益生菌Nissle 1917鞭毛蛋白高變區隨機缺失及鞭毛展呈外源抗原的構建
2022-04-25 02:50:48金正雨鄧永軍區炳明
西南農業學報 2022年3期
楊 穎,金正雨,鄧永軍,區炳明,文 明
(1.貴州大學動物科學學院,貴陽 550025;2.貴州省動物生物制品工程技術研究中心,貴陽 550025;3.肇慶學院生命科學學院,廣東 肇慶 526061)
【研究意義】大腸桿菌Nissle 1917(EscherichiacoliNissle 1917,EcN)作為益生菌使用已具有100多年的歷史[1],其不含腸毒素、溶血素、細胞毒素等致病因子,無致突變、遺傳毒性及DNA損傷活性[2],能合成細菌素和具有六鐵攝取系統以利于該菌株在腸道中與其他菌株競爭[3],此外,EcN可以利用復雜的機制靶向定殖腫瘤部位進而用于腫瘤靶向治療[4]。因為EcN的益生特性,其一直是德國等國家醫用藥品(Mutaflor?)的活性成分[5],主要用于治療人類消化道菌群失調的相關疾病[6-7]。隨著分子生物學和全基因組測序技術的發展,現已完整注釋EcN的基因組序列[8],基于此,更多EcN基因重組的研究構建出具有新性質的EcN衍生物,這將促進其在未來成為生物活性分子載體或用于口服活疫苗開發的載體系統[9-10]。【前人研究進展】細菌鞭毛表面展呈技術是將外源抗原融合到鞭毛蛋白高變區,隨后構建的嵌合鞭毛組裝成鞭毛絲,進而實現細菌對外源抗原的展呈[11-12]。由于細菌鞭毛絲由成千上萬個鞭毛蛋白單體組裝而成[13],把外源抗原嵌合到鞭毛蛋白高變區,有利于外源抗原的大量表達,并且能利用鞭毛蛋白的佐劑效應[14-15],促進機體對外源抗原的捕獲、加工和提呈。【本研究切入點】EcN具有發達的鞭毛系統,前期研究發現其鞭毛蛋白可以容許10個氨基酸的缺失和6個氨基酸外源抗原的插入[16]。【擬解決的關鍵問題】為尋找外源抗原的其他可能插入位點,并嘗試插入更大片段的外源抗原,選用EcN無質粒菌株(EcN cured of its two cryptic plasmids pMUT1 and pMUT2,EcNc),構建EcNc鞭毛蛋白高變區隨機缺失質粒庫,篩選可缺失位點和插入外源抗原進行展呈,為EcN靶向投遞外源抗原提供操作策略。
1 材料與方法
1.1 材料
1.1.1 菌株、質粒和細胞 供試菌株、質粒和細胞的特征及來源見表1。

表1 供試菌株、質粒和細胞
1.1.2 主要試劑和培養基 限制性內切酶、T4DNA連接酶、Marker購自TaKaRa公司;TransStart FastPfu DNA Polymerase購自北京全式金生物技術有限公司;MutExpress TM ⅡFast Mutagenesis Kit、DNA凝膠回收試劑盒購自南京諾唯贊生物科技有限公司;BAL-31核酸酶購自NEB(北京)有限公司;胰蛋白胨(Tryptone)、酵母提取物(Yeast extract)均購自Oxoid公司;RPMI1640-F12(1∶1)細胞培養基、新生牛血清(NCS)均購自Gibco(美國)公司。
1.2 方法
1.2.1 EcNc fliC高變區隨機缺失質粒庫的構建SpeⅠ為fliC基因中部高變區內唯一的酶切位點,選擇SpeⅠ單酶切pBR322-fliC重組質粒,回收單一線性片段后,用Nuclease BAL-31經不同時間(6、7、10、20 min)消化黏性末端,回收各時間段消化產物,用DNA聚合酶Ⅰ(Klenow)大片段補齊黏性末端后,經T4DNA ligase環化連接,轉化DH5α化學感受態,37 ℃培養1 h后加入10 mL LB(Amp)培養過夜,提取質粒進行瓊脂糖凝膠電泳檢測。
1.2.2 fliC高變區缺失區域的篩選 將fliC高變區隨機缺失質粒庫提取質粒,電轉化EcNc ΔfliC感受態后,涂布LB(Amp)固體培養基,37 ℃培養過夜,挑取單菌落接種半固體運動性平板[17],檢測各菌株運動性能,并設計引物P1: 5′-ATGGCACAAGT CATTAATACCAACA,P2: TTAACCCTGCAGCAGAGACA GA-3′對相應fliC基因進行測序,分析缺失區域。
1.2.3 EcNc FliC高變區展呈外源抗原的構建 設計引物P3: 5′-GCTGGGCAAACTGCTAGT-3′,P4:5′-TGCTACATCACCTGCTTTTGCT-3′,以pUC18-fliC質粒為模板,進行反向PCR擴增缺失fliC基因缺失910~927處的堿基序列;設計引物P5:5′-CAAAAGCAGGTGATGTAGCAAATGCATCTTATGCCGGTGT-3′,P6: 5′-CCACTAGCAGTTTGCCCAGCTACTCTTTGAA TCTGTCCGA-3′, 以F18菌株基因組為模板擴增其FedF基因178~327位堿基序列;上述2種擴增產物回收后混合,在Exnase?催化下,37 ℃反應30 min完成重組反應,實現2個線性化DNA的體外環化,構建pUC18-fliCF(fliCF片段為fliC基因910~927位插入F18FedF基因178~327位堿基序列的重組片段),再將fliCF片段連接到自殺載體pRE112,構建重組自殺質粒pRE112-fliCF并轉化EcNc,參照文獻[18]方法進行重組菌株EcNcfliCF的篩選,并用引物P1、P2進行fliCF序列擴增和測序鑒定。
1.2.4 透射電鏡和運動性觀察 將EcNc與EcNcfliCF過夜培養的菌液分別1∶100轉接于新鮮的LB中,37 ℃震搖培養至OD600nm約為1.0,各細菌均調整為1.0,參照文獻[18]的方法進行電鏡觀察和運動能力檢測。
1.2.5 IPEC-J2 細胞黏附試驗 調整對數生長期的細菌濃度至1×106CFU,以10∶1的比例加入96孔板中的單層IPEC-J2 細胞,在細胞培養箱中共孵育1 h,用PBS洗滌孵育孔3次,去除未黏附的細菌,然后加入0.5%的Triton X-100裂解液,37 ℃作用20 min裂解細胞獲得其表面黏附的細菌,梯度稀釋涂布LB平板,培養過夜后計算黏附細菌數目,利用Graphpad軟件作圖并分析差異顯著性。
2 結果與分析
2.1 fliC高變區隨機缺失質粒庫的構建
提取質粒進行瓊脂糖凝膠電泳,檢測結果(圖1)表明,Nuclease BAL-31經不同時間(6、7、10、20 min)消化EcNcfliC高變區、再經補齊黏性末端環化轉化DH5α化學感受態后,質粒提取物在瓊脂糖凝膠電泳中呈現彌散條帶,表明EcNcfliC高變區出現不同程度的缺失,獲得EcNcfliC高變區隨機缺失質粒庫。
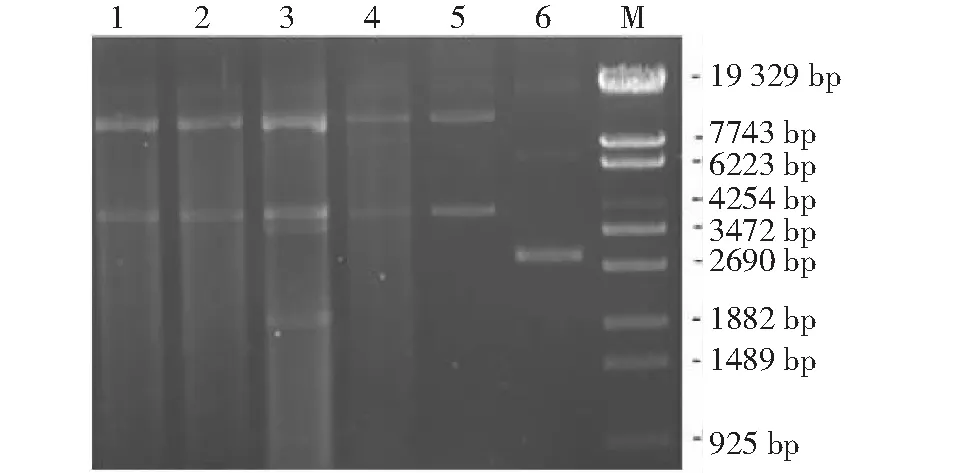
M: λ-EcoT14 I digest; 1~4: 經Nuclease BAL-31分別消化6、7、10、20 min構建的質粒庫; 5: pBR322-fliC; 6: pBR322M: λ-EcoT14 I digest; 1-4: The plasmids library is constructed by digestion of Nuclease BAL-31 for 6, 7, 10 and 20 minutes respectively; 5: pBR322-fliC; 6: pBR322圖1 EcNc fliC高變區隨機缺失質粒庫鑒定結果Fig.1 Identification of random deletion plasmid library in hypervariable region of EcNc fliC
2.2 fliC基因高變區缺失區域的篩選
從圖2看出,EcNcfliC高變區隨機缺失質粒庫電轉化EcNc ΔfliC感受態后,挑取過夜培養的單菌落接種半固體運動性平板,運動性檢測顯示,大部分菌株未呈運動性(圖2-A,2-B);擴增相應fliC序列測序結果顯示,大部分高變區隨機缺失的區域為非3的整數倍缺失(圖3);試驗篩選到一株fliC高變區缺失菌株具有運動性(圖2-C),經測序其fliC基因缺失910~927 bp處堿基序列(圖3)。
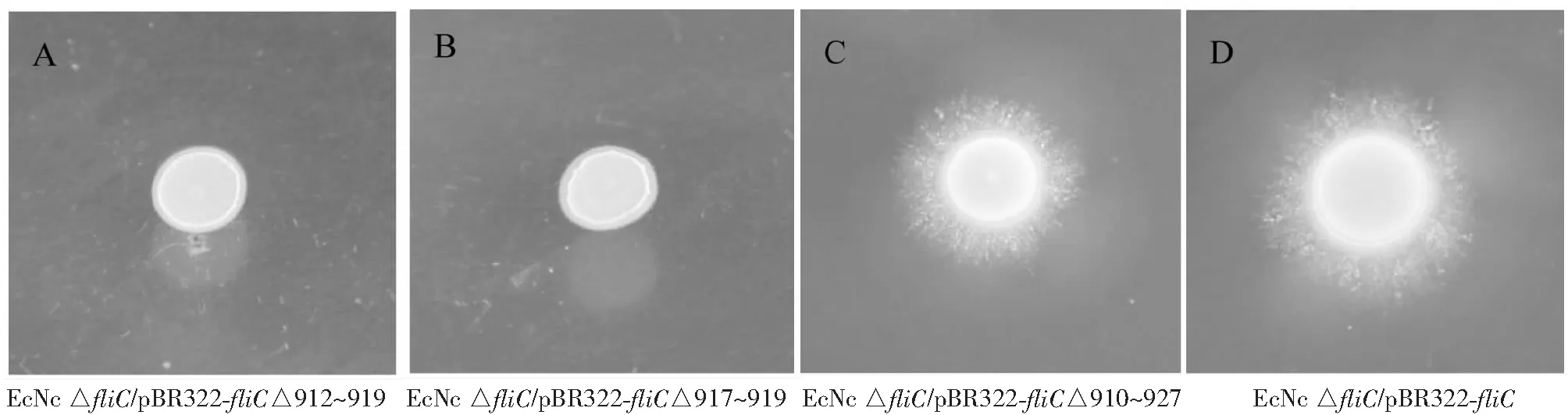
圖2 部分菌株運動能力檢測結果Fig.2 Motility detection of some EcNc strains

圖3 部分EcNc fliC高變區缺失區域Fig.3 Hypervariable region deleted area of fliC of some EcNc strains
2.3 重組菌株EcNcfliCF的構建
利用 MutExpress TM ⅡFast Mutagenesis Kit,構建fliC基因910~927位嵌合F18 FedF基因178~327位堿基序列的重組片段fliCF,將fliCF連接入自殺載體pRE112中轉化到EcNc,2次重組后經PCR和測序鑒定,結果顯示,重組菌EcNcfliCF構建正確。
2.4 重組菌株EcNcfliCF表型的檢測
從圖4可知,EcNcfliCF重組菌株在電鏡下能觀察到鞭毛絲,但在運動平板上無運動性能。
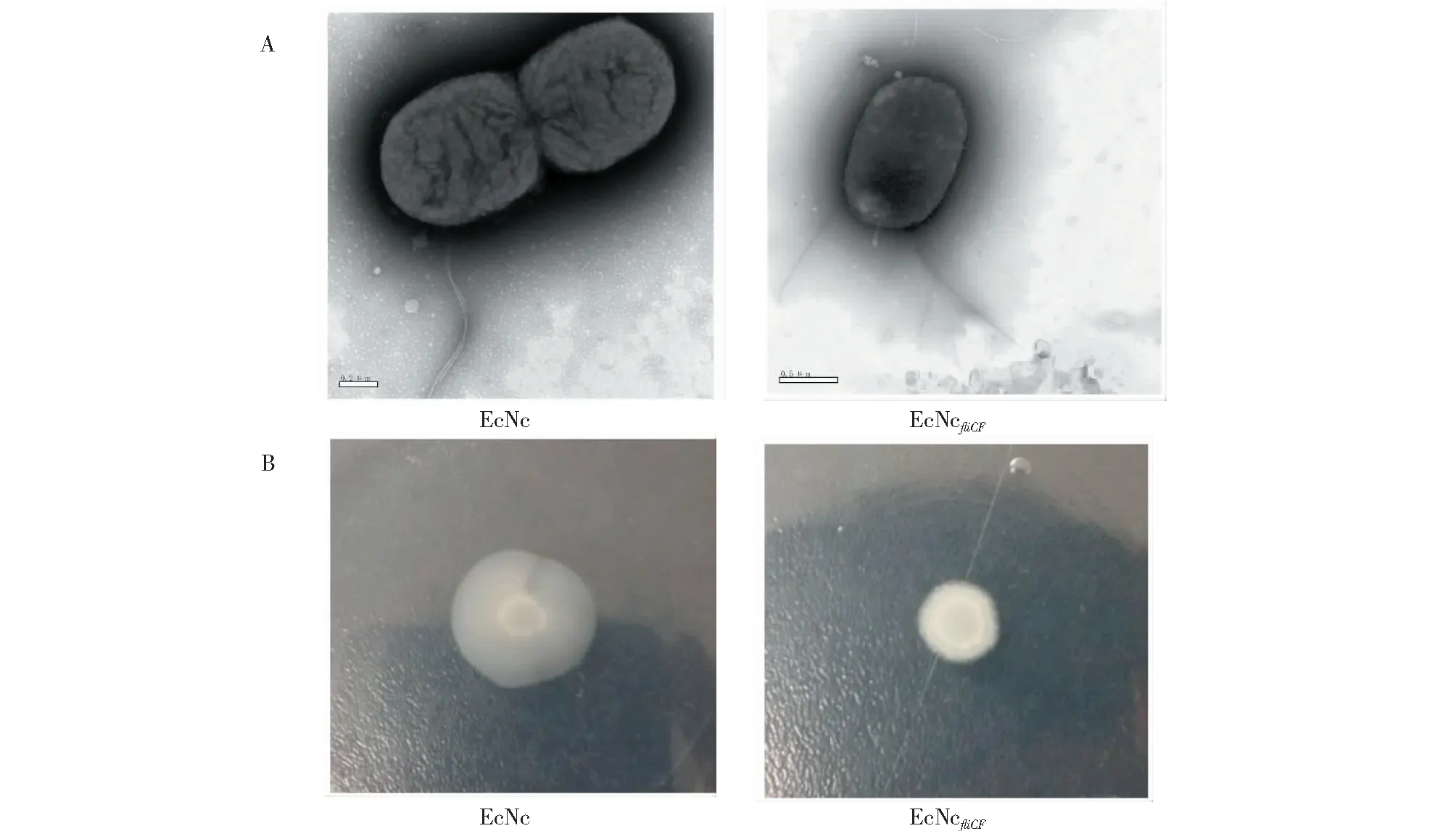
圖4 重組菌株EcNc和EcNcfliCF的表型Fig.4 Phenotypes of EcNc and EcNcfliCF
2.5 IPEC-J2細胞黏附檢測
從圖5可知,相對EcNc,EcNcfliCF對 IPEC-J2細胞的黏附能力減少約0.28%,差異不顯著(P>0.05)。
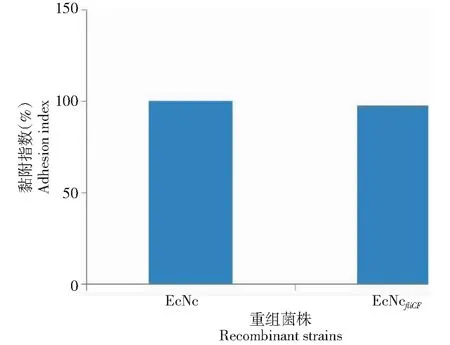
圖5 重組菌株對IPEC-J2的黏附指數(EcNc的黏附指數假設為100%)Fig.5 Bacterial adherence to IPEC-J2 cells (EcNc strain adhesion index is assumed as 100%)
3 討 論
益生菌通過不同機制預防和控制胃腸道致病菌感染,改善動物生產性能和產量,作為抗生素促進劑的替代物越來越受到關注。自Alfred Nissle發現攝入特定的大腸桿菌菌株可以治療感染性疾病患者起,EcN菌株作為益生菌被應用于防治人類疾病。楊穎等[19-22]從豬糞便中分離到EcN,其能持續在豬體內定殖,該菌株可以保護豬宿主免受產毒素大腸桿菌的侵害,EcN對機體的保護作用有賴于其F1C菌毛和鞭毛介導的對有害菌株的黏附抑制[22]。
EcN鞭毛絲含有大量鞭毛蛋白單體,在前期研究中發現可利用鞭毛蛋白展呈技術將外源抗原展呈于EcN表面,重組EcN菌株定殖于機體發揮益生作用的同時,還可誘導機體產生針對外源抗原的體液及黏膜免疫應答[23]。但是不同血清型的細菌,其鞭毛蛋白尤其是高變區氨基酸差異極大,使用鞭毛蛋白展呈技術需要確定較好的外源抗原插入位點,進而優化外源抗原的展呈效果。故本研究基于鞭毛蛋白構建重組質粒,產生高變區隨機缺失的質粒庫,篩選鞭毛蛋白可缺失的區域(fliC基因缺失910~927 bp處堿基序列),該區域缺失后,并不影響菌株鞭毛絲的形成和運動能力。EcN含有2種隱性質粒,分別命名為pMUT1和pMUT2,前期研究中,已構建無質粒的EcN變體(EcNc),其生物學特征與野生型EcN菌株沒有區別,可以更好地用作基因工程活載體菌[24]。
EcNc已被證明可以通過其鞭毛系統展呈6His標簽[16]。EcNc鞭毛蛋白高變區較長,位于195~320 aa處。本研究基于篩選到的EcNc鞭毛蛋白可缺失區域(910~927 bp處堿基序列),在該缺失區域處插入F18大腸桿菌黏附素FedF亞單位與仔豬小腸上皮細胞相互作用的受體結合域(60~109位氨基酸殘基區段),成功將FedF亞單位受體結合域的50個氨基酸展呈于EcNc表面。構建的重組菌株EcNcfliCF能夠形成鞭毛絲,雖然不具備運動能力,但不影響其對IPEC-J2細胞的黏附能力;且在EcNc鞭毛蛋白高變區插入50個氨基酸,與6His標簽的長度比較有了顯著的容納性,大大提升插入容量。
4 結 論
研究成功將FedF亞單位受體結合域的50個氨基酸展呈于EcNc表面,這為后期進一步篩選鞭毛蛋白高變區其他缺失區域,和基于已篩選位點插入展呈其他外源抗原,優化構建遺傳穩定、定植力強的功能性益生菌打下基礎。
