‘陽光玫瑰’雄性不育株花蕾發育期抗氧化酶活性和內源激素含量的變化
2022-04-18 01:40:36陶建敏農慧蘭徐穎一耿藝寧凌千惠
西北植物學報 2022年3期
洪 奔,陶建敏,農慧蘭,徐穎一,耿藝寧,凌千惠,鄭 煥
(南京農業大學 園藝學院, 南京 210095)
葡萄是重要的果樹經濟作物, 在世界農業中占有重要地位,雄性不育在提高水果無核品質、增加產量、商業化批量雜交育種等方面尤為重要[1]。葡萄雄性不育系因其單性結實的特性可培育無核品種,同時在雜交育種中作為母本免去人工去雄這一過程,可極大提高育種效率[2]。因此,研究和應用雄性不育對葡萄雄性不育新種質的合理開發與利用具有重要意義。
植物雄性不育的發生過程中通常伴隨著異常的生理代謝特征。馮小磊[3]認為植物雄性不育的生理生化特性研究可將植物遺傳與個體表型變異聯系起來, 有助于揭示植物雄性不育發生機理。近年來, 不少學者在膜脂過氧化和內源激素方面對植物雄性不育材料的生理生化機制進行了研究, 取得了一系列新進展。活性氧在植物中的積累被認為是雄性不育產生的重要原因之一。它可以使細胞膜中不飽和脂肪酸發生過氧化 或脫脂化,逐級降解為MDA,一般認為MDA在植物體內積累是活性氧毒害的表現,其含量是判斷膜脂過氧化程度的一個重要指標。植物在長期演化過程中形成了防御活性氧的保護機制,其中抗氧化酶系統發揮至關重要的作用[4]。高羊茅[5]、百合[6]、厚軸茶[7]等植物不育株花蕾發育過程中POD、SOD、CAT等抗氧化酶活性較可育株表現出不同程度的差異,而花蕾發育階段MDA含量通常高于可育株。前人普遍認為在雄性不育發生過程中會產生大量的活性氧,導致MDA升高,可能是植物敗育的一個重要原因[8]。另外,植物內源激素參與細胞、組織、器官分化、開花與結實、成熟與衰老、休眠與萌發等諸多方面生理活動,單獨或協同調控植物的生長發育過程。目前,大量研究表明植物花蕾不同發育時期內源激素含量發生變化,都將影響植物自身花藥育性,造成雄性不育[9]。茄子[10]、大豆[11]等作物雄性不育研究結果表明,不育系花器官發育過程中IAA、ABA、GA3、CTK、JA、BR等內源激素含量異常以及內源激素平衡失調與雄性不育的發生緊密相關。
‘陽光玫瑰’葡萄為歐美雜交種。該品種生長勢強、抗病性強、豐產質優、口感香甜、市場認可度高。‘Y-14’為南京農業大學選育的4株‘陽光玫瑰’實生后代雄性不育優良單株,于2017年定植,有性繁殖;其雄蕊花絲極短,且緊緊包裹雌蕊不易開裂(圖1),花粉無活力;自然授粉果穗大小粒現象嚴重,無核率達96%,花前套袋為全無核果實。目前,關于葡萄雄性不育材料生理生化方面的研究較少。本研究以‘陽光玫瑰’實生后代雄性不育株‘Y-14’和‘陽光玫瑰’各發育期花蕾為試材,測定花粉母細胞時期、減數分裂時期、四分體時期、單核早期、單核晚期、二核期、成熟花粉粒時期的抗氧化酶活性和內源激素含量,從而揭示葡萄不育株‘Y-14’膜脂過氧化、內源激素含量與雄性不育之間的關系,闡明葡萄雄性不育的生理代謝特征,為葡萄雄性不育新種質的合理開發與利用提供理論依據。
1 材料和方法
1.1 試驗地點與材料
以南京農業大學湯山翠谷葡萄基地的‘陽光玫瑰’實生后代雄性不育株(編號‘Y-14’)和‘陽光玫瑰’為實驗材料(圖1)。從2021年4月下旬幼蕾期開始采集花蕾至5月中上旬開花為止,每3 d采集1次,液氮速凍,-80 ℃冰箱保存;將花蕾分為花粉母細胞時期、減數分裂期、四分體期、單核早期、單核晚期、二核期及成熟花粉粒時期。
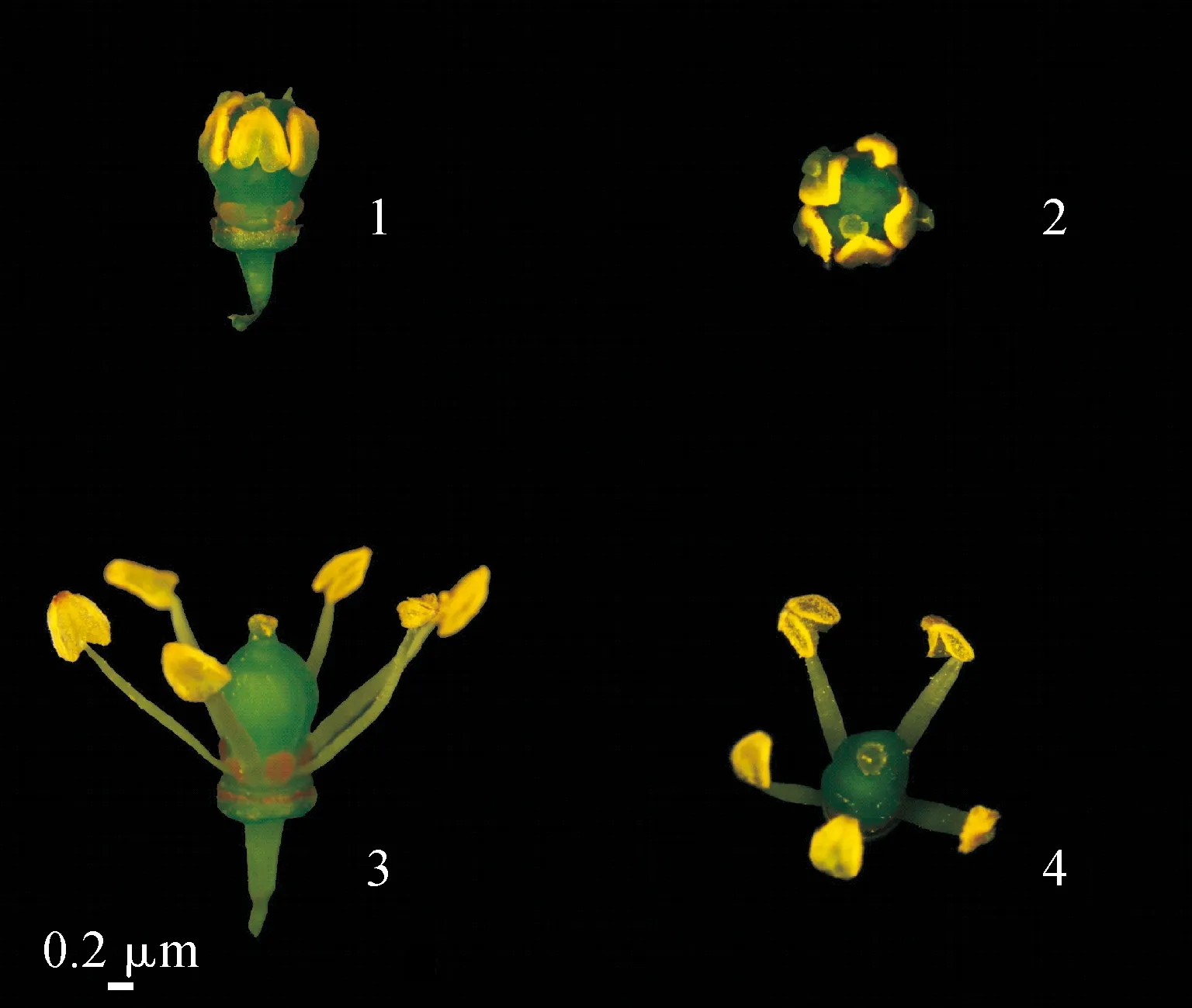
1.‘Y-14’正視圖;2.‘Y-14’俯視圖;3.‘陽光玫瑰’正視圖;4.‘陽光玫瑰’俯視圖
1.2 測定項目
花蕾樣品POD、CAT、SOD活性和MDA含量采用試劑盒測定。試劑盒均購自北京索萊寶科技有限公司,樣品的處理及各項指標測定均按照試劑盒使用說明書進行,每個樣品重復測定3次。
花蕾樣品內源激素含量的定性定量分析采用ESI-HPLC-MS/MS法。以異丙醇-水-鹽酸提取植物樣品內源性激素(endogenous phytohormones),以安捷倫(Agilent)1290 高效液相色譜儀串聯AB Sciex QTRAP 6500+質譜儀測定植物內源性激素IAA、ABA、反式玉米素核苷(TZR)、茉莉酸甲酯(MeJA)、GA3含量,并在提取過程中加入內標物質校正檢測結果。(1)標準溶液配制:以甲醇(0.1%甲酸)為溶劑配制濃度梯度為0.1、0.2、0.5、2、5、20、50和200 ng/mL的IAA、ABA、TZR、MeJA、GA3標準溶液,并加入終濃度為20 ng/mL的內標溶液,在實際繪制標準曲線方程時剔除線性異常點。(2)液相條件:色譜柱:Poroshell 120 SB-C18反相色譜柱(2.1×150,2.7 μm);柱溫:30 ℃;流動相:A∶B=(甲醇/0.1%甲酸)∶(水/0.1%甲酸);洗脫梯度:0~1 min,A=20%;1~9 min,A遞增至 80%;9~10 min,A=80%;10~10.1 min,A 遞減至 20%;10.1~15 min, A=20%;進樣體積:2 μL。(3)質譜條件:電噴霧電離源 ESI(electron spray ionization)霧化溫度:400 ℃;氣簾氣(curtain gas,CUR):15 psi;噴霧電壓(ion spray voltage,IS):4 500 V;霧化氣壓力(Gas1):65 psi;輔助氣壓力(Gas2):70 psi;監測模式:MRM(多反應監測模式 multiple reaction monitoring)。在Q-Trap6500 中,每個離子對是根據優化的去簇電壓(declustering potential,DP)和碰撞能(collision energy,CE)進行掃描檢測。
1.3 數據處理
GraphPad Prism 8.0.1使用Analyst及GraphPad Prism 8.0.1進行處理數據及數據分析,Excel 2019作圖。
2 結果與分析
2.1 ‘Y-14’和‘陽光玫瑰’花蕾各發育時期抗氧化酶活性及MDA含量比較
2.1.1 POD活性‘Y-14’和‘陽光玫瑰’花蕾POD活性隨著花蕾發育時期總體呈逐步下降趨勢(圖2,A)。其中,各發育時期‘Y-14’花蕾POD活性都顯著低于‘陽光玫瑰’POD活性;‘Y-14’與‘陽光玫瑰’的POD活性差異較大時期集中出現在花蕾發育的四分體時期(S3)和單核早期(S4),此時‘Y-14’POD活性分別為同期‘陽光玫瑰’的75.71%和57.31%。
2.1.2 SOD活性圖2,B顯示,‘Y-14’和‘陽光玫瑰’花蕾SOD活性在花蕾發育時期總體上呈先下降后上升趨勢。其中,‘Y-14’和‘陽光玫瑰’花蕾SOD活性均單核早期(S4)最低;整個花蕾發育過程中,‘Y-14’SOD活性始終顯著低于同期‘陽光玫瑰’,并分別以花粉母細胞時期(S1)和單核早期(S4)差異最大和最小,分別占同期‘陽光玫瑰’的37%和59.21%。
2.1.3 CAT活性如圖2,C所示,‘Y-14’和‘陽光玫瑰’花蕾CAT活性在花蕾發育時期總體上呈緩慢下降趨勢,‘Y-14’在四分體時期(S3)有驟降現象。其中,‘Y-14’花蕾CAT活性除減數分裂期略高于‘陽光玫瑰’外,其余時期均低于‘陽光玫瑰’,且除花粉母細胞期(S1)和減數分裂期(S2)外,兩者差異均達到顯著水平;‘Y-14’花蕾的CAT活性與‘陽光玫瑰’差異在花蕾發育四分體期、單核早期、單核晚期更為明顯,分別占同期‘陽光玫瑰’的70.41%、71.92%和63.36%。
2.1.4 MDA含量‘Y-14’和‘陽光玫瑰’花蕾的MDA含量在花蕾發育過程中呈逐步下降的趨勢,且總體而言,‘Y-14’花蕾的MDA含量要始終明顯高于同期‘陽光玫瑰’(圖2,D)。其中,‘Y-14’和‘陽光玫瑰’花蕾的MDA含量均在單核晚期至二核期下降幅度較大;在花粉母細胞時期(S1)和單核晚期(S5),‘Y-14’與‘陽光玫瑰’之間的MDA含量差異尤為明顯,‘Y-14’MDA含量分別為同期‘陽光玫瑰’的1.38倍和1.34倍。
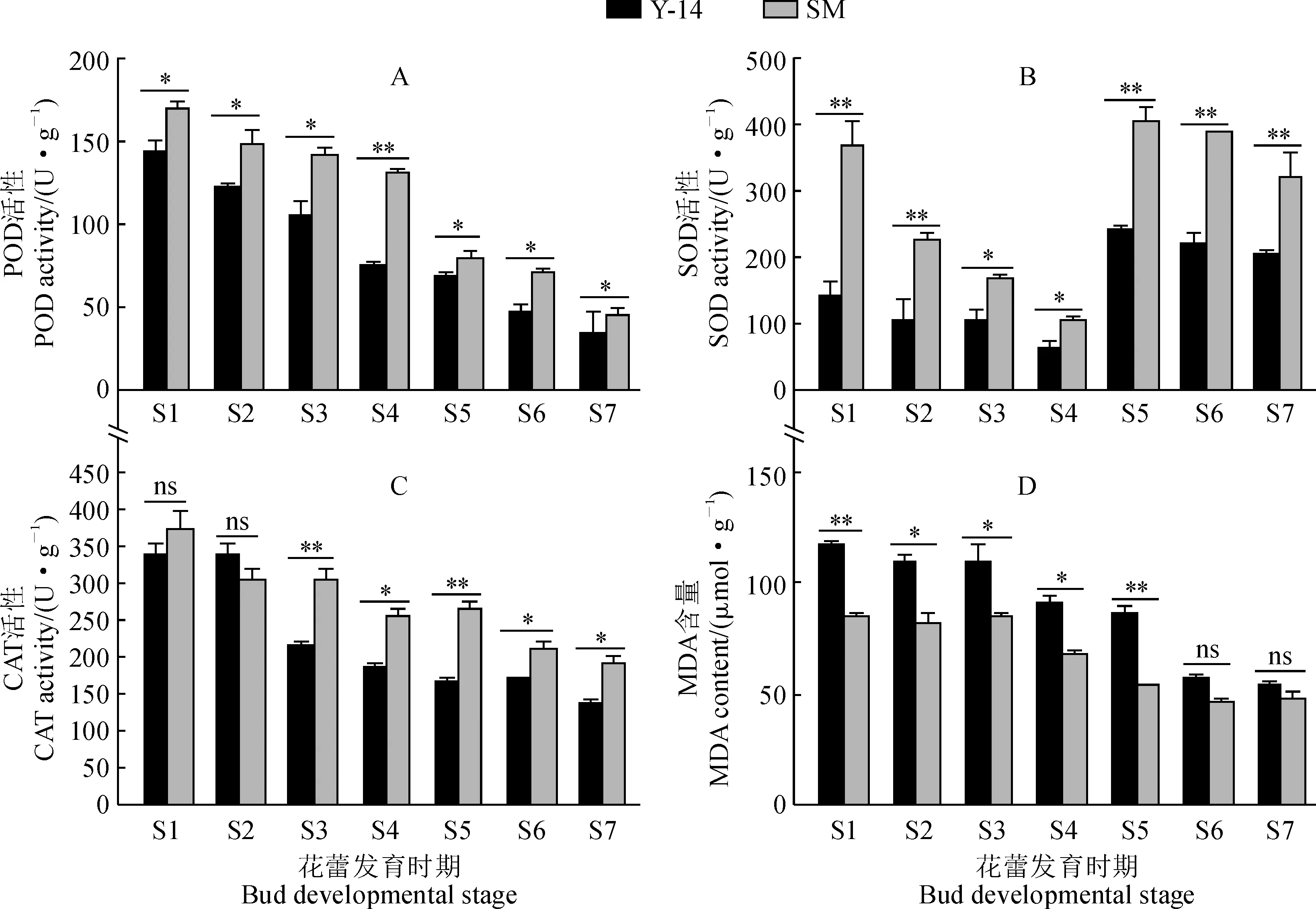
*表示差異顯著(P<0.05),**表示差異極顯著(P<0.01); S1.花粉母細胞期;S2.減數分裂期;S3.四分體期;S4.單核早期;S5.單核晚期;S6.二核期;S7.成熟花粉粒期;下同
以上結果說明‘Y-14’各發育時期抗氧化酶活性總體上低于‘陽光玫瑰’,而MDA含量高于‘陽光玫瑰’,說明‘Y-14’較‘陽光玫瑰’膜脂過氧化程度更嚴重。兩材料花蕾POD與CAT活性總體上呈下降趨勢,POD活性從花蕾發育早期即差異顯著,而CAT活性在四分體期差異極顯著;兩材料花蕾SOD活性均在單核早期達到最低,然后在單核早期突然上升,且各時期均差異顯著。
2.2 ‘Y-14’和‘陽光玫瑰’花蕾發育期內源激素含量變化特征
2.2.1 IAA含量如圖3,A所示,在葡萄花蕾發育過程中,‘Y-14’花蕾IAA含量始終不同程度地高于同期‘陽光玫瑰’,但僅在花粉母細胞時期、四分體時期和單核早期差異達到顯著水平;‘Y-14’花蕾IAA含量總體呈下降趨勢,‘陽光玫瑰’花蕾IAA含量變化趨勢不明顯。其中,‘Y-14’花蕾IAA含量在四分體時期達到最大值,而‘陽光玫瑰’IAA含量在單核早期達到最大值;‘Y-14’的IAA含量在花粉母細胞時期、四分體時期與‘陽光玫瑰’表現出顯著差異,分別是同期‘陽光玫瑰’的1.80倍、1.46倍。
2.2.2 GA3含量‘Y-14’和‘陽光玫瑰’花蕾GA3含量在花蕾發育過程中均呈先上升后下降的變化趨勢(圖3,B)。其中,‘Y-14’花蕾GA3含量在花蕾發育前期(S1~S3)低于‘陽光玫瑰’,在單核早期(S4)開始高于‘陽光玫瑰’含量,但在S1~S4期間兩者間差異均未達到顯著水平;Y-14’花蕾GA3含量在單核期(S5)和二核期(S6)顯著高于同期‘陽光玫瑰’(P<0.05),并以二核期差異最大,此時‘Y-14’GA3含量是‘陽光玫瑰’的1.84倍;‘Y-14’GA3含量在花粉成熟期(S7)又明顯降低,稍低于同期‘陽光玫瑰’。
2.2.3 ABA含量‘Y-14’和‘陽光玫瑰’花蕾ABA含量在花蕾發育過程中總體呈上升趨勢,‘陽光玫瑰’ABA含量在單核早期(S4)顯著大幅上升,但‘Y-14’ABA含量的變化相對比較平緩(圖3,C);在花蕾整個生長發育過程中,‘Y-14’花蕾ABA含量始終顯著低于同期‘陽光玫瑰’,且在花粉母細胞時期就達到極顯著水平,此時僅為‘陽光玫瑰’的23.42%。
2.2.4 TZR含量圖3,D顯示,‘Y-14’和‘陽光玫瑰’花蕾TZR含量在花蕾發育期總體上變化平緩且普遍含量較低(0.05 ng/g左右),‘Y-14’TZR含量始終低于同期‘陽光玫瑰’,但它們之間在各發育時期始終差異不顯著。
2.2.5 MeJA含量在花蕾發育過程中,‘Y-14’花蕾MeJA含量總體呈升高-降低交替變化的趨勢,并在減數分裂期達到最高,而‘陽光玫瑰’花蕾MeJA含量總體呈上升趨勢,并在花粉母細胞時期最低,在成熟花粉粒時期達到最高(圖3,E)。‘Y-14’花蕾MeJA含量在花蕾發育前期明顯高于同期‘陽光玫瑰’,并在花粉母細胞時期差異極顯著水平,是同期‘陽光玫瑰’的2.56倍;從單核早期(S4)開始,‘Y-14’花蕾MeJA含量均不同程度地低于同期‘陽光玫瑰’,并且其間差異隨著發育時期越來越明顯,在二核期和花粉成熟期均達到極顯著水平。

圖3 ‘Y-14’與‘陽光玫瑰’花蕾發育期內源激素含量的變化
以上結果說明整個花蕾發育期以ABA的含量在‘Y-14’與‘陽光玫瑰’間差異尤其顯著,ABA在葡萄花蕾中具有一定的組織特異性。‘Y-14’花蕾IAA含量在花蕾發育過程中高于‘陽光玫瑰’,與ABA呈一定程度負相關。‘Y-14’的MeJA與ABA含量在花粉發育后期均極顯著低于‘陽光玫瑰’,而GA3含量在花粉發育中后期略高于‘陽光玫瑰’,與ABA存在一定程度負相關。‘Y-14’和‘陽光玫瑰’的TZR含量普遍偏低(0.05 ng/g左右) 且無顯著差異,就本研究結果來看,TZR含量變化與‘Y-14’雄性不育的形成無直接關系。
3 討 論
3.1 ‘陽光玫瑰’葡萄雄性不育與膜脂過氧化的關系

在本研究中,不育系‘Y-14’花蕾MDA含量在整個花蕾發育過程中明顯高于‘陽光玫瑰’,而其SOD、POD、CAT活性總體低于‘陽光玫瑰’。此外,‘Y-14’和‘陽光玫瑰’花蕾的MDA含量都在花蕾發育前期最高而后總體呈下降趨勢,這可能與其SOD活性在花蕾發育中期突然升高有關。SOD作為植物細胞中最主要的抗氧化酶,其作為抗氧化酶防御系統的第一道防線,加之較其他抗氧化酶更為穩定的特點[18],其活性變化往往直接影響MDA的含量。由此可見,‘Y-14’和‘陽光玫瑰’葡萄本身抗氧化酶防御系統亦存在一定的應激反應。綜上,本研究中‘Y-14’花蕾的抗氧化酶系統活性較‘陽光玫瑰’降低,而其MDA含量升高,可能是造成‘Y-14’敗育的原因;同時,‘Y-14’與‘陽光玫瑰’抗氧化酶系統中SOD活性差異最為顯著,推測SOD活性是影響‘陽光玫瑰’實生后代雄性不育的關鍵。這與其他一些作物雄性不育研究類似,但也存在與之相反的結論。如劉倩在‘魏可’葡萄研究中發現不育株SOD活性高于可育株[19]。因此,即使在相同物種的不同品種中,相關研究結論也不盡相同,這可能與植物種類、不育發生關鍵時期,甚至栽培條件與生長條件均有關。
3.2 ‘陽光玫瑰’葡萄雄性不育與內源激素的關系
植物通過內源激素調控能量代謝、物質代謝、信號傳導以及基因表達等諸多方面,內源激素平衡維持植物正常的生理活動,保證植物正常的生長發育[20]。迄今為止,大量研究表明植物內源激素如IAA、GA3、ABA、CK、JA、BR等與雄性不育的發生密切相關,相關內源激素含量過高或是過低都會引起植物雄性不育[13]。本研究所測的5種內源激素中,整個花蕾發育期以ABA的含量在‘Y-14’與‘陽光玫瑰’間差異尤其顯著。研究表明:ABA可誘導植物體內相關酶的重新合成來提高植物的抗逆性[31],在小麥的研究中發現ABA可以顯著提升小麥抗氧化酶(POD、SOD和CAT)活性[21]。故推測‘Y-14’花蕾ABA含量較低導致了抗氧化酶活性的降低,加劇了‘Y-14’的膜脂過氧化,最終導致雄性不育。這與‘魏可’葡萄不育株中的研究結果一致[22]。然而,也有大量研究表明ABA含量過高會影響維管系統的發育,促使營養物質轉運受抑制,小孢子母細胞因營養匱乏發育受阻,從而造成雄性不育[7]。劉紅艷認為ABA盈積可能會伴隨著花粉母細胞的衰老和死亡,導致花粉敗育和雄性不育的發生[23]。這一觀點與西瓜[24]小麥[25]的研究結果類似。馬艷青認為同一內源激素在不同的作物種類中與雄性不育的關系表現并不一致,不同的植物或不育源可能存在不同的調控基因[26],因此推測葡萄雄性不育發生與ABA含量關系具有物種特異性。
前人普遍認為IAA的虧損會導致植物維管束的異常發育,影響營養物質在花藥的正常分配,從而導致雄性不育[27]。研究發現小麥[27]、西瓜[24]等植物中不育系IAA含量均低于可育系。本研究結果卻與之相反,‘Y-14’花蕾IAA含量在花蕾發育過程中顯著高于‘陽光玫瑰’,這可能與ABA能抑制IAA的合成有關,‘陽光玫瑰’花蕾ABA含量較高導致了IAA含量的降低。也有研究表明POD活性升高可能會導致IAA的分解[7],‘陽光玫瑰’POD活性較高也可能導致了IAA含量的降低。另外,在百合[6]、水稻[3]、油菜[28]多項研究中發現不育株IAA含量高于可育株。這可能是因為高濃度的IAA可誘導乙烯過量合成,進而促進雄性表達或誘導雄性不育[29]。
GA3是一種活性最高的赤霉素類激素。研究表明,GA3可促進雄花分化,在大白菜[30]、葡萄[22]、西瓜[24]等多種作物中,不育株GA3含量在花蕾各發育時期均低于可育株。在本研究中,‘Y-14’花蕾發育前中期GA3含量低于‘陽光玫瑰’,與前人結果類似;但花蕾發育后期‘Y-14’含量高于‘陽光玫瑰’,這種GA3含量變化情況與厚軸茶中研究結果類似[7],故推測‘Y-14’花蕾GA3含量在花蕾發育前期和發育后期異常變化可能導致花粉母細胞發育異常,從而影響了‘陽光玫瑰’實生后代雄性不育。
MeJA最先在擬南芥中被證明與花藥開裂有關[31],隨后在水稻、小麥等大量研究中發現與雄性不育密切相關[32-33]。擬南芥雄性不育突變體中花藥不能正常開裂,同時出現JA含量低、花絲伸長受到抑制的現象[34]。本研究中‘Y-14’在花蕾發育后期MeJA含量顯著降低的現象與上述研究結果類似。郭航認為JA是誘導花絲伸長、花藥開裂的信號分子,在花藥開裂過程中起著重要的作用,JA途徑中任何階段的缺陷都會引起類似花絲伸長減少、花藥開裂延遲或不開裂的表型[35]。‘Y-14’花絲萎縮、花藥粘連不易開裂,推測可能由于MeJA含量的異常變化以及花蕾發育后期MeJA含量的含量較低,未能及時誘導花絲伸長,花藥開裂,從而出現雄性不育。
ZR是植物體內普遍存在的一種細胞分裂素,其具有順式和反式兩種形式,即T-trans和C-cis,其中反式(TZR)生物活性遠高于順式(CZR)[20]。前人的研究表明植物花藥發育過程中ZR含量過高或過低均可能導致雄性不育的發生。但在本研究中,‘Y-14’和‘陽光玫瑰’的TZR含量普遍偏低(0.05 ng/g左右) 且無顯著差異,就本研究結果來看,TZR含量變化與‘Y-14’雄性不育的形成無直接關系。
梨[36]、葡萄[37]、水稻[38]、辣椒[39]、厚軸茶[40]等作物大量細胞學研究表明,單核期是植物發生雄性不育的關鍵時期。在本研究中,‘陽光玫瑰’花蕾ABA含量在單核早期劇增,而同期‘Y-14’變化平緩;‘Y-14’和‘陽光玫瑰’花蕾SOD活性均在單核早期最低,均在單核晚期劇烈升高。ABA作為應激激素可能在單核早期影響了抗氧化酶尤其是SOD的活性[20-21]。推測在單核早期SOD活性的降低引發了植物自身的應激響應使得ABA含量提升,接著ABA又引導了SOD的重新合成;而由于‘Y-14’花蕾ABA含量變化遲緩,導致SOD活性未能及時提高,加劇了膜脂過氧化,最終引發了雄性不育。另外,‘Y-14’花蕾MeJA含量在花蕾發育前期高于‘陽光玫瑰’,在單核早期開始低于‘陽光玫瑰’且差異愈發明顯,這一變化趨勢與BNS小麥類似[25]。‘Y-14’花蕾GA3含量也在單核早期開始高于‘陽光玫瑰’。因此推測單核早期可能是‘陽光玫瑰’實生后代雄性不育株花蕾生理生化出現異常的關鍵時期,這與厚軸茶相關生理生化研究中的結論一致[7]。
綜上所述,‘陽光玫瑰’實生后代雄性不育株‘Y-14’花蕾的SOD、POD、CAT活性顯著降低,MDA含量增高;其花蕾內源激素ABA、MeJA含量降低,IAA含量增高,GA3含量異常波動,可能與雄性不育的發生有關。‘Y-14’花蕾發育至單核早期,多項指標出現顯著變化,單核早期可能是‘Y-14’生理生化出現異常的關鍵時期。但其中的雄性不育具體機制,還有待進一步研究。
猜你喜歡
中學生天地(A版)(2022年9期)2022-10-31 06:36:28
好日子(2022年3期)2022-06-01 15:58:27
故事作文·高年級(2021年10期)2021-10-23 13:21:26
動漫界·幼教365(中班)(2020年7期)2020-07-14 03:07:21
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
涼山文學(2016年6期)2016-12-05 11:51:42
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
