樟樹、陰香種子萌發及幼苗生長研究
2022-04-15 23:35:23董斌黃永芳張暉張祥會李榮喜李孟南洪文泓劉文
江蘇農業科學 2022年6期
董斌 黃永芳 張暉 張祥會 李榮喜 李孟南 洪文泓 劉文
摘要:樟樹(Cinnamomum camphora)和陰香(Cinnamomum burmannii)是樟科優質的木本植物,生產中以播種繁殖為主。為探索出樟樹和陰香高效苗圃育苗技術,對其苗圃催芽、萌發營養物質變化以及一年生幼苗生長規律開展系統研究。結果表明,樟樹種子和陰香種子苗圃播種的最優組合條件為土壤基質、冬播、50 ℃浸種處理。樟樹種子和陰香種子的可溶性糖、粗蛋白質和脂肪含量均較高,在萌發過程中,3種營養成分整體呈下降趨勢。相反地,2種種子初始淀粉含量均較低,但是在萌發過程中持續升高。樟樹一年生冬播苗各項生長指標均優于春播苗,其最佳播種育苗期是冬季。陰香在冬季或者翌年春季播種育苗均可行,一年生冬播苗和春播苗生長指標差異不顯著。
關鍵詞:樟樹;陰香;種子;發芽;苗木
中圖分類號:S718.43 ??文獻標志碼: A
文章編號:1002-1302(2022)06-0131-06
收稿日期:2021-05-21
基金項目:廣東省高校重點科研平臺和項目(編號:2020GCZX009,2019KZDZX2035);廣東省林業科技創新項目(編號:2017KJCX005);廣東農工商職業技術學院熱帶作物應用技術協同創新中心建設項目(編號:XJZX1902);2020年度廣東省教育廳高職教育教學改革研究與實踐項目(編號:JGGZKZ2020063)。
作者簡介:董 斌(1982—),男,廣東廣州人,博士,副教授,研究方向為城市綠化及作物學。E-mail:bbeenn@163.com。
通信作者:黃永芳,教授,碩士生導師,研究方向為經濟林及森林培育。E-mail:hyfang@scau.edu.cn。
樟樹(Cinnamomum camphora)和陰香(Cinnamomum burmannii)是樟科應用廣泛的優質木本植物,因其生態、用材、藥用、油用等諸多應用價值。目前香樟和陰香已成為我國南方重點發展的鄉土闊葉樹種,作為優良的園林綠化樹種或工業用材等材料進行模范化開發的前景非常廣闊,市場對其苗木需求量也非常巨大。針對這2種優良樹種,科技工作者開展了多方面的研究,近期有環境適應與生態效益[1-2],造林密度與用材生產[3-4],精油提取與精深加工[5-6]等研究成果見報,進一步加深了對這2種樹種的認識。
在諸多的城市綠化苗木來源中,苗圃育苗占據著綠化苗木供應量的90%以上,是主要的苗木繁育途徑。樟樹和陰香可采取播種、扦插、組培等多種方式育苗,其中,種子育苗最經濟高效。針對樟樹種苗培育的研究較多,其中,肖祖飛等跟蹤了不同貯藏條件樟樹種子發芽特性及2個月幼苗的生長指標[7],陳一群等和羅敦等跟蹤了不同移植基質對香樟組培苗幼苗生長的影響[8-9],李彥強等跟蹤研究了樟樹扦插苗的生長規律及生物量[10-12] 。但是與樟樹相比,針對陰香苗木培育開展的研究卻較少見報,僅周紀剛等概括性介紹了陰香栽培技術[13],以及莊姍等對陰香扦插進行了試驗研究[14]。總體而言,基于生產條件對2種樹種開展種苗培育研究較少。因此,本研究基于苗圃條件對樟樹和陰香開展種苗培育試驗,了解種子萌發過程中主要營養物質的動態變化,并對幼苗生長開展一年期的長期跟蹤研究,對于探索一套適合生產一線的苗圃育苗方案、促進產業發展具有積極意義。
1 材料與方法
1.1 試驗材料
從廣西壯族自治區調入成熟樟樹種子和陰香種子各1批,調入前經過種子調制和混樣,可以直接開展后續試驗。種子調回后統一放置于華南農業大學經濟林研究中心實驗室備用,4 ℃低溫密封保存。
1.2 試驗方法
1.2.1 苗圃發芽試驗
分別將樟樹種子和陰香種子放置在常溫、50、80 ℃的水中浸泡48 h,保持種子與水的體積比約為1 ∶3,讓其自然冷卻。浸種結束后將種子分別放入沙和黃心土2種基質中進行自然發芽。所有試驗均在同一溫室大棚中進行,保持環境條件和水肥條件一致性,所有組合重復3次。
發芽率=(實際發芽數/供試種子總數)×100% 。
1.2.2 種子萌發過程中營養物質變化研究
將萌發的種子根據胚根長劃分為5個階段:階段A(浸種前未萌發種子);階段B(胚根長0~<1 cm的種子);階段C(胚根長1~<2 cm的種子);階段D(胚根長2~<3 cm的種子);階段E(胚根長3~4 cm的種子)。分別測定樟樹種子和陰香種子不同萌發階段可溶性糖、可溶性淀粉、蛋白質和脂肪的百分含量。
可溶性糖含量測定采用蒽酮比色法;淀粉含量測定采用蒽酮法;粗脂肪含量測定采用索式抽提法;蛋白質含量通過提取蛋白質上清液后,用Sepectrophotometer ND-1000分光光度計測定。每處理每指標3次重復,取平均值。
1.2.3 基質配備及營養袋育苗
1.2.3.1 栽培基質的配備
栽培基質(泥炭土 ∶椰糠 ∶珍珠巖=20 ∶4 ∶1)與黃心土按1 ∶3~4的比例混合,加入少量石灰、過磷酸鈣和有機肥。
1.2.3.2 營養袋育苗
分別于冬季和翌年春季進行種子播種,待種子萌發后即移入7 cm×11 cm的營養袋中進行育苗。
1.2.4 生長量的測定
1.2.4.1 月度生長指標記錄
隨機挑選100株健壯的樟樹幼苗和陰香幼苗并做好標記,于每月26—30日記錄各株苗木的地徑、苗高和葉片數,為期1年。冬播苗測量時間為3月至翌年2月,春播苗測量時間為5月至翌年4月。
1.2.4.2 年生長量測定
隨機挑選一年生樟樹幼苗和陰香幼苗,測定其植株總干質量、根干質量、莖干質量、葉干質量和葉面積5個生長指標,每指標5個重復。
質量測定采用烘箱高溫烘干法。分別稱取全株、根、莖、葉片的鮮質量,然后將其放于烘箱中,調至135 ℃恒溫烘干4 h至恒質量,隨后快速放入干燥皿中,冷卻后稱取質量。
葉片面積用Li-3100型臺式葉面積儀測定。
1.3 數據分析
采用Microsoft Excel 17.0、SAS 9.0、SPSS 23.0等統計軟件進行數據分析。
2 結果與分析
2.1 苗圃發芽試驗
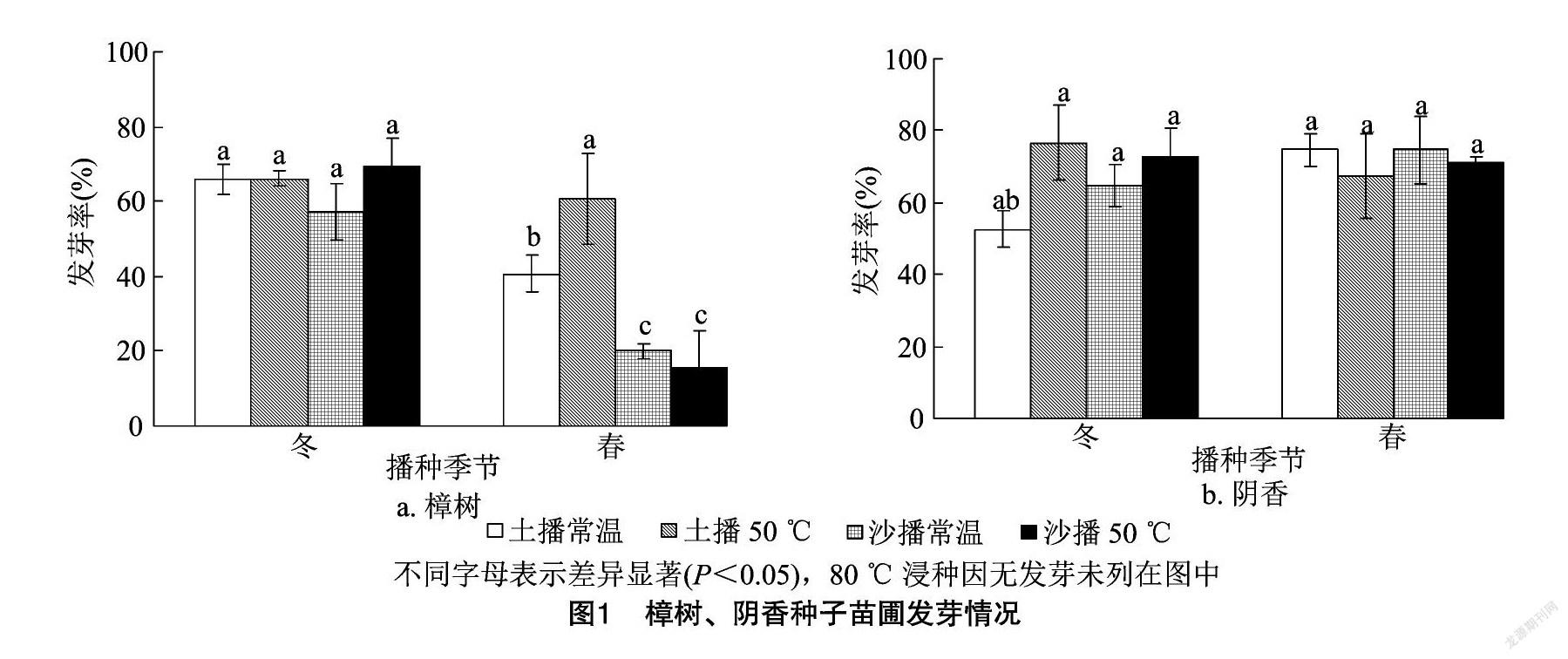
如圖1所示,樟樹種子沙冬播50 ℃條件發芽率最高,達到69.33%;土冬播50 ℃、土冬播常溫和土春播50 ℃這3個處理發芽率較高,達60.00%以上;80 ℃浸種的幾個處理均未見發芽。不同處理的樟樹種子發芽率由高到低排序依次為沙冬播50 ℃>土冬播常溫、土冬播50 ℃>土春播50 ℃>沙冬播常溫>土春播常溫>沙春播常溫>沙春播50 ℃>沙冬播80 ℃、沙春播80 ℃、土冬播80 ℃、土春播80 ℃。
陰香種子土冬播50 ℃處理發芽率最高,達到76.67%;沙冬播50 ℃、沙春播常溫、沙春播50 ℃和土春播常溫這幾個處理發芽率較高,可達70.00%以上;80 ℃浸種的幾個處理均未發芽。不同處理的陰香種子發芽率由高到低的排序依次為土冬播 50 ℃>土春播常溫、沙春播常溫>沙冬播50 ℃>沙春播50 ℃>土春播50 ℃>沙冬播常溫>土冬播常溫>沙冬播80 ℃、沙春播80 ℃、土冬播80 ℃、土春播80 ℃。
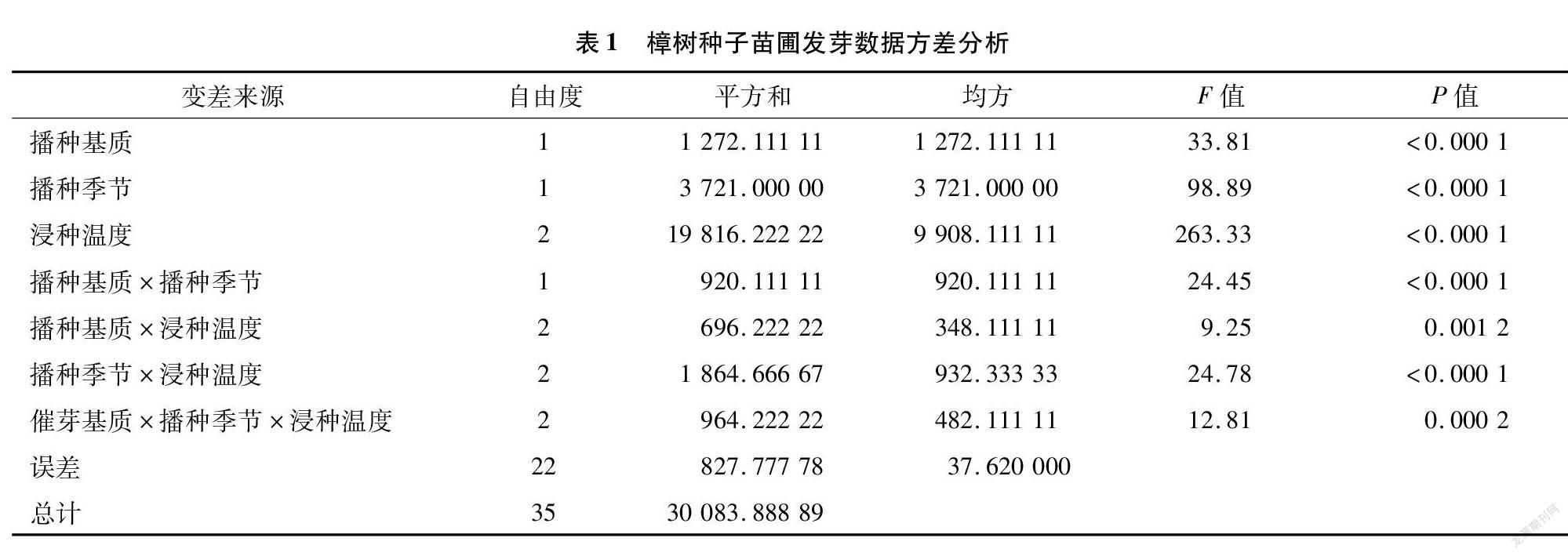
對樟樹種子苗圃發芽結果進行方差分析(表1), 其中催芽基質、 播種季節、浸種溫度、播種基質與播種季節互作、播種季節與浸種溫度互作、催芽基質與播種季節和浸種溫度互作影響極顯著(P<0.01)。
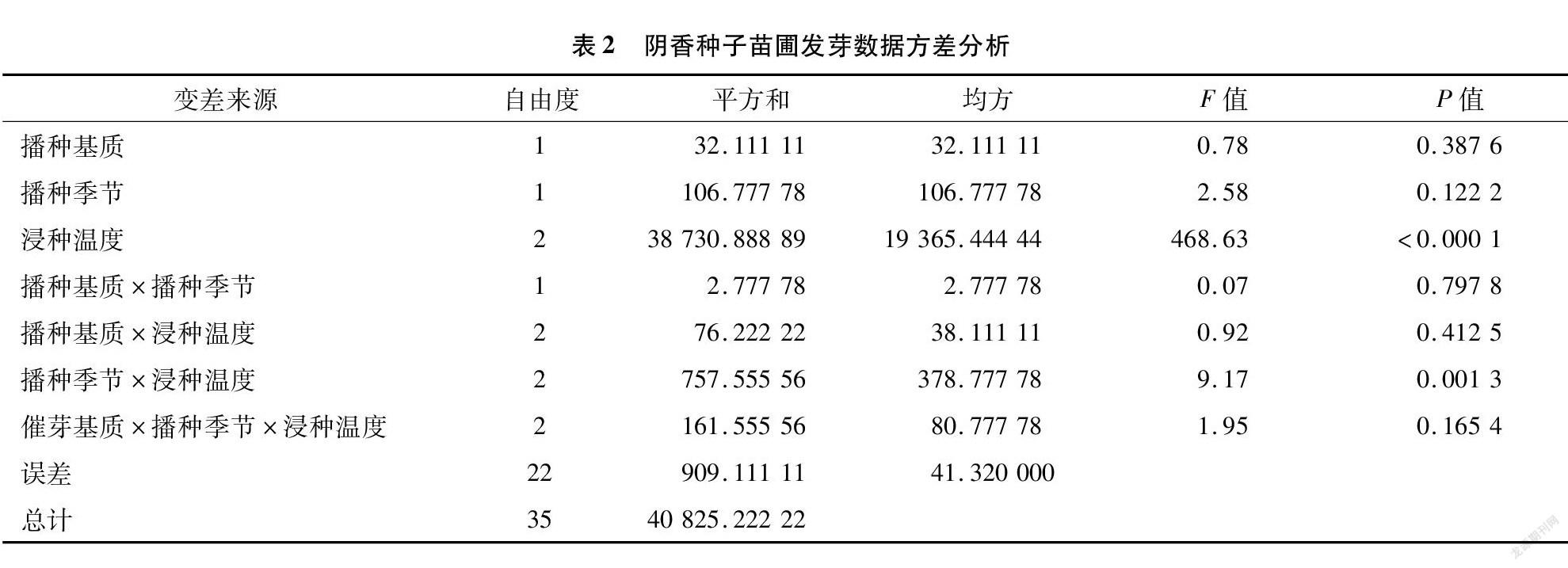
而陰香種子苗圃發芽方差分析結果顯示(表2),浸種溫度、播種季節與浸種溫度互作的影響極顯著(P<0.01),其余結果均不顯著。
2.2 種子萌發過程中營養物質的動態變化
可溶性糖、蛋白質、脂肪和淀粉是種子重要的營養成分。在萌發過程中,樟樹種子和陰香種子可溶性糖、蛋白質和脂肪這3種營養物質含量均整體下降。其中,樟樹種子的可溶性糖、蛋白質含量在萌發初期即大幅下降,之后變化幅度小且波動。至階段E,樟樹種子的可溶性糖、蛋白質和脂肪含量分別降低了50.91%、41.14%和22.19%,其中脂肪的降幅較可溶性糖和蛋白質都小。陰香種子蛋白質和脂肪降幅較大,至種子階段E,2種營養成分分別降低了43.15%和39.58%,而可溶性糖含量相對穩定(圖2)。
2種種子淀粉含量與其余3種營養成分變化相反,呈波動升高。其中,樟樹種子初始淀粉含量為0.17%,至階段E增長至0.69%,為初始值的4.06倍;陰香種子初始淀粉含量為0.47%,至種子階段E含量增長至1.09%,為初始值的2.32倍(圖2)。
2.3 苗期生長特性的研究
2.3.1 一年生苗木地徑變化規律
如圖3所示,冬播樟樹苗3月上袋后地徑即快速增粗,直至10月后逐步增粗放緩;5月上袋的春播苗與冬播苗上袋時地徑值相當,隨后幾個月增粗較快,到10月后進入增粗緩慢增期,變化趨勢與冬播苗一致。綜合可見,3月至10月為樟樹幼苗增粗生長快速期,11月至翌年2月則為緩慢期。
陰香冬播苗6月至7月和10月至翌年3月地徑增粗較慢,其余時間增粗較快;春播苗6月至7月和12月至翌年3月增粗較慢,其余時間段增粗較快。綜合可見,6月至7月和11月至翌年3為陰香幼苗增粗生長緩慢期,其余時間段增粗較快。
2.3.2 一年生苗木苗高變化規律
試驗結果(圖4)顯示,冬播樟樹幼苗3月上袋后出現了2個苗高增長高峰,第1個是6月至10月,第2個是12月至翌年2月。而春播樟樹苗同樣出現6月至9月和12月至翌年3月2個生長高峰,與冬播苗基本一致。綜合可見,樟樹苗9月至12月的苗高增速較慢,其余月份較快。
陰香冬播苗在4月至6月和9月至翌年1月增高緩慢,春播苗在翌年2月至4月增高緩慢,其余時間段增高較快。
2.3.3 一年生苗木葉片數變化規律
從圖5可見,冬播樟樹苗3月至8月葉片數增長較快,8月至12月停滯,隨后又進入新一輪快速增長期。春播樟樹苗5月至9月葉片數快速增長,10月至翌年1月基本停止增加,隨后又進入新一輪快速增長期。
陰香冬播苗9月至翌年1月,春播苗10月至翌年1月,皆為葉片數增長緩慢期,其余時間葉片數增長較快。冬播苗和春播苗葉片數生長趨勢基本一致。
2.3.4 一年生苗木生長量綜合比較
為進一步綜合評價樟樹和陰香冬播苗和春播苗的生長差異,對2種幼苗地徑、苗高、葉數、葉面積指數、植株干質量、葉干質量、莖干質量、根干質量共8個生長指標進行比較,以評價不同播種時期的生長差異。
如表3所示,樟樹一年生冬播苗各項生長量指標均優于春播苗,其中莖干質量、苗高、地徑、葉數這4個指標差異較大。進一步進行方差分析和多重比較,樟樹冬播苗和春播苗的莖干質量、苗高差異極顯著(P<0.01),地徑和葉片數差異顯著(P<0.05)。而陰香一年生冬播苗各生長指標與春播苗相當,均差異不顯著。其中,地徑、苗高、葉面積指數、株總干質量、葉干質量、莖干質量冬播苗稍高于春播苗,而葉片數和根干質量則春播苗稍高于冬播苗。
3 結論與討論
樟樹種子無論采取常溫浸種還是50 ℃溫水浸種,冬播發芽率較高于春播,表明樟樹適宜冬播環境,生產中應優先選擇冬播育苗。而在不同基質的春播催芽試驗中,樟樹土壤基質播種發芽率明顯優于沙基質,原因可能是土壤的保水保溫性能更好,給種子提供更好的水溫環境。如果生產中不可避免需在春季播種育苗,可采用50 ℃土播的催芽方式,以提高樟樹種子發芽率。陰香不同處理條件種子苗圃播種發芽率均較高,冬季采取50 ℃溫水浸種可以明顯提高發芽率。綜合分析浸種溫度對樟樹種子和陰香種子萌發的影響,大部分處理50 ℃溫水優于室溫,主要原因是溫水能促進種子吸水,而種子充分吸水是種子萌發的必要條件,能有效提高種子發芽率[15]。但是,由于80 ℃的浸種溫度過高導致種子死亡,嚴重影響發芽率,因此不能根據樟樹硬質種殼而采用高溫浸種的催芽方式。結合發芽數據和方差分析,樟樹種子苗圃播種最優組合為土壤、冬播、50 ℃浸種處理;陰香種子苗圃播種最優組合為土壤、冬播、50 ℃浸種處理。
對樟樹種子和陰香種子主要營養物質進行測定發現,樟樹種子和陰香種子內脂肪和蛋白質含量均較高,可溶性糖次之,淀粉含量最低。由此判斷樟樹、陰香種子屬于脂肪含量較高的油料種子,而高脂肪含量對于后續種子萌發和幼苗生長具有積極意義[16]。相關研究顯示,脂肪類種子對于貯藏和萌發期環境要求較高[17],必須給予較適宜的貯藏環境和萌發條件。在本試驗中,可溶性糖、蛋白質和脂肪含量隨種子萌發均降低,表明種子通過消耗這些營養物質為萌發提供必需能量[18]。其中,樟樹種子和陰香種子蛋白質和脂肪消耗量較可溶性糖多,表明前兩者是種子萌動的主要營養源。蛋白質會逐漸轉化成可溶性蛋白,進而水解成游離氨基酸[19],用于合成新蛋白質或被去氨基后為呼吸氧化提供碳架,為種子萌發和新組織、新器官的形成提供氮源,對種子各類生理活動的正常進行和萌發過程中的營養供給有重要作用[20]。而脂肪首先水解為甘油和脂肪酸,然后大部分脂肪酸在乙醛酸體內氧化分解為乙酰CoA,并通過乙醛酸循環轉化為糖,進而被萌發時的生長發育等生理活動消耗[21]。樟樹、陰香種子貯藏的脂肪就是通過乙醛酸循環迅速消耗,并且伴有可溶性糖和淀粉增長的現象[22]。可溶性糖能為種子萌發提供碳源、能量等,是萌發到光合自養前的呼吸底物[23-24],而2種種子通過消耗自身的可溶性糖,以及通過脂肪等分解后形成的可溶性糖,均能支持種子的正常萌發。相關研究表明,可溶性糖和淀粉在種子中作為貯藏的碳水化合物,兩者的合成方向和合成量處于動態平衡[25-26],這2個因素可能是導致樟樹種子和陰香種子在萌發過程中淀粉含量不斷升高的主要原因。
對一年生幼苗生長指標的跟蹤發現,樟樹幼苗3月至10月為增粗快速期,6月至9月和12月至翌年3月是增高快速期,4月至9月為葉片數增長快速期;陰香幼苗4月至11月為增粗快速期,6月至10月是增高快速期,1月至10月為葉片數增長快速期。因此,針對性地在莖增粗生長期多施用磷肥和鉀肥,在長葉和拔高期多施用氮肥和鉀肥具有重要的指導意義[27-29]。樟樹一年生冬播苗的各項生長指標均比一年生春播苗高,尤其莖干質量、苗高、葉數和地徑4個指標差異明顯。因此,樟樹最佳播種育苗期在冬季,此時可在同等條件下培育出更健壯的苗木。陰香除根干質量、葉片數這2個指標外,一年生冬播苗的各項生長指標均比一年生春播苗高,但是差異不顯著,因此陰香在冬季或者次年春季進行播種育苗皆宜。
參考文獻:
[1]燕一波. 4種樟科幼樹葉片對汽車尾氣脅迫的生理響應[J]. 四川農業大學學報,2017,35(2):234-240.
[2]李小婷,陳 斌,王海軍,等. 城市夜間燈光對香樟生長的影響[J]. 應用生態學報,2019,30(7):2284-2290.
[3]劉新亮,章 挺,邱鳳英,等. 造林密度對材用樟樹幼林生長和蓄積量的影響[J]. 中南林業科技大學學報,2019,39(3):23-27,60.
[4]陳柏旭,黃騰華,韋鵬練,等. 陰香木材物理力學性質研究[J]. 西北林學院學報,2016,31(5):271-275.
[5]Yang Z R,An W L,Liu S S,et al. Mining of candidate genes involved in the biosynthesis of dextrorotatory borneol in Cinnamomum burmannii by transcriptomic analysis on three chemotypes[J]. Peer J,2020,8:e9311.
[6]Fu J,Zeng C,Zeng Z L,et al. Cinnamomum camphora seed kernel oil ameliorates oxidative stress and inflammation in diet-induced obese rats[J]. Journal of Food Science,2016,81(5):H1295-H1300.
[7]肖祖飛,張北紅,李鳳,等. 樟樹種子發芽及幼苗生長研究[J]. 安徽農學通報,2020,26(7):40-45,63.
[8]陳一群,丘佐旺,汪迎利,等. 樟樹組培苗輕基質育苗技術研究[J]. 亞熱帶植物科學,2015,44(2):140-145.
[9]羅 敦,黃漢林,吳坤林,等. 移栽基質對香樟組培苗生長的影響[J]. 安徽農業科學,2020,48(2):130-132.
[10]李彥強,孫小艷,鐘永達,等. 幾種常見綠化樹種扦插苗生長及固碳能力[J]. 江蘇農業科學,2016,44(7):256-258.
[11]李彥強,胡曉健,高 柱,等. 大棚內不同基質扦插對香樟苗生長的影響[J]. 北方園藝,2015(22):58-60.
[12]葉潤燕,童再康,張俊紅,等. 樟樹莖段組培快繁[J]. 浙江農林大學學報,2016,33(1):177-182.
[13]周紀剛,徐 平,舒夏竺,等. 陰香高效栽培技術[J]. 林業實用技術,2014(4):58-59.
[14]莊 姍,胡松竹,王光云,等. 陰香扦插繁殖試驗[J]. 安徽農業科學,2009,37(18):8639-8640.
[15]程 瑤,方向文,蔣志榮,等. 溫水浸種對蒙古黃芪種子萌發特性的影響[J]. 植物科學學報,2017,35(3):413-420.
[16]Martínez-Arias R,Ronquillo-López M,Schechert A.Quantification of oil content in intact sugar beet seed by near-infrared spectroscopy[J]. Agronomy,2018,8(11):254.
[17]朱 鶴,李憬霖,徐敏,等. 作物種子的營養成分與其萌發特性關系的探討[J]. 種子,2019,38(8):47-52.
[18]Fatihi A,Boulard C,Bouyer D,et al. Deciphering and modifying LAFL transcriptional regulatory network in seed for improving yield and quality of storage compounds[J]. Plant Science,2016,250:198-204.
[19]Liu R R,Wang L,Tanveer M,et al. Seed heteromorphism:an important adaptation of halophytes for habitat heterogeneity[J]. Frontiers in Plant Science,2018,9:1515.
[20]Müntz K,Belozersky M A,Dunaevsky Y E,et al. Stored proteinases
and the initiation of storage protein mobilization in seeds during germination and seedling growth[J]. Journal of Experimental Botany,2001,52(362):1741-1752.
[21]Wei S,Liu Y N,Wu M L,et al. Disruption of the transcription factors Thi2p and Nrm1p alleviates the post-glucose effect on xylose utilization in Saccharomyces cerevisiae[J]. Biotechnology for Biofuels,2018,11:112.
[22]孫 建,周紅英,樂美旺,等. 芝麻種子萌發動態及其代謝生理變化研究[J]. 中國農業科技導報,2020,22(8):41-48.
[23]馬 琳,王志清,張秀蓮,等. 北細辛種子儲藏過程中儲藏物質變化的研究[J]. 種子,2015,34(12):1-3,8.
[24]王海杰,楊 宇,黃家權. 瓜兒豆種子萌發糖代謝動態分析[J]. 山東農業大學學報(自然科學版),2019,50(1):99-102.
[25]Pairochteerakul P,Jothityangkoon D,Ketthaisong D,et al. Seed germination in relation to total sugar and starch in endosperm mutant of sweet corn genotypes[J]. Agronomy,2018,8(12):299.
[26]Tan-Wilson A L,Wilson K A.Mobilization of seed protein reserves[J]. Physiologia Plantarum,2012,145(1):140-153.
[27]高小麗. 施磷對豌豆產量及其構成因素的影響[J]. 作物雜志,2016(1):125-128.
[28]李 琦,裴懷弟,馬忠明,等. 鉀肥與有機肥配施對食用百合根際土壤酶活性、養分含量及鱗莖產量的影響[J]. 中國土壤與肥料,2020(1):91-99.
[29]權寶全,呂瑞洲,王貴江. 不同施氮量對甘薯生長發育及產量的影響[J]. 東北農業科學,2019,44(6):14-17.
