雜草抗藥性分子機制研究進展
2022-04-11 03:22:12項青
花卉 2022年6期
項青
(蕪湖職業技術學院園林園藝學院,安徽 蕪湖 241003)
0 引言
雜草與人類生活息息相關,其影響最大的方面就是農業生產。雖然農業生產中植物病原物、農業害蟲和農田雜草均會給作物的產量和質量造成巨大的損失,但其中以雜草造成的損失最大,可以造成34%的產量損失[1]。雜草主要通過與農作物競爭水分、肥料、生存空間和的陽光等,影響作物的正常生長發育,從而影響作物的產量和質量。除此之外,一些雜草還會作為病原物的載體,傳播病原。因此,防治雜草是保障糧食產量的重要方式。在化學除草劑出現之前,主要是通過物理防治,在田間人工拔除雜草。20 世紀30 年代,化學除草劑被用于防治雜草,最初除草劑能夠很好地對雜草進行防治,但是隨著大量、長期、頻繁地使用,雜草逐漸對現有的除草劑均產生了抗性[2]。由于雜草和作物之間高度的相似性,因此目前大部分的除草劑都為選擇性的除草劑。雜草對選擇性除草劑的抗性發展較廣譜的除草劑更快。目前,雜草已經對幾乎所有類型的除草劑均產生了抗性。雜草抗藥性的產生導致除草劑的防效降低。為保證糧食產量,就需要使用更加大劑量的除草劑,這又會加劇抗藥性的發展,形成一個惡性循環,最終會導致無藥可用的后果。除此之外,大劑量除草劑的使用還會導致環境的污染,危害人類的生命健康。因此,對抗藥性的研究和治理十分緊迫[3]。
雜草抗藥性的發展是以種群為單位,建立在帶有耐藥基因型的個體在種群內不斷積累發展起來的。在無除草劑使用的種群中包含不同基因型的個體,其中帶有敏感基因型的個體占主體,只有少量帶有耐藥基因的個體。在使用除草劑后,大量敏感個體的生長會受到抑制或死亡,而耐藥的個體受到的影響則較小。長期的除草劑使用,種群中耐藥的個體產生更多的子代,種群中耐藥的個體積累,達到一定的閾值,種群就會表現出對除草劑的抗性(圖1)。因此,對雜草抗藥性進行治理,首先是在分子水平上找到抗藥性形成的機制[4]。
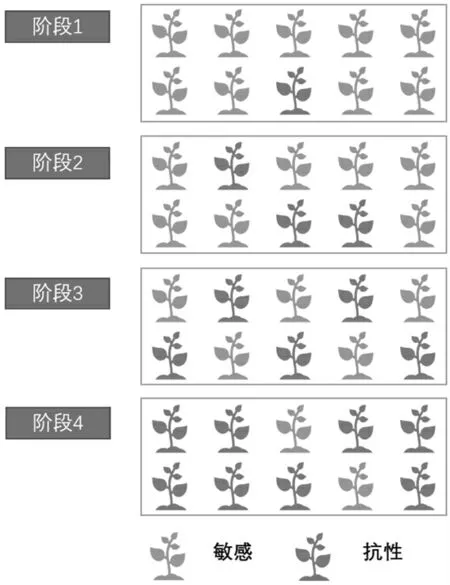
圖1 雜草抗藥性在種群中的發展過程
1 雜草抗藥性的分子機制
除草劑防治雜草的作用機制是除草劑分子噴施到雜草上,進入到雜草細胞中與細胞中的受體結合,導致受體的正常功能紊亂,從而導致的雜草生長發育受到抑制或死亡。在發揮作用的過程中會存在很多的阻礙的方式,這些都會導致除草劑不能發揮作用,從而使雜草表現出耐藥的表型。根據這個過程,目前報道的主要機制包括靶標抗性、非靶標抗性等。
1.1 靶標抗性
選擇性除草劑的開發主要就是依據其可以作用于特定的雜草的靶標,而不傷害作物。因此,除草劑靶標是抗藥性研究中的重中之重。因為除草劑分子與雜草中靶標的接觸是特異性的,因此雜草靶標上發生一些結構性的變化都有可能導致除草劑分子與雜草靶標的親和力受到影響,造成除草劑起不到作用。靶標蛋白上發生的突變包括氨基酸替換、缺失、插入和可變剪接等。其中最常見的為部分位點發生點突變[4]。根據除草劑靶標蛋白的不同,目前主要的除草劑可分類以下幾類:乙酰輔酶A 羧化酶抑制劑類、乙酰乳酸合成酶抑制劑類、光系統Ⅱ抑制劑類、乙酰羥酸合成酶抑制劑類和草甘膦等,分別作用于乙酰輔酶A 羧化酶(acetylˉCoA carboxylase, ACCase)、乙酰乳酸合成酶(acetolactate synthase, ALS)、光系統ⅡD1 蛋白(photosystem II D1 protein, PSII D1)、乙酰羥酸合成酶(acetohydroxyacid synthase, AHAS)和5-烯醇式丙酮酸莽草酸-3-磷酸合酶(5-enolpyruvylshikimate-3-phosphate synthase, EPSPS)。目前在這些靶標蛋白上均發生過與抗藥性有關的多種點突變(表1)。Palmieri 等[5]2021 年研究了ALS 蛋白上發生的A122S、A205V、D376E、W574L 和S653N 突變與長芒莧對ALS 抑制劑類除草劑的抗性有關。除此之外,一些靶標基因的擴增,也可導致抗藥性的產生。據報道,EPSPS 基因在至少8 種雜草中擴增最多達到150 多個拷貝[6]。

表1 不同類型除草劑作用靶標和抗藥性相關靶標突變位點
1.2 非靶標抗性
除了靶標抗性之外,一些雜草對除草劑的抗性還與其他的因素導致,其中主要的機制為代謝抗性[4]。一些代謝酶基因包括細胞色素P450 和谷胱甘肽S-轉移酶等代謝能力的增強,會增強對除草劑分子的代謝能力,從而將有毒的大分子轉化為無毒的代謝產物小分子,減小對植物的傷害。P450 通過NADPH P450 還原酶將氧分子插入除草劑分子降低其活性[7];GST 通過將谷胱甘肽與除草劑分子結合,使其對雜草無毒性[8]。Wang 等[9]2013 年報道早熟禾和看麥娘對精惡唑禾草靈的抗性就與P450 基因過表達和基因擴增有關。Cummins 等[10]2013 年報道北方看麥娘和硬直黑麥草對多種除草劑的抗性就與GST 基因過表達和基因擴增有關。另外,通過阻礙除草劑的轉移和液泡隔離除草劑的方式也能增強雜草對除草劑的抗性。Ge 等[11]2011 年使用31P 核磁共振對抗性加拿大蓬進行研究,發現液泡的隔離作用與雜草的抗藥性產生有關。由于非靶標抗性不具有專一性,因此通常產生非靶標抗性的雜草也會對其他雜草產生交互抗性。
2 雜草抗藥性機制研究技術
技術進步對科學研究具有巨大的推動作用,從最開始的表型生物測定到現在的分子生物學檢測,技術的進步讓抗藥性的研究更加快速、方便。目前主要的研究技術主要有基于PCR 的技術、基于Sanger 測序的技術和基于二代測序的技術等。
2.1 基于PCR 的研究技術
PCR 技術廣泛應用于分子生物學的研究,可以用于突變的分型和基因表達量定量。突變的分型是研究靶標突變的主要手段。設計特異性的引物對植物靶標基因片段進行擴增,含有特定基因型的片段可以被擴增,而不含有該基因型的片段則不會被擴增。PCR 等位基因特異性擴增(PASA)就被廣泛用于靶標抗性基因型的檢測。2011 年,Mátyás 等[12]使用雙向等位基因特異性擴增技術(bi-directional PCR amplification of specific alleles,Bi-PASA)對抗三嗪類除草劑的豚草進行檢測,發現光系統IID1 蛋白上的S264G 與抗藥性產生有關。基因表達量定量是研究抗藥性的重要手段。基于PCR 的實時定量PCR 技術能夠對轉錄后的mRNA 進行定量計算。比較不同種群的雜草抗藥性候選基因的表達量,可以得到差異表達的基因,則與抗藥性表型有關。結合基因功能的研究技術,可以確定差異表達的基因是否與抗藥性有關。Gaines 等[13]2010 年使用的實時定量PCR 技術對抗藥性相關候選基因的表達量進行檢測,發現EPSPS 基因在抗性的長芒莧中高表達,進一步研究其表達量升高是導致抗藥性產生的原因,并且其表達量升高是由于EPSPS 基因的擴增導致的。
2.2 基于DNA 微陣列的技術
DNA 微整列技術又稱為基因芯片技術,在硅片、玻璃等表面排列固定一些列的DNA 片段作為探針與待測樣品的中的DNA進行雜交,雜交完成會發有熒光信號的產生,檢測熒光就可以定性檢測雜交是否進行和定量檢測雜交的數量。靶標抗性突變的檢測可以設計含有特定基因型探針進行檢測,若DNA 片段雜交則說明存在特定的基因型。目前,微陣列主要用于檢測殺蟲劑靶標抗性突變,Chung 等[14]2011 年使用低密度的特異性等位基因探針微陣列分別檢測與煙粉虱的擬除蟲菊酯抗性相關的鈉離子門控通道上的M918V、L925I、T929V 和有機磷農藥抗性有關的乙酰膽堿酯酶上的F392W 突變,結果證明該檢測方法表現出了極好的便捷性和準確性。檢測基因表達量的測定是在芯片表面排列代謝基因片段,根據熒光信號的強弱來判斷抗藥性相關基因是否特異性表達。Pilcher 等[15]2017 年使用微陣列技術在全基因組范圍內對DNA 微整列的技術對小麥抗賽克嗪品種和敏感品種的基因表達量進行檢測,發現169 個基因的表達量上調,127個基因表達量下調。相比于基于PCR 的檢測技術,其可以初步實現了高通量檢測,但是也存在一些缺點的,準確性不如PCR 技術,因此檢測的結果需要進行PCR 驗證。
2.3 基于測序建立的技術
隨著人類基因組計劃提出以來,基因測序的技術經歷了飛速的發展。對基因上發生的突變的檢測,使用測序的手段無疑是最直接的了。早期的Sanger 法測序便能對靶標的序列進行測序,比較不同個體的基因序列就能快速找出發生的突變。Loubet 等[16]對抗甲氧咪草煙和苯磺隆豚草的ALS 基因進行Sanger 測序,發現9 個突變與豚草對ALS 抑制劑類除草劑的抗性有關。不過這樣的測序技術需要設計引物擴增后逐個測序,費時費力。二代測序的出現改變了這一情況,對種群內的抽樣的全部個體進行混樣二代測序就能一次性的測序獲得種群內的靶標上發生的全部突變信息,計算突變的頻率,估算抗藥性的發展情況。2020 年,Délye 等[17]使用擴增子測序技術對抗ALS 抑制劑類除草劑歐洲千里光的ALS 基因進行測序,發現D376E、A205T 和W574L 等突變均與其抗藥性有關。Chen 等[18]2014 年對早熟禾的種子進行轉錄組測序,檢測了ALS 蛋白上發生的突變,并計算突變的頻率,檢測結果可以用于估算雜草種群中抗藥性的發展情況。轉錄組測序技術是目前高通量檢測基因表達量的主要手段,通過對敏感和抗性種群進行混樣轉錄組測序,可以方便地計算代謝酶基因表達量的變化。Wang 等2021 年對抗甲基二磺隆菵草和敏感菵草進行轉錄組測序,研究導致其產生非靶標抗性的機制,發現11 個contigs 在敏感和抗性種群間的表達量變化顯著,其中主要為P450 和GST。
3 雜草抗藥性機制研究展望
除草劑是現今集約農業中極度依賴的工具。雜草抗藥性也是農業生產中巨大的挑戰。經過近幾十年的研究,雜草抗藥性的主要機制已經基本明確。但是對于每種雜草每種除草劑的具體機制都不相同,可能是不同的突變也可能是不同的基因,這些都需要根據具體情況研究治理的手段。目前,雖然進行了大量的研究,但是這些研究很分散。我們要善于在已有研究的基礎上總結,并進行更加深入的研究。比如利用互聯網的便利條件,整理總結已有的研究構建數據庫,給相關工作者提供更加全面的信息資源。同時也要善于引進新的技術,特別的組學技術不斷發展的現在,使用一些新的技術可能完成以前需要費時數年的任務。當然,我們研究雜草抗藥性的機制最終是服務于雜草抗藥性的治理。基于機制的研究,基因編輯技術可以用于編輯特定的基因型,使得雜草不再具有抗性,或者開發新型的除草劑,緩解抗藥性的進化壓力。最后,利用生物防治也是能夠緩解抗藥性發展的重要手段,如利用的動物除草等。
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
海峽科技與產業(2016年3期)2016-05-17 04:32:12
營銷界(2015年23期)2015-02-28 22:06:18
現代農業(2015年5期)2015-02-28 18:40:49
中國質量與標準導報(2013年8期)2013-03-11 19:54:07
