酵母耐高溫分子機制的研究進展*
2022-04-12 11:21:32祁艷華覃啟劍房文霞
廣西科學 2022年1期
關鍵詞:途徑
祁艷華,覃啟劍,汪 斌,金 城,房文霞
(廣西科學院,非糧生物質酶解國家重點實驗室,廣西南寧 530007)
酵母作為真菌研究中的模式生物,在工業生產中應用十分廣泛,如用于生產啤酒、葡萄酒、面包、異源蛋白質、疫苗和高價值代謝物等[1-4]。不同于實驗室的培養條件,工業發酵的酵母需要應對的一個重大挑戰是發酵環境的變化,如溫度、低pH值、高滲和高濃度乙醇等,并在此過程中產生高價值代謝物,因此酵母菌株耐受脅迫的能力對工業生產十分重要[5-7]。在酵母多重脅迫耐受機制的研究中,耐高溫是目前國內外發酵行業研究中十分熱門且重要的課題。酵母的最優生長及發酵溫度通常為28-35℃,適宜在37℃以上生長的酵母為高溫酵母[8]。目前工業上常用的發酵菌株為釀酒酵母,其最適生長溫度一般不超過37℃。有報道稱,某些釀酒酵母菌株可在高達43℃的溫度下產生乙醇,但這種耐高溫的釀酒酵母菌株資源極其有限,因此人們常從自然界中分離出多株耐高溫的非傳統酵母菌株,包括耶氏酵母(Yarrowia)、克魯維酵母(Kluyveromyces)、念珠菌(Candida)和畢赤酵母(Pichia)等,這些酵母菌株在高溫下生長良好且能產生有價值的次級代謝產物[9-13]。一個典型的例子是酵母利用木質纖維素原料發酵生產乙醇,該過程需經過高溫預處理和糖化步驟,即將木質素分解為微生物發酵可利用的五碳糖或六碳糖,隨后酵母在中低溫條件下將糖轉化為生物乙醇[14,15]。同步糖化發酵法(Simultaneous Saccharification and Fermentation,SSF)是乙醇生產的首選方法,其優點是水解和發酵可在同一個發酵罐中進行,操作簡單、成本低、完成時間短,缺點是糖化和發酵對溫度的要求不同[16]。因此,獲得具有耐高溫特性的酵母是生物乙醇產業中提高工業生產能力、降低生產成本的關鍵策略。
酵母的耐高溫特性是一個復雜的性狀,由多基因協同表達、多蛋白共同作用來決定。目前的研究發現,酵母已經進化出復雜而微妙的機制,可通過調節特定的代謝途徑以保護機體免受高溫損傷,或通過激活和調節特定的耐熱相關基因來合成特定的化合物[17,18]。為鑒定新基因并闡明酵母耐高溫的復雜機制,研究者們對酵母耐高溫特性進行了深入的研究。本文主要從代謝途徑、基因表達、酶活特性和蛋白質相互作用等4個方面對酵母耐高溫分子機制進行綜述(圖1),為耐高溫酵母的篩選、改造及應用提供理論基礎,有助于酵母耐熱性的合理設計,提高工業生產的效率并降低生產成本。
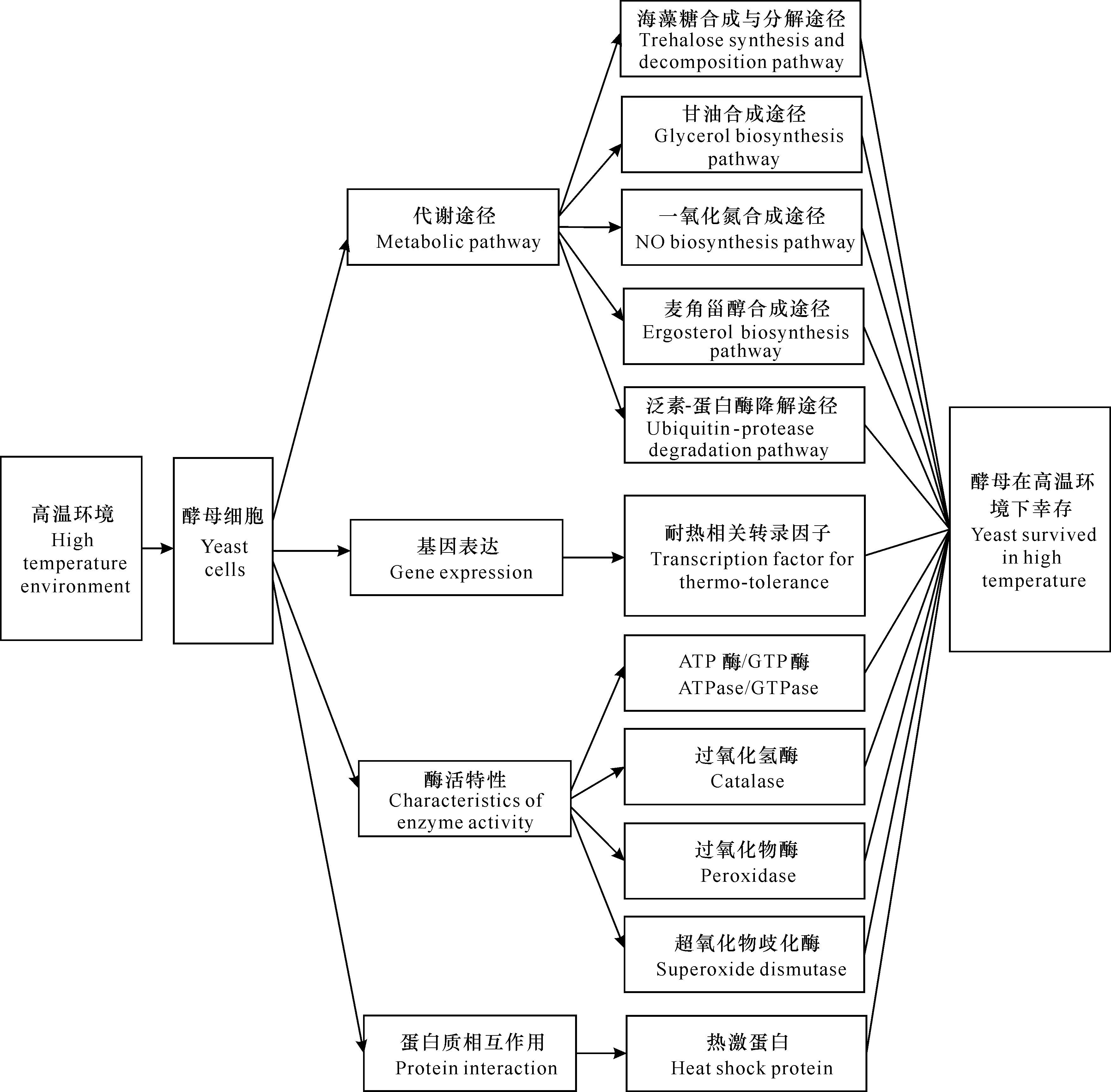
圖1 酵母耐高溫分子機制
1 代謝途徑
新陳代謝為細胞發揮功能提供能量和基礎,新陳代謝的調節需要對不同的外部信號(如環境變化)和內部信號(如細胞周期狀態)作出反應。當酵母處于高溫環境下,其代謝途徑可根據溫度的變化及時調整以適應環境的變化,其中海藻糖合成與分解途徑、甘油合成途徑、麥角甾醇合成途徑、一氧化氮合成途徑、泛素-蛋白酶降解途徑在應對高溫脅迫方面發揮重要的作用[19-22]。
海藻糖是由兩分子葡萄糖組成的非還原性雙糖,存在于多種生物體中,如細菌、真菌、昆蟲和植物等。對稱的葡萄糖分子結構具有較好的穩定性,可作為細胞蛋白質和生物膜的應激保護劑以抵抗環境脅迫[23,24]。在釀酒酵母中,海藻糖由一種酶復合物合成,該酶復合物由tsl1、tps1、tps2和tps3編碼的蛋白組成,其中基因tps1和tps2分別編碼海藻糖-6-磷酸合成酶和海藻糖-6-磷酸酶,而tps3和tsl1編碼復合物的2個調節亞基[25]。海藻糖的分解代謝主要由中性海藻糖酶和酸性海藻糖酶負責[26,27]。nth1和nth2基因編碼的胞漿中性海藻糖酶受cAMP依賴的磷酸化過程、營養物質和溫度的調節,負責海藻糖的細胞內穩態或循環;由ath1基因編碼的空泡酸性海藻糖酶受營養物質的調控,負責利用胞外海藻糖[28,29]。當酵母細胞受到高溫脅迫時,海藻糖合成與分解途徑中參與合成與分解的基因表達水平升高,促進海藻糖的合成與分解,使得細胞既可利用其作為高溫保護劑,也可分解為單糖后進入中心糖代謝為細胞應對高溫環境供能;當細胞遠離脅迫環境時,細胞內海藻糖-6-磷酸含量的增加可抑制Tps1的活性,不利于海藻糖的合成[19,30]。
甘油是酵母在高溫條件下的主要保護劑之一,其主要作用原理是甘油的積累使酵母重新調整細胞膜的滲透壓以保證在高溫脅迫下細胞功能的正常發揮,從而維持酵母正常的生命活動。在酵母發酵的過程中,甘油是僅次于乙醇和二氧化碳的第三大發酵產物,其合成途徑受2個關鍵基因gpd1和gpd2的調控[31,32]。有研究證實,gpd1和gpd2基因是細胞在高溫脅迫下生長所必需的,且受高滲甘油(High Osmolarity Glycerol,HOG)途徑的調節[33,34]。當酵母受到熱刺激時,通過Sho1-Ste20-Ste50-Ste11蛋白級聯將信息傳遞下去可激活HOG途徑中關鍵的蛋白激酶Hog1或通過Nts1將細胞壁完整性(Cell Wall Integrity,CWI)途徑與HOG途徑連接起來。Hog1由Pbs2磷酸化后進入細胞核激活gpd1和gpd2基因的表達,同時甘油分泌途徑中Fsp1通道關閉,導致甘油分泌減少而間接增加細胞內的甘油含量,保護細胞免受高溫的侵害[20,35]。
一氧化氮作為一種信號分子,參與多種細胞功能的調節,其主要來源是精氨酸代謝。當酵母受到高溫沖擊時,精氨酸可轉化為瓜氨酸和一氧化氮,低水平的一氧化氮對酵母具有一定的保護作用[36]。在酵母合成一氧化氮的過程中,除一氧化氮合成酶發揮作用外,酵母的黃素蛋白Tah18也參與其中。基因敲除結果表明,Tah18依賴的一氧化氮合成賦予酵母細胞耐高溫脅迫的能力[22]。
麥角甾醇是真菌細胞膜的重要組成部分,決定膜相關蛋白的流動性、通透性和活性[37]。酵母在高溫應激下生長可使細胞膜的流動性增加,影響蛋白質折疊和穩定性,破壞細胞骨架結構以及細胞代謝平衡[38]。有研究表明,甾醇脫氫酶Erg3和Erg5的缺失與酵母的耐熱性相關,“扁平”甾醇如麥角-8(9),22-二烯醇或麥角-5,7,22,24(28)-三烯醇替代麥角甾醇不會增加釀酒酵母的耐熱性,但“彎曲”甾醇如糞甾醇可增加酵母的耐熱性。因此,erg3基因突變后會導致甾醇甲基轉移酶Erg6活性增強,Erg6催化酵母甾醇甲基化形成糞甾醇,從而引起“彎曲”甾醇的積累,賦予酵母更高的耐熱性[39];erg5基因的缺失,導致細胞積累更多具有飽和側鏈的甾醇促使細胞膜具有更好的排序效果,更好地維持細胞膜的流動性,因此可使erg5突變體比未突變erg5的酵母菌株具有更好的耐熱性,并且該基因的功能在真菌中具有一定的保守性[21]。此外,有研究表明ABC轉運蛋白基因Pdr18的缺失會導致細胞內麥角甾醇合成前體的積累,這種積累可能觸發Erg1表達的反饋抑制,使得酵母細胞表現出更高的滲透性以及對多種環境的挑戰,如高溫環境的敏感性[40]。
高溫環境會引起酵母細胞中蛋白的錯誤折疊從而影響蛋白活性,因此細胞可通過泛素-蛋白酶降解途徑降解錯誤折疊的蛋白以保護細胞免遭功能失調的危機,例如酵母中rsp5基因的過度表達可使細胞具有更高的耐熱性[41,42]。rsp5基因編碼E3泛素連接酶,參與蛋白質的泛素化,調節細胞結構中蛋白質的運輸和最終降解[43,44]。此外,Rsp5可與Bul1、Bul2、泛素結合蛋白Ubc1、泛素蛋白酶Ubp4和Ubp16等蛋白相互作用以提高應激反應基因spi1的表達水平,進而提高細胞對熱休克和氧化應激的耐受能力[45]。
此外,通過轉錄組、蛋白質組等技術,已有對高溫酵母整體代謝變化趨勢的相關研究。例如,通過對定量蛋白質組學和轉錄組學的數據進行綜合分析,Li等[46]發現,酵母高溫發酵后,馬克斯克魯維酵母參與轉錄、翻譯、氧化磷酸化和脂肪酸代謝的蛋白下調,一些分子伴侶和蛋白酶體蛋白上調,ATP酶活性顯著降低,總脂肪酸逐漸積累。Fu等[13]通過轉錄組分析發現,高溫可刺激馬克斯克魯維酵母的線粒體呼吸,抑制三羧酸TCA循環,導致活性氧(Reactive Oxygen Species,ROS)含量增加,并產生更多麥角甾醇來應對乙醇脅迫。Chamnipa等[47]發現,庫德氏畢赤酵母RZ8-1在高溫42℃乙醇發酵過程中,熱休克蛋白基因(ssq1、hsp90)、乙醇脫氫酶基因(adh1、adh2、adh3、adh4)以及甘油醛-3-磷酸脫氫酶基因(tdh2)的表達水平升高,表明有些基因的表達不僅限于乙醇發酵途徑,也與酵母的耐高溫相關。總之,酵母在高溫與適溫下的代謝調節存在顯著差異,細胞可通過控制耐高溫相關代謝基因的表達來調節代謝產物的含量,從而有利于酵母在高溫下生存。酵母細胞與耐熱相關的代謝途徑詳見表1。

表1 酵母細胞與耐熱相關的代謝途徑
2 基因表達
生物體耐高溫是多基因協同表達以賦予細胞在異常溫度下存活和生長的特性。通過分子生物學、分子遺傳學、生物信息學、組學分析等手段,目前已有許多耐高溫相關基因被鑒定,使得對生物體耐高溫的遺傳基礎有了深入的了解。酵母的耐高溫特性是酵母在工業發酵中十分重要的特點之一,當遭遇高溫脅迫時,酵母通過調控自身的基因表達水平以適應高溫環境。
有研究表明,酵母的轉錄因子Hsf1、Msn2、Msn4、Yap1、Hac1、Rlm1、Cad1在高溫環境下可調控多個基因的表達水平,以提高細胞耐高溫的能力[48-54]。熱休克轉錄因子Hsf1非常保守,其核心結構包括翼螺旋-轉螺旋DNA結合域以及對三聚體形成至關重要的疏水螺旋區域,此外還有兩個分別位于N端和C端的反式激活域。正常情況下Hsf1是不活躍的單體或二聚體形式,當酵母受到高溫脅迫時可形成三聚體并高度磷酸化,然后與熱應激相關基因的啟動子nGAAn反向重復序列結合,發揮其轉錄活性,激活相關基因的轉錄[55-57]。Msn2和Msn4已被證實是釀酒酵母中兩個對溫度敏感的轉錄激活子,其氨基酸序列具有一定的同源性且識別DNA序列的氨基酸殘基高度保守[58]。正常條件下Msn2和Msn4蛋白位于細胞質,當細胞處于高溫環境則被運輸到細胞核,隨后蛋白中的鋅指結構可直接結合到DNA序列上或通過其他蛋白間接調控約200個基因的表達[59-61]。氧化休克因子Yap1通過共價作用將傳感器蛋白Gpx3激活,使其向細胞核移動,上調抗氧化基因的表達,如Yap1可誘導gsh1和gsh2的表達,以便酵母細胞在處于熱休克期間合成還原型谷胱甘肽,從而提高細胞的耐受性[62-64]。轉錄因子Hac1作為一種堿性亮氨酸拉鏈蛋白,可與未折疊蛋白反應(Unfolded Protein-Response,UPR)元件結合從而調節參與UPR的基因表達[52]。Rlm1作為蛋白激酶C(Protein Kinase C,PKC)途徑的1個轉錄因子,其轉錄活性被MAP激酶Mpk1通過Ser427和Thr439的磷酸化來調節,進而調控至少25個與細胞壁合成相關基因的表達,用于維持細胞壁的完整性[53]。Cad1作為轉錄激活劑,參與蛋白質穩定性相關基因的表達調控[54]。除上述轉錄因子外,Skn7可與Hsf1相互作用共同激活hsp12、hsp26、hsp70、hsp82和hsp104基因以響應溫度變化帶來的氧化應激[65]。
3 酶活特性
細胞需要不同的氧化環境以促進蛋白質折疊和活性,保證生命活動的正常進行。當細胞遇高溫、高滲透壓等不利的環境條件時,會導致細胞氧化還原功能障礙,影響細胞生長代謝過程,如信號轉導,RNA、DNA和蛋白質的合成以及細胞周期調節等,因此細胞可通過ATP酶催化ATP水解產生能量,維持細胞穩態[66,67]。酵母的質膜H+-ATP酶(Plasma Membrane H+-ATPase,Pma1)由基因pma1編碼,屬于分布廣泛的一類重要的P2型ATP酶[68]。Pma1是產生電化學質子梯度的重要酶,可穿過質膜,為次級溶質運輸系統提供能量,維持離子穩態和細胞內pH值,并間接調節細胞代謝[69]。
生物體賴以生存的環境中營養可用性、滲透壓平衡、溫度和有害物質在不斷變化。為抵御這些外部壓力,所有需氧生物都配備了廣泛的分子伴侶,如氧化還原分子伴侶[70,71]。酵母胞漿過氧化物酶PrxS可根據環境條件從低分子量(Low Molecular Weight,LMW)形式轉變為高分子量(High Molecular Weight,HMW)形式。當其處于LMW形式時,PrxS主要發揮過氧化物酶功能;而處于HMW形式時,PrxS可作為分子伴侶發揮功能,這種過氧化物酶-分子伴侶功能的轉換賦予酵母細胞抗應激能力[72,73]。有研究表明,酵母細胞遭遇熱應激后,可誘導Ytp1 (一種GTP酶)可逆聚成HMW形式,這種結構變化使Ytp1失去原有的GTP酶活性,但賦予該蛋白一種新的分子伴侶活性;當細胞從熱應激中恢復,Ytp1蛋白會回到LMW形式,發揮其GTP酶功能。Ytp1蛋白在HMW和LMW形式之間的變換,賦予酵母細胞適應不同溫度環境的能力[74,75]。此外,Rho1也是一個具有GTP酶活性且與酵母氧化應激相關的蛋白,已有報道證實Rho1的突變體對溫度敏感且會使胞質中ROS積累水平升高,因此Rho1可通過調節包括Ycf1在內的多種下游靶點來保護酵母細胞免受氧化應激帶來的損害[76,77]。
在正常的細胞代謝過程中,活性氧ROS主要通過呼吸和光合電子傳遞鏈不斷產生[78]。這些高反應性分子可與多種細胞成分發生反應,破壞DNA、蛋白質和脂質[79]。因此,細胞必須有一定的防護機制嚴格控制ROS的濃度。當酵母細胞遭受熱應激后,會導致細胞的氧化損傷,因此酵母細胞也含有超氧化物歧化酶SOD、過氧化氫酶和過氧化物酶Prx等抗氧化系統,以去除細胞中的活性氧[19]。酵母細胞中的超氧化物歧化酶,包括線粒體基質中的錳超氧化物歧化酶SOD2和胞漿/膜間隙中的銅鋅超氧化物歧化酶SOD1,可將過氧化氫歧化為水和二氧化碳,保障電子傳遞鏈的正常運行,維持細胞穩態[80]。過氧化物酶體過氧化氫酶A(Cat1)和細胞溶質過氧化氫酶T(Ctt1)在細胞受到不同的應激條件(如熱應激)時可被誘導表達,因此這兩種酶的活性對于保護酵母細胞在高溫條件下免遭氧化損傷的侵害至關重要[81]。過氧化物酶可分為4種類型:2-Cys-Prx,1-Cys-Prx,Prx-Q和Ⅱ型Prx (PrxⅡ),這4種類型的過氧化物酶均具有同樣的反應機理,即催化半胱氨酸接收來自過氧化氫的氧原子,生成磺酸后,再由不同的分子(如硫氧還蛋白TRX)將其再生為還原形式,從而達到去除過氧化氫的目的。釀酒酵母的過氧化物酶主要是2-Cys-Prx (Tsa1、Tsa2、Ahp1和Dot5)和1-Cys-Prx (Prx1)兩種類型,其中Tsa1和Tsa2已被證實與酵母耐熱性相關,而Prx1能夠以硫氧還蛋白特有的方式去除過氧化氫,維持線粒體在高溫條件下的穩定性[79]。酵母細胞中與耐熱相關的酶見表2。

表2 酵母細胞中與耐熱相關的酶
4 蛋白質相互作用
細胞體內含有多種多樣的蛋白質,蛋白質發揮自身活性作用或與其他蛋白質相互作用構成細胞的生化網絡,維持細胞的生命活動。熱休克蛋白可作為分子伴侶在酵母熱應激期間協助蛋白質折疊以及復性錯誤蛋白或激活沉默蛋白,以維持細胞的正常功能。
有研究表明,酵母熱休克轉錄因子Hsf1的活性抑制依賴于熱休克蛋白Hsp70-Ssa1的直接結合,即Hsp70-Ssa1可利用兩個暴露于蛋白表面的半胱氨酸殘基與Hsf1結合來抑制Hsf1的活性,當細胞缺乏Hsp70-Ssa1時,轉錄因子Hsf1不能有效失活[82-84]。除Hsp70外,轉錄因子Hsf1也可與Hsp90相互作用,增加細胞短期的熱適應[85]。Hsp90作為酵母細胞中重要的熱休克蛋白,有Hsp82和Hsc82兩種亞型,除能與轉錄因子Hsf1相互作用外,其活性和靶蛋白特異性受到其他分子伴侶,如Sti1、Cns1、Cdc37、Sba1、Aha1、Cpr6、Cpr7和Ppt1的調控。例如Sti1和Cns37可促進Hsp70/Hsp90的相互作用,Cdc37可調節Hsp90作為折疊蛋白酶的活性,防止某些蛋白的聚集,而Sba1、Aha1、Cpr6、Cpr7和Ppt1可控制Hsp90作為ATP酶活性的分子伴侶來調控細胞的能量供應[86-89]。熱休克蛋白Hsp104在進化上是Hsp100家族中非常保守的一員,作為一種應激誘導型分子伴侶,可形成六聚體與Hsp70、Hsp40協同作用激活一些損傷蛋白,從而保護酵母細胞免受高溫環境脅迫引起的損傷[90]。除上述的熱休克蛋白Hsp70和Hsp90外,小熱休克蛋白sHsp26通常以二十四聚體的形式存在,當細胞遭受高溫時可及時解聚成二聚體與Hsp70、Hsp104相互作用對相關底物進行復性[91,92]。上述研究表明,酵母可利用不同的熱休克蛋白Hsp、小熱休克蛋白sHsp、熱休克轉錄因子等蛋白之間的相互作用,提高細胞對溫度變化的適應能力,從而促進細胞在高溫環境下的生存能力。
5 結束語
酵母作為工業上應用廣泛的微生物之一,其耐高溫的性能對工業發酵具有十分重要的作用。盡管人們對酵母耐高溫的分子機制研究已取得較大的進展,但細胞耐受高溫脅迫是一個復雜的過程,仍存在許多未解之謎亟待研究,如酵母細胞對高溫環境感知與應答的分子機制,是否有其他未報道的轉錄因子參與耐高溫的調控,非傳統酵母與傳統酵母的耐高溫分子機制存在哪些差異,非傳統酵母在耐高溫脅迫方面優于傳統酵母的原因等。因此,對酵母耐高溫分子機制進行深入研究,不僅有利于我國發酵工業對耐高溫菌株的選育,而且有利于能耗的降低和水資源的利用,簡化和優化發酵過程,促進我國發酵工業的環境友好型轉變。
猜你喜歡
語數外學習·高中版中旬(2023年2期)2023-05-10 13:26:53
語數外學習·高中版中旬(2022年5期)2022-07-13 20:47:51
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
