不同發育階段杉木人工林凋落物能量的輸入特征
2022-04-26 06:04:44周麗麗李樹斌蔡麗平吳鵬飛馬祥慶
江西農業學報 2022年2期
周麗麗,黃 鵬,李樹斌,蔡麗平,吳鵬飛,馬祥慶*
(1.閩江學院 海洋研究院,福建 福州 350108;2.福建農林大學 材料工程學院,福建 福州 350002;3.福建農林大學 林學院,福建 福州 350002;4.國家林業和草原局 杉木工程技術研究中心,福建 福州 350002)
森林凋落物是森林植物在其生長發育過程中新陳代謝的產物,是連接森林與土壤、大氣的重要“紐帶”,也是森林植被與環境之間進行物質和能量交換的主要途徑,在森林生態系統養分循環和能量流動中起重要作用[1]。自德國科學家Ebermayer于1876年對凋落物在森林生態系統養分循環中的作用進行了系統闡述以來,國外許多學者先后對不同類型、不同地區的森林凋落物進行了研究[2]。我國對森林凋落物的研究起步較晚,始于20世紀60年代,先后開展了多種森林類型凋落物產量、化學組成及分解規律的研究。然而目前國內外關于森林生態系統能量流的研究還很薄弱[3]。植物通過光合作用將能量儲存于各器官中,在參與自身代謝的同時,通過凋落物分解將能量輸入給林地,進一步驅動土壤微生物活動和土壤有機碳礦化[4]。因此,對森林凋落物的能量輸入特征進行研究,對揭示森林生態系統太陽能的轉化效率及能流轉化規律具有重要的理論及實踐意義[5]。
杉木[Cunninghamia lanceolata (Lamb.) Hook]是中國南方重要的速生豐產林樹種,其栽植面積廣,栽植歷史悠久,在我國人工林生產中具有舉足輕重的地位[6]。國內關于杉木凋落物的研究一直是熱點,通過CNKI檢索到相關科技文獻818篇,主要集中在產量及季節動態、分解及養分釋放、碳儲量等方面[7-8]。目前,關于杉木凋落物能量輸入林地過程的研究還比較少,僅見楊玉盛[9]、陳光水[10]、郭劍芬[11]等對杉木純林與杉木+觀光木混交林凋落物能量歸還季節動態的報道,而關于林齡對杉木人工林凋落物能量分配和輸入動態影響的研究還鮮見報道,因此,加強杉木人工林生態系統凋落物能量輸入特征的研究,對深入理解杉木人工林各生態過程、完善杉木人工林生態系統能流體系具有重要意義。鑒于此,筆者以本課題組前期建立的福建三明莘口教學林場不同發育階段(幼齡林、成齡林和過熟林)的杉木人工林為研究對象,研究了不同發育階段杉木林凋落物各組分的熱值差異,比較了不同發育階段杉木林凋落物的能量輸入特征及季節動態,以期為豐富杉木人工林能流理論及科學經營杉木人工林提供理論及技術指導。
1 試驗地概況與研究方法
1.1 試驗地概況
試驗地位于福建省三明市福建農林大學莘口教學林場(26o10′N,117o27′E),氣候類型屬亞熱帶季風性氣候,水熱資源豐富,季風明顯,年平均氣溫為19.1 ℃,年均降水量為1749.0 mm,年均蒸發量為1585.0 mm,≥10 ℃年積溫為5240.2~ 6677.9 ℃·d,全年日照時數為1840 h,無霜期為300 d左右。試驗地海拔205~500 m,土壤類型為山地紅壤,成土母質以粉砂頁巖為主。3種杉木林分林下植被種類相似,灌木層主要有大葉紫珠(Callicarpa kochiana)、杜莖山(Maesa japonica)、毛冬青(Ilex pubescens)和粗葉榕(Ficus hirta)等,草本層主要有狗脊(Woodwardia japonica)、江南卷柏(Selaginella moellendor)、山姜(Alpinia japonica)和觀音座蓮(Angiopteris fokiensis)等,藤本植被種類較少,包括網絡崖豆藤(Millettia reticulata)、玉葉金花(Mussaenda pubescens)、網脈葉酸藤果(Embelia rudis)、顯齒蛇葡萄(Ampelopsis grossedentata)等。3種杉木林分的基本情況見表1。
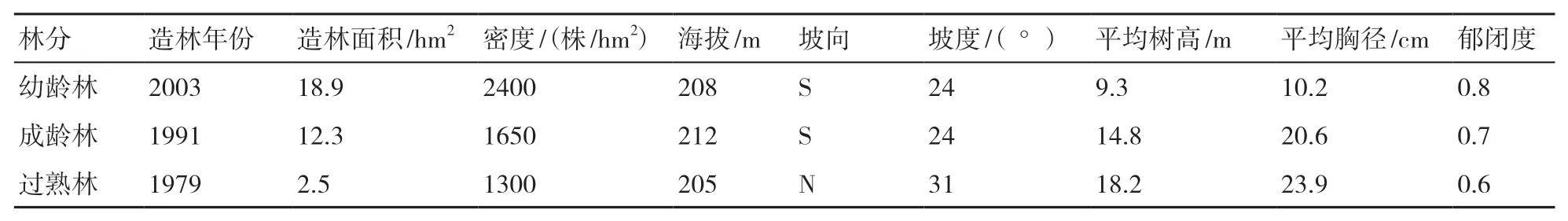
表1 3種杉木林分的基本情況
1.2 研究方法
1.2.1 凋落物的收集 在每種林分內分別設立3個20 m×20 m的標準地,在每個標準地內隨機布置10個1 m×1 m的收集器,布設高度為50 cm,收集器的尼龍網孔徑為1 mm。2012年3月~2013年2月每月中旬對凋落物進行收集。
1.2.2 凋落物的處理 將每個收集器內的凋落物按葉、枝(包括枝皮)、花果(包括脫落的種子)和其他組分(主要指林地內其他樹種、林下植被的凋落物,以及蟲鳥糞、蛹、小動物殘體等)進行分類,稱鮮重;隨后立即置于80 ℃恒溫干燥箱內烘干至恒重,再稱干重,用于計算各組分的凋落物產量(g/m2)。將同一標準地相同組分的凋落物混合,將樣品磨碎,過60目篩,貯存于廣口瓶中,備用。
1.2.3 凋落物能值的測定 采用武漢生產的GR-3500型恒溫式氧彈熱量計測定凋落物各組分的干重熱值(Gross Caloric Value,簡稱GCV),在室內恒溫10 ℃條件下進行測定,對每個樣品重復測定2~3次,重復間的相對誤差控制在±1%以內。采用干灰化法測定樣品的灰分含量;計算去灰分熱值(Ash-free Caloric Value,簡稱AFCV),計算公式為AFCV= GCV/(1-灰分含量)[10]。為了比較不同季節AFCV的動態特征,將3~5月、6~8月、9~11月及12月~次年2月分別計作春、夏、秋、冬四季,將同一標準地內同一季節相同組分的凋落物混合,分別測定不同組分的AFCV;不同季節的能量輸入特征采用AFCV值與不同組分凋落物產量的乘積來表示。
1.3 數據處理
發育階段、凋落物組成、季節及其交互效應對灰分含量、GCV和AFCV的影響采用三因素方差分析法進行分析,以發育階段、凋落物組成及季節為固定因子;發育階段、季節及其交互效應對能量輸入的影響采用雙因素方差分析法進行分析,以發育階段和季節為固定因子;對同一組分不同發育階段之間、同一發育階段不同組分之間的灰分含量、GCV及AFCV差異采用單因素方差分析法進行分析;對同一季節不同發育階段之間、同一發育階段不同季節之間的AFCV值及能量輸入量差異采用單因素方差分析法進行分析。差異多重比較采用最小顯著差異法(Least Significant Difference, LSD)。數據統計分析軟件和圖表制作軟件分別為SPSS 17.0和Excel 2007。
2 結果與分析
2.1 不同發育階段杉木人工林凋落物各組分的熱值特征
三因素方差分析結果表明:不同發育階段、不同凋落物組成及不同季節的灰分含量差異極顯著(P<0.001);發育階段、凋落物組成、季節及其兩者之間的交互效應對凋落物干重熱值(GCV)及去灰分熱值(AFCV)的影響均達到了極顯著水平(P<0.001)(表2)。從平均值來看,不同組分的灰分含量變異較大,表現為其他組分最高(5.73%~8.59%),其次為花果和葉,以落枝最低(3.73%~5.46%)(圖1a)。不同組分的GCV和AFCV值均表現為葉和花果較高,其他組分次之,以落枝最低(圖1b、圖1c)。植物灰分含量對GCV和AFCV的測定具有直接影響,研究發現,過熟林凋落花果的AFCV值高于凋落葉的,但由于其花果的灰分含量較高,導致落花果的GCV值低于凋落葉的,因此,選擇AFCV作為衡量植物熱值差異的指標更精確。
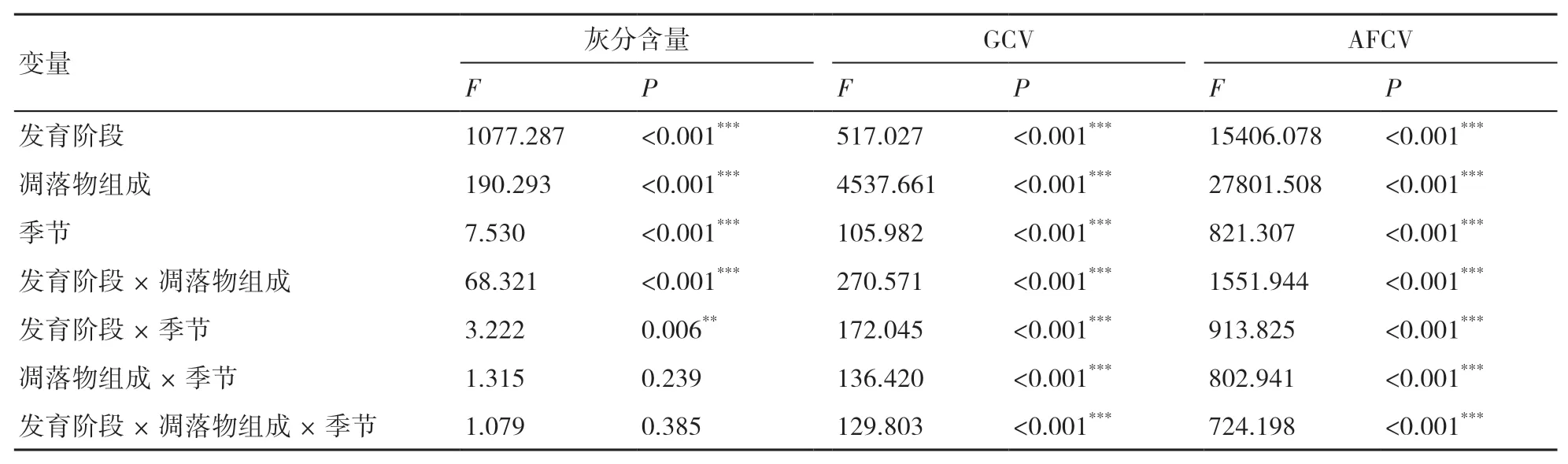
表2 發育階段、凋落物組成、季節及其交互效應對灰分含量、GCV和AFCV的影響
不同發育階段不同組分的灰分含量介于3.51% ~8.59%,GCV和AFCV值分別介于18.14~ 21.19 kJ/g和18.80~22.55 kJ/g(圖1)。單因素方差分析結果表明:同一組分不同發育階段杉木林凋落物的灰分含量差異顯著(P<0.05),均表現為過熟林>成齡林>幼齡林;不同發育階段杉木林凋落物的GCV值表現規律不一,而凋落葉、枝和其他組分的AFCV值表現出與灰分含量相同的規律,即隨杉木發育階段的推進呈增加趨勢。灰分含量與營養元素的富集量及狀態有關[11]。杉木幼齡林凋落葉的灰分含量較低,可能與葉片凋落前較高的養分轉移有關,有報道指出K素的轉移率以幼齡林最高(81.23%),其次為成齡林(78.11%),以過熟林最低(71.34%)[12]。
2.2 不同發育階段杉木林凋落物各組分的AFCV值的季節動態
由圖2可知,各季節杉木林凋落葉、枝、花果和其他組分AFCV在不同發育階段之間的差異均達到了顯著水平(P<0.05),其中在春季、秋季和冬季各組分的AFCV均表現為過熟林>成齡林>幼齡林。不同季節杉木幼齡林凋落物葉的AFCV值表現為夏季>秋季>春季>冬季,成齡林表現為秋季>春季>夏季>冬季,過熟林表現為秋季>冬季>夏季>春季(圖2a)。落枝凋落物的AFCV以過熟林最高;但是落枝凋落物AFCV的季節動態隨杉木發育階段的推進而不同,其中幼齡林表現為冬季最高、秋季最低,而成齡林與過熟林均表現為秋季和冬季較高、夏季最低(圖2b)。幼齡林和過熟林落花果的AFCV季節變化表現為秋季最低,而成齡林則表現為夏季最低、秋季最高(圖2c)。幼齡林和成齡林其他凋落物組分的AFCV季節變化表現為冬季最高、春季最低,而過熟林則表現為冬季最高、夏季最低(圖2d)。
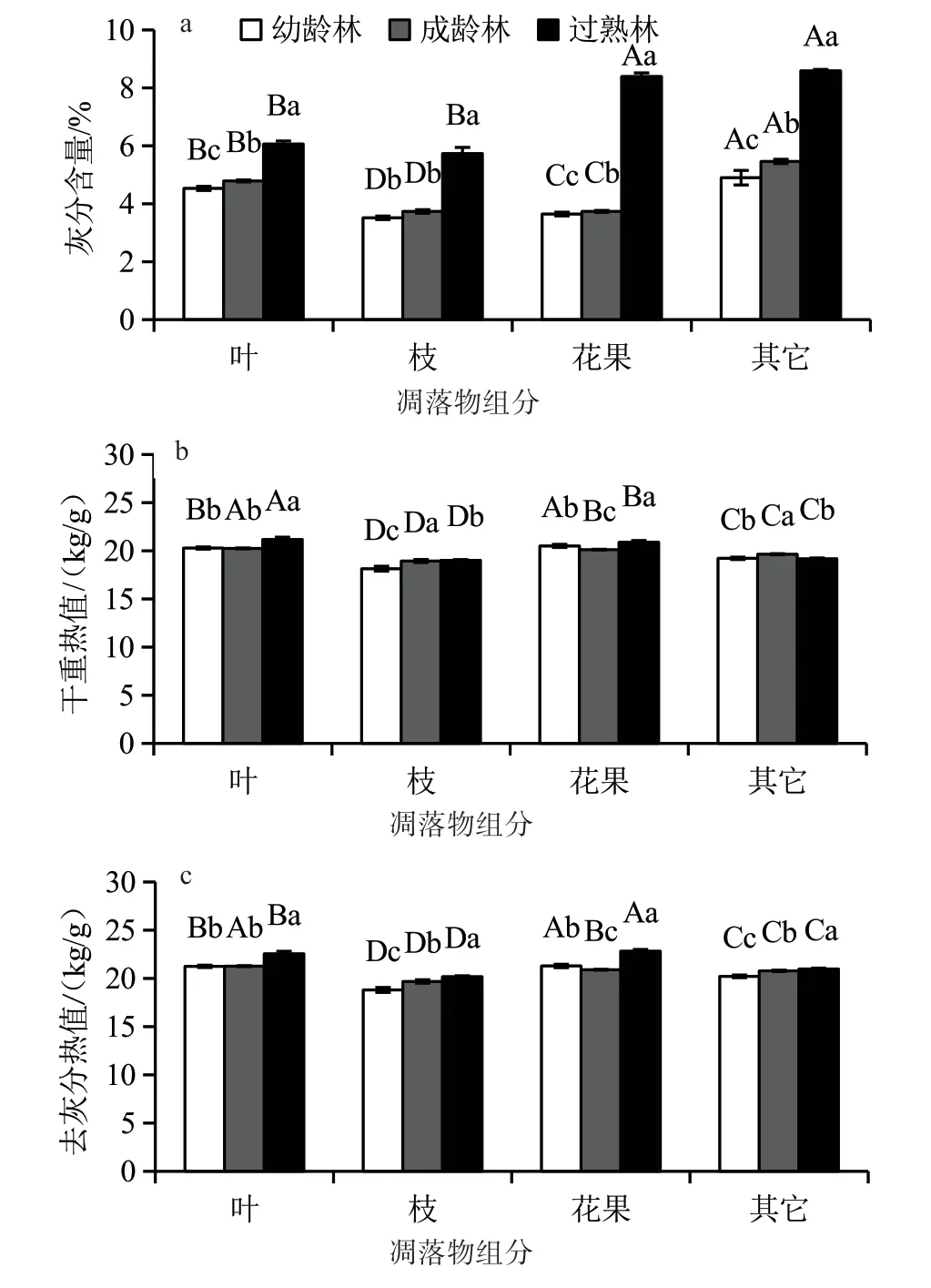
圖1 不同發育階段杉木人工林凋落物各組分的灰分含量、GCV和AFCV值
2.3 不同發育階段杉木林凋落物能量輸入的季節動態及年輸入總量
采用凋落物產量與AFCV值的乘積來表示不同組分凋落物能量的輸入[16],這部分能量對驅動土壤微生物、有機質礦化等生態過程具有重要作用。雙因素方差分析結果(表3)表明:除發育階段對凋落葉和其他組分,以及發育階段×季節交互效應對凋落物花果能量輸入的影響不顯著外,發育階段、季節及其交互效應對杉木凋落物各組分能量輸入的影響均達到了顯著水平(P<0.05)。
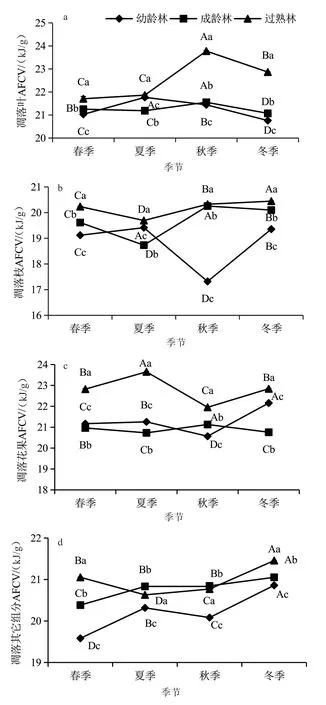
圖2 不同發育階段杉木人工林凋落物各組分的AFCV值的季節動態
由圖3可見,不同發育階段杉木林凋落物各組分及總量的能量輸入具有明顯的季節動態變化特征,總體表現為春季和夏季的能量輸入較高,這2個季節的能量輸入量占年總量的68.72%~75.52%。以凋落葉和枝為例,幼齡林凋落葉的能量輸入表現為春季>夏季>秋季>冬季,成齡林表現為冬季>春季>夏季>秋季,過熟林表現為春季>夏季>冬季>秋季(圖3a);幼齡林與過熟林凋落枝的能量輸入均表現為春季>夏季>冬季>秋季,成齡林則表現為春季>冬季>夏季>秋季(3b)。
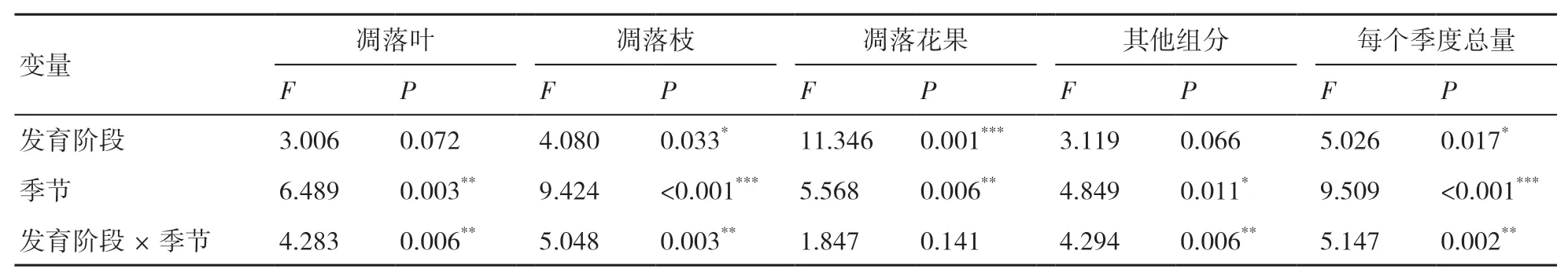
表3 發育階段、季節及其交互效應對杉木凋落物能量輸入的影響
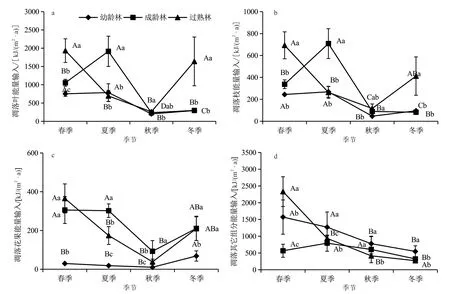
圖3 不同發育階段杉木人工林凋落物各組分的能量輸入的季節動態
從表4可以看出,3種林分凋落物的年能量輸入量表現為過熟林[10755.75 kJ/(m2?a)]>成齡林[7924.80 kJ/(m2?a)]>幼齡林[6987.46 kJ/(m2?a)],差異均達顯著水平(P<0.05)。各組分的年能量輸入量在不同發育階段差異顯著,凋落葉和枝的年能量輸入量表現為過熟林>成齡林>幼齡林,而其他組分則表現為幼齡林>過熟林>成齡林。杉木各發育階段不同凋落物組分的年能量輸入量差異顯著(P<0.05),幼齡林表現為其他組分>凋落葉>凋落枝>凋落花果,成齡林與過熟林均表現為凋落葉>其他組分>凋落枝>凋落花果,可見在各組分中凋落葉及其他組分的能量輸入量較大,分別占總能量輸入量的29.17%~44.23%和28.90%~59.65%,其次為凋落枝(9.38%~15.37%),而凋落花果所占比例最小(1.81%~11.50%)。
3 結論與討論
灰分含量可以反映植物對養分選擇吸收和積累的特點,其因植物種類、器官及發育階段等不同而差別較大,直接影響植物AFCV的測定;一般認為灰分含量與干重熱值呈負相關,與去灰分熱值的相關性不確定[13]。去灰分熱值是除去灰分后的熱值,更能準確地反映單位干物質所含的能量,比干重熱值更具可比性。不同器官、不同樹種、不同季節植物的AFCV值都會有所不同,一般認為葉和花果的AFCV值較高,枝的AFCV值最低,這主要與各器官的功能及結構不同有關。葉作為光合作用的重要器官,其含有較多的蛋白質等高能量有機物;花果作為重要的生殖器官,也富集了大量的蛋白質和脂肪,其熱值也較高;而落枝主要由木質纖維等低值有機物組成,故熱值較低[14]。在本研究中,不同發育階段杉木凋落物的AFCV值也以葉和花果較高,其他組分次之,以落枝最低。另外,本研究發現,杉木各凋落物組分的灰分含量、干重熱值和去灰分熱值隨杉木發育階段的推進而普遍呈增加趨勢,這與張貴等[15]對湖南會同第2代不同林齡(7、11、14、18和20年)杉木林凋落物能值的研究結果一致,說明伴隨杉木生長的進行,杉木凋落物各組分化學組成的改變直接影響其能值。
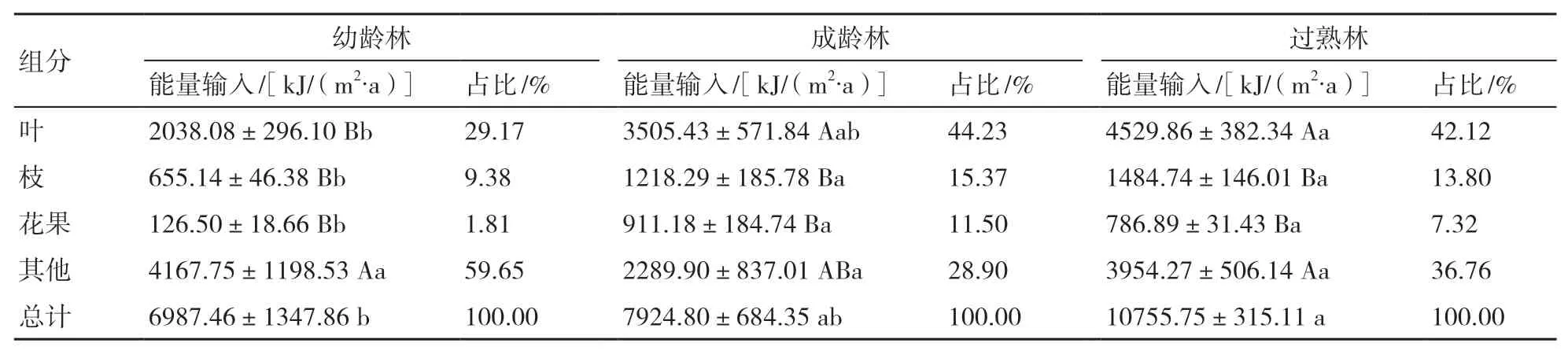
表4 不同發育階段杉木林凋落物各組分的年能量輸入量及其所占比例
與熱帶及溫帶地區植物凋落物的熱值相比,本研究中不同發育階段杉木人工林各組分的AFCV值(18.80~22.55 kJ/g)與福建南靖縣和溪鎮亞熱 帶 雨 林 凋 落 物 的AFCV值(20.93~22.64 kJ/g)相似,也與福建三明32年生福建柏林的AFCV值(22.05~23.41 kJ/g)[16]和福建建甌25年生馬尾松林的AFCV值(21.24~22.22 kJ/g)[17]相似,但明顯低于亞馬遜地區熱帶雨林的(24.80 kJ/g)[18]。亞馬遜地區植物的高熱值現象與土壤貧瘠導致光合作用產生高能的化合物如蠟、樹脂和脂肪有關。另外,本研究結果顯示,杉木凋落物各組分去灰分熱值的變化具有明顯的季節節律,其中凋落葉的AFCV在夏、秋2個季節較高,而在冬、春季較低,這是由于夏季溫度升高,太陽輻射增強,導致葉的光合作用旺盛,體內多糖類物質積累,引起葉內熱值的升高;秋季正好是杉木生長的高峰,葉的光合作用仍較旺盛,致使含能量物質在葉內積累,能值處于較高水平;而在冬季和春季,葉片的光合作用微弱,同時,葉內所積累的糖類物質被不斷消耗掉,因而其熱值處于較低水平。
本研究杉木過熟林凋落物的年能量輸入量[10755.75 kJ/(m2?a)]與福建建甌萬木林自然保護區25年生杉木林的[11377 kJ/(m2?a)]相似[17],但低于福建三明相似林齡(27年生)杉木純林的[12648 kJ/(m2?a)][9]。本研究杉木成齡林凋落物的年能量輸入量[7924.80 kJ/(m2?a)]與武夷山甜櫧林的[7679.71 kJ/(m2?a)]相似[19],但明顯低于25年生馬尾松人工林的[13734.47 kJ/(m2?a)][17]。本研究發現,凋落物的能量輸入量的規律與凋落物產量的基本一致,因此,一切影響凋落物產量的因素如自身生物學特性、外界氣候因子等均會影響凋落物能量的輸入及轉化,進而影響林分的微生物活動、土壤養分循環及土壤碳礦化等功能過程[1]。杉木林分凋落物的能量輸入具有明顯的季節動態,以春、夏2季輸入量較高,秋季和冬季輸入量較小,這與謝梅芳[17]對福建建甌25年生杉木人工林凋落物的能量輸入結果相似。凋落物能量輸入的季節變化與凋落物產量的季節動態密切相關,本課題組研究發現,杉木凋落物產量在1年中有2次峰值,分別出現在4月和8月,因此,春季和夏季凋落物的能量輸入量較大[20]。另外,從凋落物產量來看,杉木落葉占總凋落物的絕大部分(27.5%~43.6%),落花果所占比例很小(0.8%~10.5%)[1],因此,凋落葉組分對于杉木生態系統能量的輸入意義更大,是土壤微生物能量的主要來源。
為了進一步比較不同類型植物群落能流的差異,計算了1年中太陽輻射能進入凋落物的轉化效率(即1年中凋落物能流量占林地太陽有效輻射能的百分數),依據福建三明當地的太陽有效輻射2251206 kJ/(m2?a)[17]來計算,不同發育階段杉木林凋落物的太陽能轉化率表現為過熟林(0.48%)>成 齡 林(0.35%)>幼 齡 林(0.31%),低 于25年生馬尾松林的(0.61%)[17]、福建南靖縣和溪鎮亞熱帶雨林的(0.81%)[21],以及福建紅樹林群落的(0.88%)[23],但明顯高于武夷山甜櫧林群落的(0.17%)[19]和科特迪瓦熱帶雨林的(0.21%)[23],說明福建省三明市杉木林的凋落物具有較高的太陽能轉化效率,并且隨發育階段的推進而增加,因此,適當延長杉木的輪伐期有利于增加林地的能量流輸入以及提高太陽能的利用效率。
