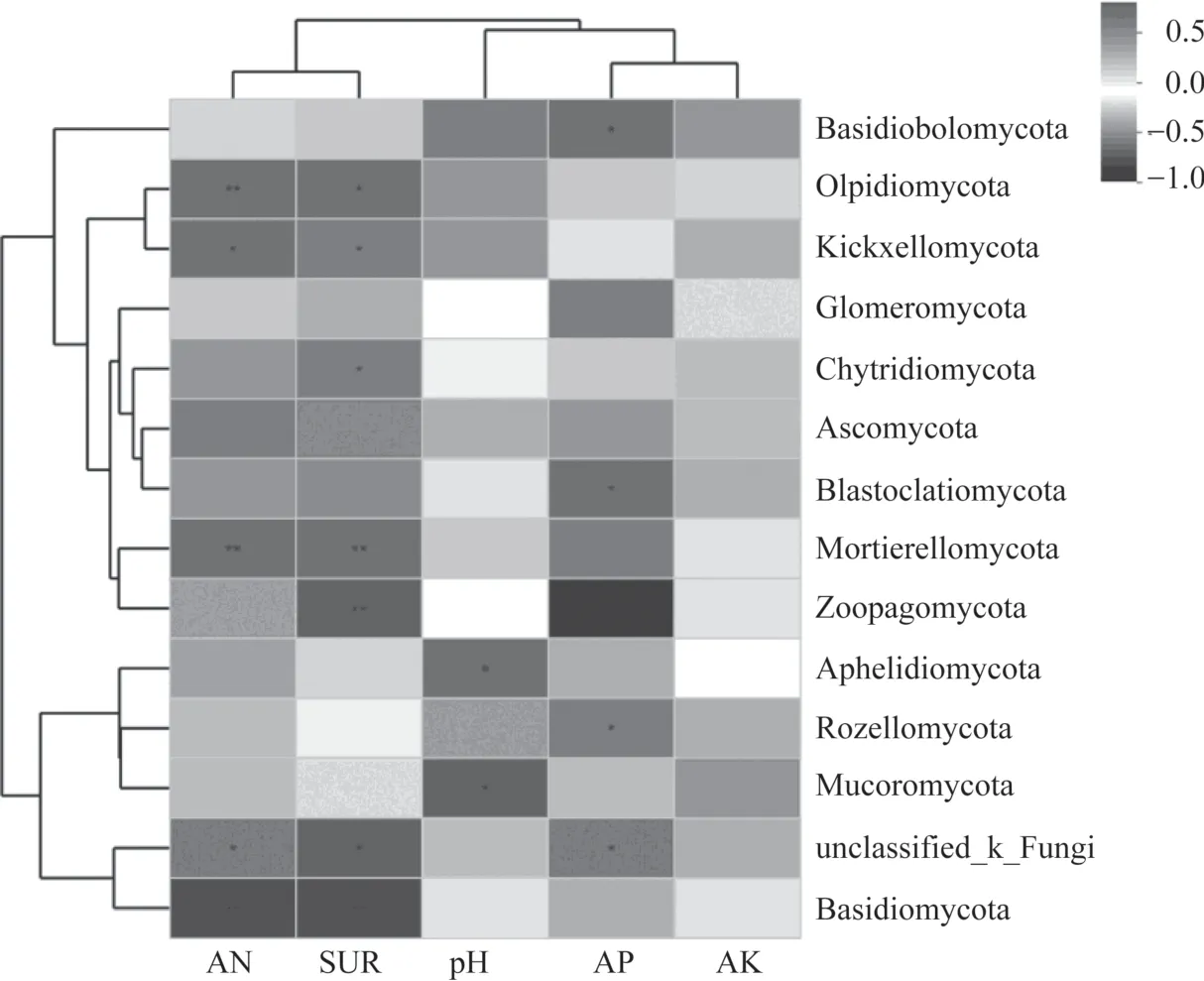
圖7 總豐度前14 真菌門與土壤理化性質及酶活性相關性熱圖
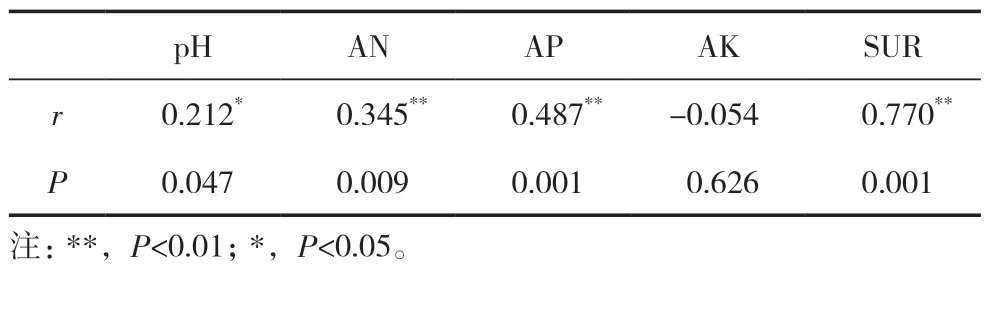
表2 溫室黃瓜不同連作年限土壤真菌群落與土壤理化性質的Mantel 檢驗
3 結論與討論
作物長期連作會導致土壤理化性質改變,土壤酶活性降低,土壤微生物總量、群落結構及多樣性降低,從而導致其產量和品質降低[16-17]。真菌對土壤有機質的分解起著重要的作用,動植物殘體的分解主要依靠腐生真菌來進行,土壤真菌的多少與土壤有機質含量密切相關[18]。長期集約化的溫室連作栽培導致病害增多,尤其真菌型病害更為普遍[19-20]。隨著新一代高通量測序技術的發展,關于土壤微生物群落和多樣性的研究越來越多集中在連作與微生物群落的關系上。
本研究中,溫室黃瓜連作不同年限后真菌群落組成在門和綱分類水平上發生變化。在門水平上,子囊菌門(Ascomycota)為優勢菌門,相對豐度占55.74%~77.78%,其次是未被歸類的真菌、擔子菌門(Basidiomycota)和Mortierellomycota,壺菌門(Chytridiomycota)、Aphelidiomycota 和Rozellomycota 相對豐度占比很少,且在F1 中并未發現Rozellomycota 真菌門。日光溫室黃瓜連作不同年限后真菌在門水平上的群落組成變化明顯,子囊菌門相對豐度在F3 樣品中無明顯變化,在F5 和F7 樣品中相對豐度分別下降9.94%和28.24%,而擔子菌門相對豐度在F3 樣品中顯著降低后又逐年上升。隨著連作年限的增加,微生物菌群的改變和豐度的變化會擾亂土壤微生態的平衡,更嚴重的會導致植物的死亡[21]。Schoch 等[22]研究表明,子囊菌門在根際土壤有機物的降解中起著重要作用,推測子囊菌門相對豐度的降低可能與土壤有機質和土壤肥力下降有關。
設施黃瓜長期連作增加了土壤中真菌的多樣性,黃瓜連作后Shannon 指數隨著連作年限的增加逐漸增高,但差異不顯著,說明土壤樣本的群落多樣性逐漸升高;估計物種總數的Chao1 指數呈現先上升后下降的趨勢且差異顯著,表明連作5 和7 年土壤中真菌總數都相對較少,這一結果與不同連作年限懷牛膝的土壤微生物多樣性的研究結果相一致[23]。馮翠娥等[24]同樣在研究硒砂瓜連作對土壤真菌群落結構的影響時得到了一致的結果。這可能是由于隨著連作年限的延長,長期高強度、重復的農業耕作破壞了土壤穩定結構,降低了土壤中微生物的豐度和均勻度,使得土壤由“細菌型”向“真菌型”轉化,土壤中細菌群落多樣性降低,真菌群落多樣性增高。
研究表明,腐生菌參與有機物降解,植物病原菌/腐生/共生過渡型容易受到人活動的影響從而進行功能轉換[25-27]。在本研究中,設施黃瓜連作不同年限根際土壤中真菌功能類群的相對豐度不同,腐生菌和植物病原菌/腐生/共生過渡型菌是黃瓜根際中兩個主要功能類群。其中,腐生/共生過渡型菌(6.76%)和植物病原菌/腐生/共生過渡型菌(44.75%)兩種過渡型在連作3 年土壤中相對豐度最高,植物病原菌/共生過渡型菌(18.22%)和植物病原菌/腐生/共生過渡型菌(0.91%)兩種過渡型在連作5 年土壤中相對豐度最高,表明黃瓜根際土壤在連作3 和5 年逐漸過渡為共生菌(1.69%)和腐生菌(35.45%)型,其相對豐度在連作7 年樣品中最高,植物病原菌(8.70%)的相對豐度在連作5 年達最高。本研究有50.24%的OTU 在數據庫中無法進行功能分類,需要采用其他手段進一步研究。
在連作條件下,土壤理化性質可以有效地體現土壤的健康狀況,影響土壤微生物種群數量及分布,土壤堿解氮、有效磷含量對土壤微生物區系從細菌型向真菌型轉化和病害發生率都起著至關重要的作用[28],pH 是影響土壤真菌的主要環境因子[29],不同植被的營養代謝活動導致土壤中的養分含量具有一定的差異[30]。本研究中環境因子聚類分析表明,土壤AK 與總豐度前14 的真菌門均無顯著相關性。環境因子聚類分析表明,AN 和SUR 聚為一類,AP 和AK 聚為一類,土壤真菌群落與pH、AN、SUR 和AP 具有顯著(P<0.05)或極顯著(P<0.01)相關性,對真菌菌群影響表現為SUR>AP>AN>pH。
