低溫弱光脅迫對設施黃瓜葉片形態變化的影響
2022-04-25 02:41:58唐衛東劉振文劉冬生
西南農業學報 2022年3期
唐衛東,劉振文,劉冬生
(1.井岡山大學電子與信息工程學院,江西 吉安 343009;2.江西省農作物生長重點實驗室,江西 吉安 343009)
【研究意義】葉片不僅是構成植株冠層結構的重要部分,也是影響農作物生產的重要器官。低溫弱光是溫室越冬和早春季節生產中最具威脅性的逆境之一[1-3],研究表明低溫弱光脅迫對黃瓜等喜溫作物生育期葉片的形態功能影響較大,可導致葉性狀變化出現異常[4-5]。由于葉性狀特征能夠較好地反映作物生長發育狀況及其與外部環境之間的作用關系,從不同角度研究葉性狀特征及其與環境因子的相互關系,以便進一步探索作物生長對環境變化的適應性,是近年來國內外學者廣為關注的問題。【前人研究進展】為了探究葉性狀對外境變化的適應性,人們常根據葉性狀生理特征的變化來探尋滿足作物生長所需的溫光環境等。例如,Pan和Paula等[6-7]探究了基于溫度、光強等環境因子作用的黃瓜葉片光飽和點、最大光合速率及表觀量子效率等變化規律;熊宇等[8]根據不同光照下黃瓜葉片的光合參數和熒光參數,發現遮光使得葉片中葉綠素含量升高,而類胡蘿卜素含量變化不大,且葉片氣孔導度和蒸騰速率均下降,而凈光合速率降低。葉片功能性狀雖然能間接地反映葉器官對外部環境的生理適應性但比較抽象,而葉片形態性狀既是生理功能的外在表現,又能夠直觀地表征葉器官對外部環境變化的適應性。例如,Skupien和Georgios等[9-10]探究了溫度變化對黃瓜葉片光合速率、蒸騰速率以及葉綠素組成結構與含量的影響;陳春宏等[11]從光合有效輻射對黃瓜葉片光合作用、器官發育及干物質累積與分配等方面研究了光照強度對不同生育階段黃瓜葉片的影響規律;李丹丹等[12]探究了光照強度對黃瓜葉片光合產物積累及代謝功能的生理影響。以上相關成果較好地探尋了外部環境因子對葉性狀特征及其變化的作用規律,但是多數以單因素對葉性狀特性的影響研究為主[13-14]。例如,已有研究大部分致力于溫度或光照對葉片形態參數、光合特性等性狀的作用,但實際上溫度往往與光照等其它環境因子共同影響葉性狀特性的變化,而探究多因素對葉性狀及其特性變化的影響尚不夠深入。此外,針對黃瓜葉性狀在逆境下的變化規律,目前大多成果主要集中于探究葉性狀特性及其受外部環境因子影響規律[15-16],而缺乏在多重逆境下針對不同葉序、葉齡葉片與葉片形態性狀特征變化及其相互關系的深入研究。研究表明,與單一逆境脅迫相比,作物在低溫弱光等多重逆境作用下其葉性狀特征及參數之間呈現出一定的相關性[17-19]。例如,常溫下適度的光照不僅有利于黃瓜葉片進行光合作用而且也是確保作物正常生長發育的必要條件,但在低溫下光照將使得光合速率出現下降,并對葉片光合功能造成一定傷害,從而導致葉片形態變化受到抑制[20-21]。【本研究切入點】隨著精細農業及數字農作等技術的快速發展[22-23],深入探索低溫弱光脅迫下葉片性狀特征變化及其相互關系,不僅有助于更好地對作物生長動態進行定量分析與實時監測,還有利于進一步揭示作物-環境互作規律。【擬解決的關鍵問題】本文通過研究低溫弱光脅迫對黃瓜不同葉序葉片形態變化的影響,探尋葉面積、葉干物質量等葉片形態特征對不同程度低溫弱光脅迫的響應規律,既為逆境下作物葉性狀特征的分析和預測提供支持,也為溫室環境管理及優化調控提供參考。
1 材料與方法
1.1 試驗材料
本試驗選用常見的水果型黃瓜品種“碧玉3號”為試材,該品種為全雌無限生長型。通過組培育苗待黃瓜幼苗長至2葉1心時,選取長勢一致的幼苗定植于可控溫室中,緩苗兩周后開始進行低溫弱光脅迫試驗。黃瓜栽培采用枕式袋裝,袋內基質為珍珠巖與蛭石(2∶1配比)。每袋種植4株,栽培密度為2.4株/m2,株行間距均勻。在黃瓜栽培管理時,除植株第3節以下,其余節位均保留一個花芽,及時摘除側枝和卷須。試驗采用營養液滴灌并按照設施無土栽培技術要求進行常規管理。
1.2 試驗設計
試驗于2018年11月至2019年3月分別在4個相同規格的單體可控溫室內進行,每個溫室頂高4.5 m,肩高4 m,寬6 m,長20.0 m。在溫室內采取搭架并在其上覆蓋黑色遮陽網,架高2.5 m。試驗按照溫度變化范圍設置1個對照處理與3個低溫脅迫處理,即:溫度20~25 ℃為適溫生長(CK,即對照處理)、溫度15~20 ℃為輕度低溫脅迫(T1)、溫度10~15 ℃為中度低溫脅迫(T2)、溫度5~10 ℃為重度低溫脅迫(T3)。同時,通過遮陽網對自然光進行遮光處理,并在每個低溫脅迫處理下按照光量子通量密度(Photosynthetic photon flux density,PPFD)變化范圍設置1個對照等級(CL)與3個弱光脅迫等級(L1、L2及L3),即:PPFD值400~600 μmol/(m2·s)為對照等級(CL)、PPFD值200~400 μmol/(m2·s)為輕度弱光脅迫(L1)、PPFD值100~200 μmol/(m2·s)為中度弱光脅迫(L2)、PPFD值0~100 μmol/(m2·s)為重度弱光脅迫(L3)。每個低溫弱光處理15株幼苗,重復3次。CO2的釋放由CO2人工鋼瓶提供,濃度控制在(400±5) μmol/mol范圍內,相對濕度控制在75%。在上述不同處理下分別隨機選定長勢相當的5棵植株作為樣本進行試驗觀測,將處理T1、T2、T3下的試驗觀測數據分別與對照處理CK下的試驗觀測值進行比較分析。
1.3 觀測指標
考慮到黃瓜葉片在生育期內形態變化特性且單葉從出葉到定形大約15~25 d,本文主要對生育期內不同葉序葉片的葉面積、葉干物質量等形態參數變化進行觀測。針對所選定的試驗樣本,依次將不同葉序葉片作為觀測對象。考慮到葉片形態隨主莖上不同葉序葉片分布變化的影響,為了掌握植株不同葉序葉片形態特征變化規律,分別在植株樣本上間隔選取葉序為5、10、15、20、25且葉齡為15的葉片進行監測,并對相應葉序葉片的葉面積、葉干物質量等葉片形態指標進行數據采集。對以上不同處理下的樣本觀測結果取平均值,將其作為黃瓜葉片形態參數觀測值,并用于實驗對比分析。
1.4 數據采集方法
溫室內外的溫度、光照、濕度等環境信息主要通過溫室環境測控系統的相應傳感器獲得。葉片形態結構信息采取破壞性和非破壞性方式并通過常規觀測方法獲得。其中,針對試驗樣本的葉面積采用托普云農葉面積測定儀進行實時監測,并采取非破壞性方式進行測定;而葉片干物質量則是采取破壞性方式進行測定,即先將采摘的新鮮葉片放入干燥箱中,在105 ℃下殺青30 min后,調溫于60 ℃干燥至干質量恒定,冷卻后再用天平稱重。
1.5 相關參數計算公式
為了便于描述不同低溫弱光脅迫對黃瓜葉片形態變化的影響,本文將不同處理下獲得的葉片葉面積、干物質量分別與對照處理CK下的相應指標觀測值進行對比,并依次采用葉面積相對變化率、葉片干物質量相對變化率表示。葉面積相對變化率計算公式為:
(1)
式中,vs為葉面積相對變化率;sC為對照處理CK下的葉片葉面積(cm2);sT為各處理(T1、T2、T3)下的葉片葉面積(cm2)。
葉片干物質量相對變化率計算公式為:
(2)
式中,vg為葉片干物質量相對變化率;gC為對照處理CK下的葉片干物質量(g);gT為各處理(T1、T2、T3)下的葉片干物質量(g)。
1.6 數據處理方法
采用Microsoft Excel 2010記錄數據,SPSS 16.0統計分析數據,采用Microsoft Excel 2010繪圖。
2 結果與分析
2.1 不同部位葉片的形態變化
在處理CK下,從不同葉序的葉片(葉齡為15 d)葉面積實驗觀測結果得知(圖1),處于植株中部位置的葉序為15的葉片葉面積明顯比處于植株兩端的葉序為5、25的葉片葉面積更大,且隨著光量子通量密度(PPFD)的不斷下降,不同葉序葉片的葉面積均呈下降趨勢,其中,越接近植株中下部的葉片葉面積下降尤為顯著,處于植株下部的葉序為5的葉片葉面積下降幅度最大。同時,從不同葉序的葉片干物質量實驗測定結果得知,當PPFD值為400~600 μmol/(m2·s)(CL)時,不同葉序葉片的干物質量總體相差較小。而隨著弱光脅迫的增加,不同葉序葉片的干物質量均呈下降趨勢,其中,越接近植株中上部的葉片干物質量下降尤為顯著,處于植株上部(葉序為25)的葉片干物質量下降幅度最大。
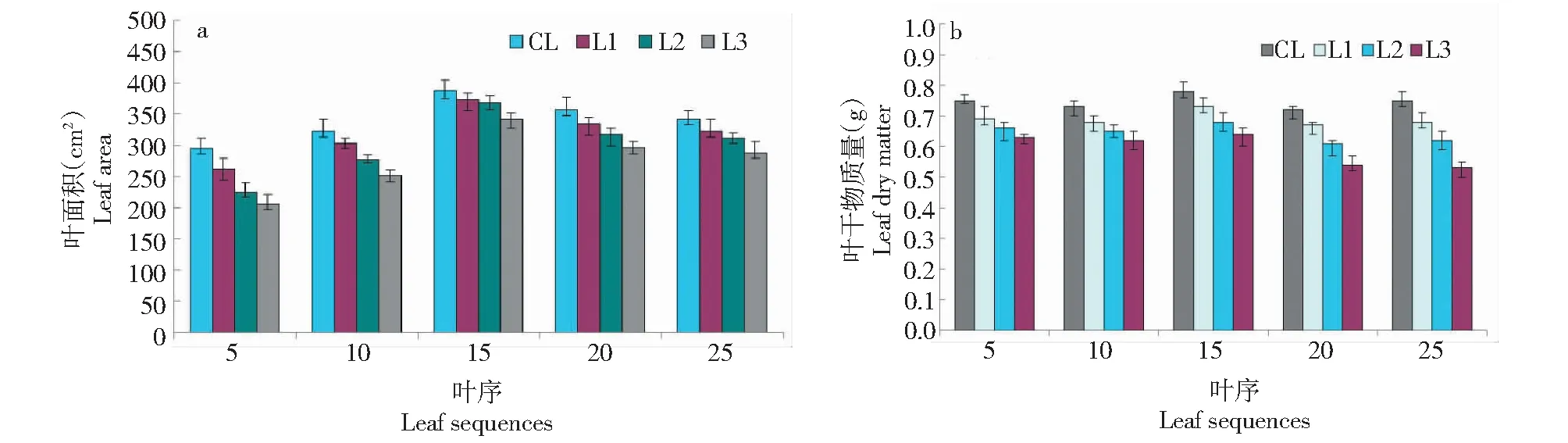
圖1 處理CK下不同葉序葉片的形態變化Fig.1 Morphological changes of different leaf sequences under CK treatment
由試驗觀測結果發現,輕度低溫脅迫下(處理T1),不同葉序的葉片葉面積、干物質量變化規律基本上與處理CK相似(圖2)。
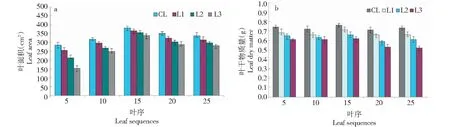
圖2 處理T1下不同葉序葉片的形態變化Fig.2 Morphological changes of different leaf sequences under T1 treatment
在中度低溫脅迫下(處理T2),不同葉序葉片的葉面積與處理CK、T1相比均有一定程度下降(圖3)。處于植株中部位置的葉序為15的葉片葉面積比處于植株兩端的葉序為5、25的葉片葉面積略大。隨著弱光脅迫的增強,植株中下部的葉片相對上部葉片的葉面積下降幅度更大。同時,不同葉序葉片的干物質量與處理CK、T1相比均有一定程度下降。隨著弱光脅迫的加劇,不同葉序葉片的干物質量均呈相應的下降趨勢。
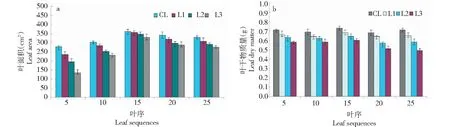
圖3 處理T2下不同葉序葉片的形態變化Fig.3 Morphological changes of different leaf sequences under T2 treatment
當出現重度低溫脅迫(處理T3),不同葉序葉片的葉面積與處理CK、T1、T2相比均有一定程度下降(圖4)。植株上不同葉序葉片葉面積差異不大,而隨著弱光脅迫的不斷加劇,植株各部位葉片的葉面積下降顯著(最大下降幅度為40.3%)。同時,不同葉序葉片的干物質量與處理CK、T1、T2相比均有一定程度下降,且隨著弱光脅迫的加大,植株各部位葉片的干物質量下降幅度顯著增大(最大下降幅度為20.6%)。
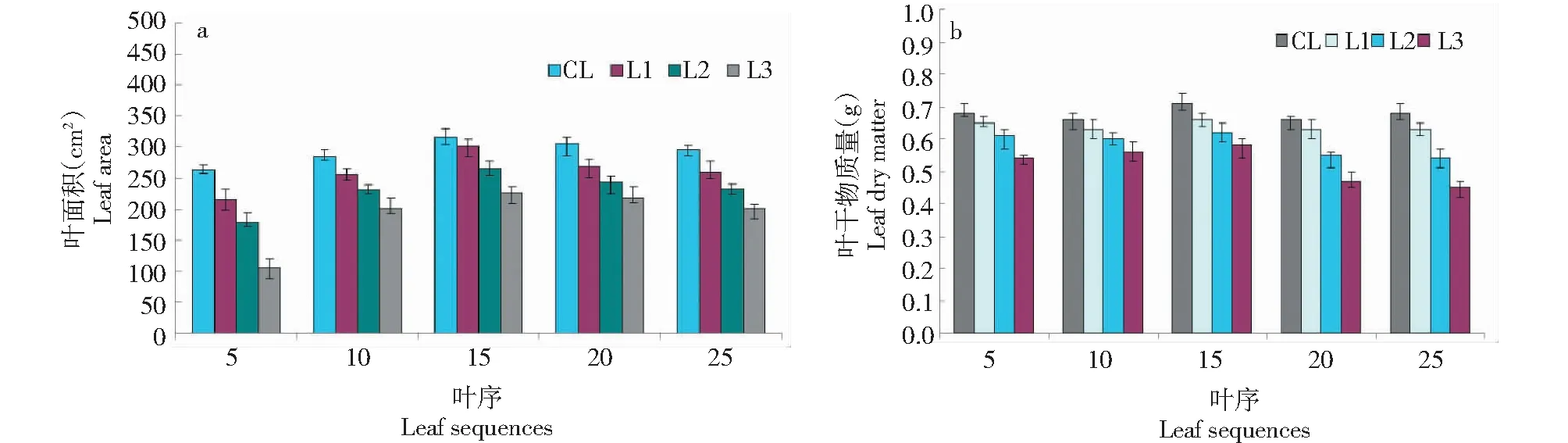
圖4 處理T3下不同葉序葉片的形態變化Fig.4 Morphological changes of different leaf sequences under T3 treatment
2.2 連續低溫對不同部位葉片形態變化的影響
從不同葉序葉片的形態變化規律得知,與對照處理CK相比,從處理T1到T3,隨著低溫脅迫的不斷加劇,葉片葉面積、干物質量都呈下降趨勢。當溫度為10~20 ℃時(輕度、中度低溫脅迫),緊鄰植株中部位置的葉片葉面積明顯比處于植株兩端的葉片葉面積更大;而溫度為5~10 ℃時(重度低溫脅迫),植株上不同葉序位置的葉片形態變化較小且彼此差異不明顯。同時,葉片干物質量與葉面積變化類似。在連續低溫處理下,當弱光等級為L2時不同葉序葉片葉面積及干物質量對比結果(圖5)。這表明分布在植株不同葉序位置的葉片形態隨溫度變化存在一定差異。
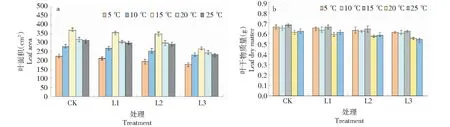
圖5 連續低溫對不同葉序葉片形態變化的影響Fig.5 Effects of continuous low temperature on leaf morphology of different leaf sequences
2.3 與對照處理CK下的葉片形態變化比較
與對照處理CK中不同弱光脅迫下的葉片葉面積相比,輕度低溫脅迫下(處理T1),不同葉序的葉片葉面積均呈相應的下降趨勢,如表1所示。其中,植株中下部的葉片葉面積下降幅度相對較大,尤其是當弱光脅迫達到最大(L3)時,葉序為5的葉片葉面積下降幅度最大(達到25.5%)。同時,與對照處理CK中不同弱光脅迫下的葉片干物質量相比,不同葉序的葉片干物質量也總體呈現對應的下降趨勢,但是下降幅度均不大(小于10%)。
與對照處理CK中不同弱光脅迫下的葉片葉面積相比,中度低溫脅迫下(處理T2),不同葉序的葉片葉面積均呈相應的下降趨勢,如表1所示。其中,植株中下部的葉片葉面積下降幅度逐漸增大,當出現重度弱光(即L3)時,葉序為5的葉片葉面積下降幅度達到30.5%。同時,與對照處理CK中不同弱光脅迫下的葉片干物質量相比,不同葉序的葉片(葉齡為15 d)干物質量也總體呈現對應的下降趨勢,下降幅度相比處理T1有所增加(最大為14.8%)。而且,隨著弱光脅迫的不斷加劇,處于植株中上部的葉片干物質量下降增幅相比植株下部更大。這表明隨著低溫弱光脅迫的加劇,葉片光合同化物出現較大程度的減少,并在植株不同葉序位置的葉片分配上也存在較大差異。
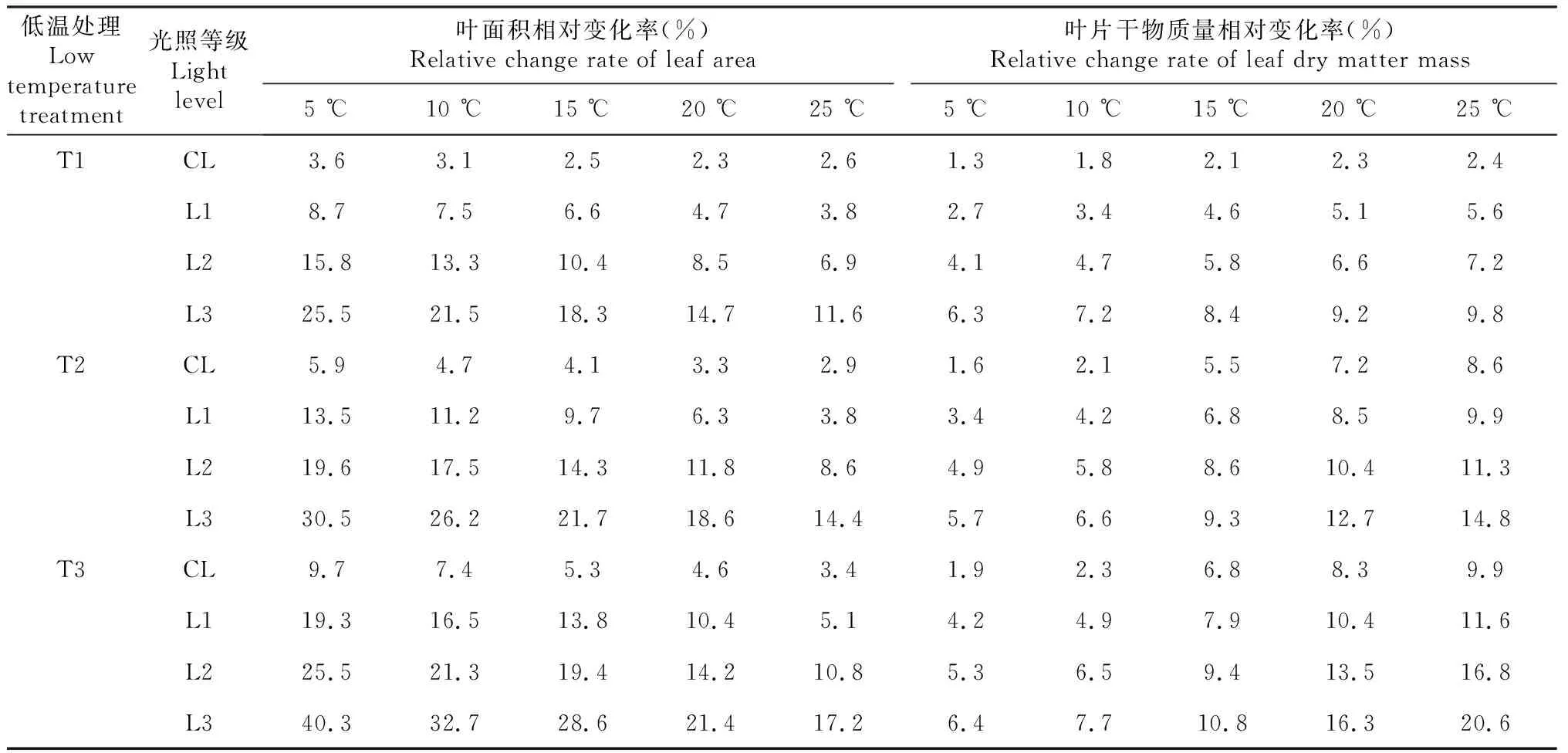
表1 低溫弱光脅迫對不同葉序葉片形態變化的影響
與對照處理CK中不同弱光脅迫下的葉片葉面積相比,重度低溫脅迫下(處理T3),不同葉序的葉片葉面積均呈相應的下降趨勢(表1)。其中,植株中下部的葉片葉面積下降幅度逐漸增大,當弱光脅迫最大(L3)時,葉序為5的葉片葉面積下降幅度達到40.3%。同時,與對照處理CK中不同弱光脅迫下的葉片干物質量相比,不同葉序的葉片干物質量也總體呈現對應的下降趨勢,下降幅度相比處理T2有所增加(最大為20.6%)。而且,隨著弱光脅迫的不斷加劇,處于植株中上部的葉片干物質量下降增幅相比植株下部更大。這表明隨著低溫弱光脅迫的加劇,葉片光合同化物出現較大程度的減少,并在植株不同葉序位置的葉片分配上也存在較大差異。
3 討 論
從不同低溫處理試驗結果得知,在一定溫度范圍內,輕度低溫脅迫對黃瓜植株上不同葉序葉片形態發育基本沒有影響,且靠近植株中部位置的葉片表現出較強的光合生產與同化物轉化能力,從而使得葉面積增長、干物質積累速率均比植株兩端葉片更大。張紅梅等[20]研究發現,過低的溫度(重度低溫脅迫)使得葉片的呼吸作用降低、同化物運輸的有效能量減少。本研究實驗結果也進一步表明,當低溫脅迫由中度轉為重度時,不同葉序葉片葉面積及干物質量增長均出現下降,這表明低溫脅迫的加劇對葉片光合生產及同化物分配能力會產生較大影響。針對連續弱光對不同葉序葉片形態變化的影響,當弱光脅迫不斷加劇(由L1到L3),黃瓜葉片形態經歷了較為復雜的變化。處于植株中下部的葉片由于受到遮光影響而使其光合能力下降,而呼吸作用及同化物轉化能力則得到增強,因此,其葉面積相比植株中上部葉片更小,而干物質量則更大。而且,弱光等級相同時,與葉片干物質量相比,葉面積隨葉序變化更明顯。這表明當PPFD在0~400 μmol/(m2·s)變化時,黃瓜葉片葉面積變化與葉序關系較為顯著,而當PPFD一定時不同葉序葉片干物質量差異較小。
此外,針對不同程度的低溫弱光脅迫對葉片形態變化的影響,LIISA等[24]通過研究得知受植物器官種類及生育期特性等因素影響,葉片在生長過程中對溫度及光照的敏感性及需求水平有所差異。因此,為避免因外部環境脅迫而導致葉片生長出現異常,及時掌握葉片在不同生育期所需的溫度與光照條件對于植物生長及其環境監測至關重要。從本文在低溫弱光脅迫下的黃瓜葉片生長實驗結果中發現,當PPFD大于200 μmol/(m2·s),溫度為15~25 ℃能夠確保不同葉序及葉齡的葉片正常生長,且緊鄰植株中部的葉片長勢更好;而溫度為10~15 ℃時,葉齡小于15 d的不同葉序葉片形態變化較小且彼此差異不大。這表明輕度低溫脅迫基本上不影響葉片的正常生長發育,但當PPFD小于200 μmol/(m2·s),并在中度弱光脅迫(L2)下,溫度過低(中度低溫脅迫)對葉片形態變化有一定影響,此時處于迅速伸展期(葉齡5~15 d)的葉片由于低溫下受到光抑制影響,葉面積增長量明顯減少,而葉片干物質量有所增加,這與前人開展的研究結果基本一致[25]。此外,從以上實驗觀測結果發現,在重度低溫脅迫下,當光照由中度弱光(L2)轉為重度弱光(L3)時,雖然葉片葉面積無明顯變化,但葉片干物質量有所減少。這進一步表明當溫度為5~10 ℃時,PPFD值0~100 μmol/(m2·s)雖然使葉片光合生產能力下降,但并未導致葉片生長代謝過程出現停滯,反而在一定程度上促進了葉片形成的光合產物在不同器官之間的輸送與分配[18]。
4 結 論
低溫弱光脅迫對黃瓜植株葉片的葉面積、干物質量均有不同程度影響,其中,植株中下部的葉片葉面積下降較為顯著,而植株中上部的葉片干物質量下降較為顯著。植株中部的葉片葉面積受輕、中度低溫脅迫影響顯著,而重度低溫脅迫對不同葉序葉片形態變化影響較小且彼此差異不明顯。弱光脅迫導致葉片葉面積隨葉序變化較大,而當光量子通量密度一定時葉片干物質量隨葉序變化較小。雖然低溫弱光脅迫對黃瓜植株中下部葉片形態影響較大,但輕度低溫弱光脅迫不影響黃瓜葉片的正常發育。本研究表明,輕度低溫脅迫對黃瓜植株不同葉序葉片形態發育基本沒有影響,且靠近植株中部位置的葉片表現出較強的光合生產與同化物轉化能力。
