獼猴桃AdMSD1和AdMSD2的克隆及其在果實后熟軟化中的表達
2022-05-06 03:25:30賴瑞聯(lián)高敏霞吳如健陳義挺
核農(nóng)學(xué)報 2022年5期
關(guān)鍵詞:植物
馮 新 賴瑞聯(lián) 高敏霞 吳如健 陳義挺,*
(1 福建省農(nóng)業(yè)科學(xué)院果樹研究所,福建 福州 350013;2 福建省落葉果樹工程技術(shù)研究中心,福建 福州 350013)
植物細胞內(nèi)活性氧產(chǎn)生與消除的動態(tài)平衡,是保證植物正常生長發(fā)育的重要基礎(chǔ)。逆境或衰老等過程通常會導(dǎo)致活性氧的大量積累,產(chǎn)生氧化脅迫,加速植物的衰老死亡[1]。超氧化物歧化酶(superoxide dismutase,SOD)是植物活性氧清除系統(tǒng)的第一道抗氧化防線,主要負責(zé)清除細胞內(nèi)產(chǎn)生的過量超氧陰離子自由基,保護細胞膜免受傷害,進而提高抗氧化能力[1]。根據(jù)輔基中金屬離子的差異,植物SOD主要分為錳超氧化物歧化酶(manganese-SOD,MnSOD)、銅鋅超氧化物歧化酶(copper/zinc-SOD,Cu/ZnSOD)和鐵超氧化物歧化酶(iron-SOD,F(xiàn)eSOD)[2]。MnSOD作為SOD家族的重要一員,主要定位于線粒體,由核基因編碼,其基因序列已陸續(xù)從落葉松[3]、芥菜[4]、香蕉[5]、丹參[6]、龍眼[7]、小麥[8]和非洲菊[9]等多種裸子植物和被子植物中鑒定得到。
諸多研究表明,MnSOD廣泛參與植物的生長發(fā)育與衰老及非生物脅迫等逆境應(yīng)答過程。單粒小麥MnSOD基因的表達在鹽脅迫、氧化脅迫和重金屬脅迫下顯著上調(diào)[10]。紫桿檉柳MnSOD基因的異源表達亦表明,提高MnSOD表達量可以增加植株在鹽脅迫、紫外線脅迫等逆境下的存活率[11]。過表達MnSOD基因的仙客來植株在高溫脅迫下的SOD活性和可溶性蛋白含量升高,而細胞膜透性和丙二醛含量上升速度下降,對高溫脅迫表現(xiàn)出更強的耐受性[12]。Zhou等[13]應(yīng)用病毒誘導(dǎo)基因沉默技術(shù)將文冠果MnSOD沉默后,文冠果受精胚珠發(fā)育停滯,產(chǎn)生成熟果實的百分比降至約5%,說明MnSOD基因與受精胚珠和果實的早期發(fā)育密切相關(guān)。桃果實在采后衰老過程中,隨著氧化脅迫加重,線粒體中MnSOD等多種抗氧化酶類的表達量及酶活性紊亂,導(dǎo)致活性氧積累,加速果實衰老[14]。
獼猴桃果實因富含多種氨基酸、礦質(zhì)元素和維生素而深受消費者喜愛,但采后具有典型的呼吸躍變和生理后熟特點,因而易軟化腐爛,不耐貯藏[15]。溫度顯著影響獼猴桃果實的后熟軟化,因此低溫貯藏是目前延緩獼猴桃軟爛最常用和有效的保鮮手段之一[16-17]。脫落酸(abscisic acid, ABA)和赤霉素(gibberellin acid, GA3)等外源激素也顯著影響獼猴桃的后熟軟化進程[18-19]。多項研究顯示,果蔬的采后衰老軟爛與其細胞內(nèi)活性氧代謝失衡密切相關(guān),清除果蔬細胞內(nèi)過多的活性氧,有利于保持果蔬品質(zhì)和提高果蔬商品性[20]。宦晨[21]對桃果實的活性氧和抗氧化酶研究發(fā)現(xiàn),超氧陰離子含量與SOD基因表達量和酶活性顯著相關(guān)。獼猴桃果實采后酶活性研究表明,其SOD活性隨著貯藏時間的延長呈先升后降的變化規(guī)律[22],但相關(guān)過程的編碼基因及其表達規(guī)律的研究偏少,且其中多為Cu/ZnSOD基因相關(guān)研究[23],鮮有MnSOD基因的報道,限制了從基因與轉(zhuǎn)錄水平研究SOD與獼猴桃果實品質(zhì)的關(guān)系。鑒于此,本研究以米良1號獼猴桃(Actinidiachinensisvar.deliciosaMiliang-1)為試材,在克隆驗證AdMSD1和AdMSD2基因序列的基礎(chǔ)上,對其序列特征、編碼蛋白特性、密碼子偏好性、調(diào)控miRNA、外顯子與內(nèi)含子組成及分子進化情況進行解析,并研究其在各組織器官、果實后熟軟化過程、低溫貯藏過程及脫落酸和赤霉素處理后的表達規(guī)律,以期為后續(xù)研究SOD在獼猴桃果實采后品質(zhì)調(diào)控中的功能奠定基礎(chǔ)。
1 材料與方法
1.1 試驗材料
本試驗使用的植物材料為米良1號獼猴桃,其根、莖、葉片、花、幼果(開花后16 d)和成熟果(開花后160 d)均于2018年采集自福建省農(nóng)業(yè)科學(xué)院果樹研究所獼猴桃種植基地,每樣品隨機從10棵樹上分別采集,重復(fù)3次,經(jīng)液氮速凍,保存于-80℃冰箱,作為基因克隆和組織表達特性分析材料。
參照福建省農(nóng)業(yè)科學(xué)院果樹研究所獼猴桃研究項目組前期的研究方法[24],以果實硬度、大小和可溶性固形物含量等相近的成熟果(開花后160 d)為材料,在25℃進行貯藏,在果實后熟軟化的7個時期取樣,分別為硬熟期(果肉硬度20.43±1.04 kg·cm-2、可溶性固形物含量6.92%±0.42%)、軟化初期(果肉硬度17.72±0.79 kg·cm-2、可溶性固形物含量12.84%±0.31%)、軟化Ⅰ期(果肉硬度13.40±0.76 kg·cm-2、可溶性固形物含量14.12%±0.44%)、軟化Ⅱ期(果肉硬度7.13±0.34 kg·cm-2、可溶性固形物含量15.11%±0.21%)、軟化Ⅲ期(果肉硬度3.84±0.33 kg·cm-2、可溶性固形物含量16.70%±0.34%)、軟化末期(果肉硬度1.11±0.19 kg·cm-2、可溶性固形物含量17.68%±0.46%)和軟爛期(果肉硬度0.42±0.10 kg·cm-2、可溶性固形物含量18.08%±0.38%),每樣品隨機取10個果實,重復(fù)3次。果肉樣品經(jīng)液氮速凍后于-80℃冰箱保存。
以成熟果(開花后160 d)作為試驗材料,進行不同貯藏溫度和激素處理,分別在4℃低溫貯藏1、3、5、7、9和11 d進行取樣,在25℃室溫貯藏1、3、5、7、9和11 d進行取樣,在濃度為50 mg·L-1的ABA溶液中浸潤果實2 min后于25℃貯藏1、3、5、7和9 d進行取樣,在濃度為50 mg·L-1的GA3溶液中浸潤果實2 min后于25℃貯藏1、3、5、7、9和11 d進行取樣。每樣品隨機取10個果實,重復(fù)3次,果肉樣品經(jīng)液氮速凍后于-80℃冰箱保存。
1.2 試驗方法
1.2.1 總RNA的提取與cDNA的合成 按照天根生化科技(北京)有限公司的多糖多酚植物總RNA提取試劑盒說明書提取米良1號獼猴桃不同處理組樣品的總RNA。分別運用1%瓊脂糖凝膠電泳與UV-1800紫外分光光度計(美譜達,上海)檢測RNA的完整性、純度和濃度。再根據(jù)賽默飛世爾中國公司的RevertAidTMFirst Strand cDNA Synthesis Kit試劑盒說明書合成cDNA,作為AdMSDs克隆的模板;根據(jù)北京全式金公司的TransScript? One-Step gDNA Removal and cDNA Synthesis SuperMix試劑盒說明書逆轉(zhuǎn)錄為cDNA,作為AdMSDs定量表達分析的模板。
1.2.2AdMSDs的引物設(shè)計與克隆 以錳超氧化物歧化酶作為檢索詞,在紅陽獼猴桃(Actinidiachinensisvar.chinensisHongyang)基因組[25]中檢索,獲得相關(guān)候選序列,之后應(yīng)用DNAMAN軟件設(shè)計包含完整編碼區(qū)的上下游引物,進行基因的反轉(zhuǎn)錄PCR(reverse transcription PCR, RT-PCR)驗證,引物信息見表1。PCR反應(yīng)液共25 μL:12.5 μL DreamTaqGreen PCR Master Mix(2×)、1 μL上游引物、1 μL下游引物、1 μL模板、9.5 μL無菌雙蒸水。PCR擴增程序:94℃預(yù)變性4 min;94℃變性30 s,55℃(MSD1)/57℃(MSD2)退火30 s,72℃延伸60 s(MSD1)/40 s(MSD2),35次循環(huán);72℃終延伸10 min。PCR擴增產(chǎn)物經(jīng)回收和純化后,委托福州尚亞生物有限公司測序。

表1 基因克隆引物
1.2.3AdMSDs的生物信息學(xué)分析 通過美國國家生物技術(shù)信息中心數(shù)據(jù)庫的BLASTN、BLASTP和Conserved Domains工具分析獼猴桃與其他植物MnSOD的同源性和蛋白結(jié)構(gòu)域。采用在線數(shù)據(jù)庫ExPASy(http://web.expasy.org/protparam/)分析蛋白基本特性、ProtComp v9.0(http://linux1.softberry.com/berry.phtml?topic=protcomppl &group=programs&subgroup=proloc)進行亞細胞定位、MitoPro Ⅱ(https://ihg.gsf.de/ihg/mitoprot.html)查看線粒體運輸肽分布、NetPhos 3.1(https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)預(yù)測磷酸化位點分布、SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和SWISS-MODEL(https://swissmodel.expasy.org/)分析二級結(jié)構(gòu)與三維空間結(jié)構(gòu)。通過EMBOSS(http://www.bioinformatics.nl/cgi-bin/emboss)的chip和cusp工具分析AdMSDs基因編碼區(qū)的有效密碼子數(shù)、GC含量和密碼子第三位的GC含量。應(yīng)用psRNATarget(https://www.zhaolab.org/psRNATarget/)預(yù)測調(diào)控獼猴桃AdMSDs基因的miRNA,期望值設(shè)置為5.0,其他參數(shù)為默認值。采用Clustal X對獼猴桃和其他植物MnSOD蛋白序列進行多重對比,并利用MEGA-X軟件的鄰位相連法(Neighbor Joining)構(gòu)建獼猴桃與其他植物MnSOD的系統(tǒng)進化樹。采用GSDS 2.0(http://gsds.gao-lab.org/)工具分析AdMSDs基因的外顯子與內(nèi)含子組成。
1.2.4AdMSDs的實時熒光定量PCR分析 根據(jù)獲得的AdMSDs序列,在DNAMAN軟件上設(shè)計特異的實時熒光定量PCR(quantitative real-time PCR, qRT-PCR)引物,引物信息見表2,并以表達相對穩(wěn)定的獼猴桃Actin基因(基因編號為Achn107181)作為內(nèi)參基因[26],參照北京全式金公司的TransStart? Top Green qPCR SuperMix試劑盒方法配置反應(yīng)體系進行擴增,Eppendorf定量儀器的程序設(shè)定為:94℃預(yù)變性30 s;94℃變性5 s,59℃/62℃退火15 s,72℃延伸10 s,40次循環(huán);最后在94℃ 15 s,60℃ 15 s,94℃ 15 s條件下熔解。程序運行結(jié)束后,通過 Mastercycler?ep realplex軟件進行標準曲線、熔解曲線和樣品Ct值分析。應(yīng)用2-ΔΔCt公式計算AdMSD1和AdMSD2的相對表達量,通過SPSS軟件的One-way ANOVA法進行AdMSDs表達量差異的顯著性分析,最后在Excel 2016中進行圖表的繪制。

表2 qRT-PCR引物
2 結(jié)果與分析
2.1 AdMSDs的克隆驗證
以米良1號獼猴桃果實的cDNA為模板,分別利用上下游引物MSD1-F/MSD1-R和MSD2-F/MSD2-R進行RT-PCR擴增,得到2條長度分別為803 bp和717 bp的目的條帶(圖1),測序結(jié)果經(jīng)BLASTN分析顯示,2個基因與油柿(Diospyrosoleifera,JQ797740)、茶樹(Camelliasinensis,KR189420)、葡萄(Vitisvinifera,KP271926)、白樺(Betulaplatyphylla,KP711292)和文冠果(Xanthocerassorbifolium,MG322616)等植物的錳超氧化物歧化酶基因具有很高的同源性(>82%),說明獲得的2個基因均為米良1號獼猴桃錳超氧化物歧化酶基因,分別命名為AdMSD1和AdMSD2,GenBank登錄號分別為KY471358和KY471359。編碼框查找結(jié)果表明,AdMSD1包含675 bp的開放閱讀框(open reading frame,ORF),共編碼224個氨基酸;而AdMSD2包含690 bp的ORF,共編碼229個氨基酸。
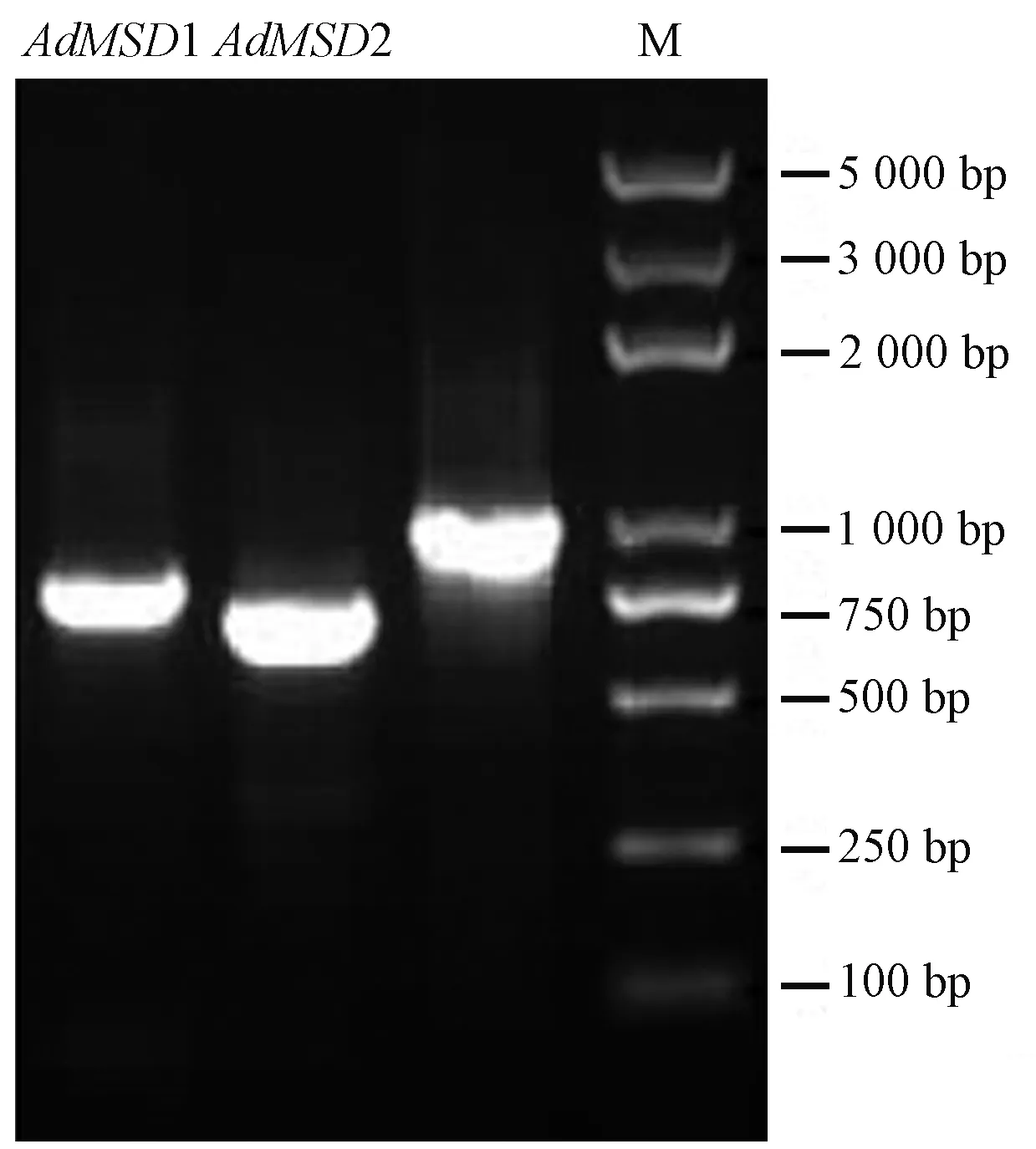
圖1 AdMSDs的擴增結(jié)果
序列兩兩比對分析結(jié)果顯示,AdMSD1與紅陽獼猴桃同源基因Ach24g147991的ORF區(qū)核苷酸相似性為97.67%,氨基酸相似性為96.93%;AdMSD2與紅陽同源基因Ach03g419431的ORF區(qū)的核苷酸相似性為99.13%,氨基酸相似性為100.00%。而AdMSD1與AdMSD2的ORF區(qū)的核苷酸相似性僅為85.65%,氨基酸相似性僅為83.84%。可見,獼猴桃MnSOD的不同成員在序列上存在較大差異,而不同品種間的同源MnSOD具有較高的一致性,說明同源MnSOD基因在獼猴桃種內(nèi)具有高保守性。
2.2 AdMSDs的生物信息學(xué)分析
2.2.1AdMSDs編碼蛋白的特性分析 結(jié)構(gòu)域分析結(jié)果如圖2所示,AdMSD1和AdMSD2均具有植物錳超氧化物歧化酶的3個保守結(jié)構(gòu)域(SodA、Sod-Fe-C和PLN02471)。與其他植物MnSOD氨基酸序列的多重比對結(jié)果亦表明,2個編碼蛋白均包含植物MnSOD的保守金屬結(jié)合域“DVWEHAYY”、Mn2+金屬結(jié)合位點和特征氨基酸(圖3),說明獼猴桃AdMSD1和AdMSD2均具有錳超氧化物歧化酶的基本功能。

圖2 AdMSDs蛋白的保守結(jié)構(gòu)域

注:黑色陰影表示氨基酸序列的完全一致,灰色陰影表示氨基酸序列的一致性較低;Mn2+結(jié)合位點用井號(#)標示,MnSOD特有的5個氨基酸(甘氨酸、甘氨酸、苯丙氨酸、谷氨酰胺和天冬氨酸)用星號(*)標示,保守的金屬結(jié)合域 “DVWEHAYY”用紅色框標示。植物MnSODs的Genbank登錄號:AtMSD(AAC24832),CsMSD1(AJW60327),CsMSD2(AJW60328),VvMSD(AKG51696),MeMSD(XP021618226),HbMSD(XP021687151),XsMSD(AVD97168),CjMSD(ADB10839),TaMSD1(AAB68035),TaMSD2(AAB68036),OsMSD(ATU90093)。
蛋白基本理化性質(zhì)分析結(jié)果表明,AdMSD1和AdMSD2的分子量分別為24.95和25.55 kDa,理論等電點分別為8.81和8.39,總親水性值分別為-0.328和-0.298,不穩(wěn)定系數(shù)分別為39.23和37.20,所帶正電氨基酸數(shù)均為24,所帶負電氨基酸數(shù)分別為21和22,說明其均屬于穩(wěn)定的堿性親水蛋白。亞細胞定位預(yù)測結(jié)果顯示,AdMSD1和AdMSD2均定位在線粒體的可能性最大,分值分別為9.65和9.70;這與MitoProⅡ 分析顯示2種蛋白的N端均具有一段分別為20和24 aa的線粒體運輸肽相符。磷酸化位點分析結(jié)果顯示,AdMSD1擁有17個潛在磷酸化位點,分別為絲氨酸磷酸化位點10個、蘇氨酸磷酸化位點2個、酪氨酸磷酸化位點5個;而AdMSD2擁有22個潛在磷酸化位點,分別為絲氨酸磷酸化位點10個、蘇氨酸磷酸化位點5個、酪氨酸磷酸化位點7個;說明AdMSD1和AdMSD2在翻譯后水平的表達調(diào)控存在差異。AdMSD1蛋白的二級結(jié)構(gòu)由50.89%的α-螺旋、12.95%的延伸鏈、7.14%的β-折疊和29.02%的自由卷曲組成;而AdMSD2的二級結(jié)構(gòu)則由52.84%的α-螺旋、12.23%的延伸鏈、6.55%的β-折疊和28.38%的自由卷曲組成;這與三維空間結(jié)構(gòu)的預(yù)測結(jié)果一致。
2.2.2AdMSDs的密碼子偏好分析 EMBOSS分析結(jié)果顯示,AdMSD1基因ORF區(qū)的有效密碼子數(shù)為59.596、GC含量為49.19%、密碼子第三位的GC含量為53.78%;AdMSD2的有效密碼子數(shù)為61.000、GC含量為48.84%、密碼子第三位的GC含量為52.17%。說明AdMSD1和AdMSD2基因?qū)γ艽a子的選擇偏性較弱,其ORF區(qū)均傾向于選擇A/T堿基,且密碼子偏好以G/C結(jié)尾。
2.2.3 調(diào)控AdMSDs的miRNA預(yù)測分析 應(yīng)用psRNATarget軟件在獼猴桃果實miRNA數(shù)據(jù)庫中分析調(diào)控AdMSD1和AdMSD2的miRNA。結(jié)果顯示AdMSD1的表達受miR172a-5p、miR172b-5p、miR2586a和miR166j的調(diào)控,其中miR172a-5p、miR172b-5p和miR166j通過裂解AdMSD1轉(zhuǎn)錄本的方式調(diào)控其轉(zhuǎn)錄,而miR2586a通過抑制AdMSD1翻譯的方式調(diào)控其表達。AdMSD2的表達則受miR172a、miR172c、miR166j和miR172c-3p的調(diào)控,其中miR172a、miR172c和miR172c-3p通過抑制AdMSD2翻譯的方式調(diào)控其表達,而miR166j通過裂解AdMSD2轉(zhuǎn)錄本的方式調(diào)控其轉(zhuǎn)錄。
2.2.4AdMSDs的外顯子與內(nèi)含子組成分析 基因組的外顯子與內(nèi)含子組成如圖4所示,AdMSD1和AdMSD2均由6個外顯子和5個內(nèi)含子組成,外顯子大小均在47~289 bp,AdMSD1的內(nèi)含子大小在94~1 735 bp,AdMSD2的內(nèi)含子大小在85~726 bp。AdMSD1第1、第2、第3、第4和第6位外顯子的長度與AdMSD2對應(yīng)外顯子的長度相同,分別為289、47、126、57和93 bp。此外,AdMSD1和AdMSD2的5個內(nèi)含子均在GT-AG處發(fā)生剪接,符合真核生物的內(nèi)含子剪接規(guī)律。

注:A: AdMSDs的基因結(jié)構(gòu)。B: AdMSDs的外顯子與內(nèi)含子大小,內(nèi)含子長度用括號標注,長度單位bp。
2.2.5AdMSDs的系統(tǒng)進化樹分析 獼猴桃與其他植物MnSODs的系統(tǒng)進化樹如圖5所示,植物MnSODs共分為兩大分支,分別是雙子葉植物分支(組Ⅰ)和單子葉植物分支(組Ⅱ)。AdMSD1和AdMSD2均聚在雙子葉植物分支(組Ⅰ)中,這與獼猴桃的種屬分類一致。其中,AdMSD1聚在雙子葉植物分支的a組(組I-a),與茶樹MnSOD的親緣關(guān)系近;而AdMSD2聚在雙子葉植物分支的b組(組I-b),與金柑MnSOD的親緣關(guān)系近;說明獼猴桃AdMSD1和AdMSD2屬于植物錳超氧化物歧化酶家族的不同成員,可能起源于2個不同的基因祖先。

圖5 AdMSDs與其他植物錳超氧化物歧化酶的系統(tǒng)進化樹
2.3 AdMSDs在獼猴桃不同組織間的轉(zhuǎn)錄模式
qRT-PCR結(jié)果如圖6所示,AdMSD1和AdMSD2在獼猴桃根、莖、葉、花、幼果和成熟果的轉(zhuǎn)錄水平存在明顯差異。其中,AdMSD1在葉片的轉(zhuǎn)錄水平最高(是根部的46.89倍),其次是莖和幼果,而在根和成熟果中的轉(zhuǎn)錄水平較低,在花中的轉(zhuǎn)錄水平最低(僅為根部的0.14倍);AdMSD2則在花中的轉(zhuǎn)錄水平最高(是根部的9.52倍),而在葉片(是根部的2.51倍)、莖(是根部的1.81倍)、幼果(是根部的1.34倍)、根和成熟果(是根部的0.88倍)中的轉(zhuǎn)錄水平差異不顯著。

注:不同字母表示顯著性差異(P<0.05)。下同。
2.4 AdMSDs在獼猴桃果實后熟軟化過程的表達特征
獼猴桃AdMSDs基因在果實采后從硬熟期至軟爛期的表達模式存在差異(圖7)。AdMSD1基因在獼猴桃果實后熟軟化過程呈“升-降-升-降”的表達模式,即在軟化初期表達顯著上調(diào),進入軟化Ⅰ期和Ⅱ期時,表達量迅速降至硬熟期的0.39和0.35倍,在軟化Ⅲ期時表達量再次升高至硬熟期相當水平,之后繼續(xù)下降,軟爛期時的表達量僅為硬熟期的0.41倍。AdMSD2基因在獼猴桃果實軟化初期的表達量略升高,但不顯著,在軟化Ⅰ期和Ⅱ期時的表達量顯著下降至硬熟期的0.65和0.35倍,進入軟化Ⅲ期后,表達量顯著升高至硬熟期相當水平,之后表達量變化不顯著。

注:S1:硬熟期(對照);S2:軟化初期;S3:軟化Ⅰ期;S4:軟化Ⅱ期;S5:軟化Ⅲ期;S6:軟化末期;S7:軟爛期。
2.5 AdMSDs在果實采后貯藏過程中的表達特性
2.5.1 不同貯藏溫度對果實AdMSDs表達的影響 進一步采用qRT-PCR分析室溫和低溫貯藏過程中AdMSDs基因的變化規(guī)律,結(jié)果如圖8所示,AdMSD1在25℃貯藏1和3 d時表達顯著下調(diào),5 d時開始上調(diào)表達,9 d時的表達量達到最大值,為0 d的4.85倍,11 d時再次下調(diào)表達,僅為0 d的0.33倍;而AdMSD1在4℃低溫貯藏前5 d的表達量均顯著下降(僅為0 d的0.18~0.20倍),7 d時升高至0 d相當水平,之后繼續(xù)下降至0 d的0.35倍以下;而且與對照(25℃貯藏)相比,AdMSD1在4℃貯藏各時間點的表達量均較低。與上述結(jié)果類似,25℃貯藏的獼猴桃果實中AdMSD2的表達量在1和3 d時下降,5 d時顯著上升,7 d時達到最大值(為0 d的6.30倍),隨后下調(diào);4℃低溫貯藏的獼猴桃果實中AdMSD2的表達量在前5 d均顯著下降(為0 d的0.27~0.33倍),7 d時升高至0 d的1.31倍,之后繼續(xù)下降至0 d的0.5倍以下;而且AdMSD2除了在4℃貯藏11 d時的表達量略高于對照(25℃貯藏11 d)外,其余時間點的表達量均比對照低。說明AdMSD1和AdMSD2在果實低溫貯藏時表達下調(diào)。
2.5.2 不同激素處理對果實AdMSDs表達的影響 果實AdMSD1和AdMSD2在脫落酸和赤霉素處理后的表達模式不同(圖8)。ABA處理后獼猴桃果實中AdMSD1表達呈“M”型,即AdMSD1在1 d時的表達量顯著升高,3 d時下降至0 d的0.60倍,5 d時再次顯著升高,之后下降至0 d的0.4倍以下;而ABA處理后獼猴桃果實中AdMSD2的表達持續(xù)下降。GA3處理后,獼猴桃果實中AdMSD1的表達呈下降趨勢;而AdMSD2在處理后1 d時表達量顯著升高,之后隨著貯藏時間的推移雖有波動,但均低于0 d的表達水平。

圖8 低溫貯藏下和ABA、GA3處理后獼猴桃果實中AdMSD1和AdMSD2的表達
3 討論
3.1 獼猴桃MnSOD家族的成員數(shù)量及其序列特征
不同物種擁有錳超氧化物歧化酶基因的數(shù)量存在差異,如黃瓜(Cucumissativus)僅有1個MnSOD基因(CsMSD)[27]、小麥(Triticumaestivum)有3個MnSOD基因(TaSOD2.1、TaSOD2.2、TaSOD2.3)[28]、香蕉(Musaacuminata)有4個MnSOD基因(MaMSD1A、MaMSD1B、MaMSD1C、MaMSD1D)[5]、芥菜(Brassicajuncea)有7個MnSOD基因(BjuAMSD1a、BjuAMSD1b、BjuAMSD1c、BjuAMSD1d、BjuBMSD1e、BjuBMSD1f、BjuBMSD1g)[29]。本研究從獼猴桃中共獲得2個MnSOD基因(AdMSD1和AdMSD2)。多項研究表明,不同植物MnSOD基因數(shù)量的差異與其基因組經(jīng)歷的多倍化、全基因組重復(fù)及基因片段重復(fù)等事件相關(guān)[28-29],獼猴桃的MnSOD數(shù)量可能與其在進化過程經(jīng)歷的1次六倍化和2次全基因組重復(fù)事件相關(guān)[25]。與小麥MnSOD蛋白具有線粒體靶向序列相似[30],AdMSD1和AdMSD2蛋白的N端均含有一段20~24 aa的線粒體運輸肽,這與其亞細胞定位結(jié)果相符;而且,與已報道的植物錳超氧化物歧化酶相同,AdMSD1和AdMSD2具有保守金屬結(jié)合域“DVWEHAYY”、Mn2+金屬結(jié)合位點和5個特征氨基酸,由此推測AdMSD1和AdMSD2蛋白主要在線粒體中行使活性氧清除功能。此外,AdMSD1和AdMSD2與擬南芥[5]、龍眼[7]、茶樹[31]等多數(shù)植物的錳超氧化物歧化酶基因相同,均由6個外顯子和5個內(nèi)含子組成,推測植物MnSOD的基因結(jié)構(gòu)具有較高保守性。但在系統(tǒng)進化分析中,AdMSD1和AdMSD2蛋白分別聚在雙子葉植物組的不同分支,且其核苷酸和氨基酸序列存在差異,說明2種蛋白屬于植物錳超氧化物歧化酶家族的不同成員,可能源自不同的基因祖先。
3.2 AdMSD1和AdMSD2在不同組織器官的表達差異
諸多研究表明,MnSOD基因在植物各組織器官廣泛轉(zhuǎn)錄表達。茄子(Solamummelongena)SmMSD在根、莖、葉、花瓣、果皮和果肉中均有表達,但以葉片中的表達量最高,果肉中的表達量最低[32]。黃瓜CsMSD在葉片、花和莖中少量表達,而在根和果實中未檢測到表達[27]。在香蕉中,4個MnSOD基因在根、假莖和葉中均有表達;其中MaMSD1A、MaMSD1C和MaMSD1D的表達模式相似,均在根部大量表達,其次是葉片,在假莖中的表達量最低;MaMSD1B則在根部的表達量最高,假莖中次之,葉中最低[5]。植物MnSOD各成員在不同組織器官的差異表達可能與其功能多樣性和互補性有關(guān)。獼猴桃AdMSD1和AdMSD2與茄子SmMSD類似,在所有被檢測的組織器官(根、莖、葉片、花、幼果和成熟果)均有轉(zhuǎn)錄,但AdMSD1在葉片的轉(zhuǎn)錄水平最高,在花中的轉(zhuǎn)錄水平最低,而AdMSD2在花中的轉(zhuǎn)錄水平最高,在成熟果中的轉(zhuǎn)錄水平最低。結(jié)合2個蛋白的分子進化不同、磷酸化位點的數(shù)量與分布差異、二級結(jié)構(gòu)和三級結(jié)構(gòu)的不同,推測AdMSD1和AdMSD2雖然參與清除獼猴桃各組織間的活性氧,但具體分工有差異,仍待后續(xù)進一步驗證。
3.3 AdMSD1和AdMSD2參與獼猴桃果實后熟軟化和采后貯藏品質(zhì)的調(diào)控
植物MnSOD基因的表達受溫度調(diào)控。非洲菊(Gerberajamesonni)GjMnSOD基因在4℃低溫處理后的表達受到極顯著抑制,且在各處理時間點的表達量均低于對照[9]。低溫脅迫后,茄子SmMSD基因的轉(zhuǎn)錄顯著下調(diào),處理3 h的表達量降為對照的20%,之后一直保持該水平[32]。本研究中,果實AdMSD1和AdMSD2在低溫貯藏的整個過程一直保持較低的轉(zhuǎn)錄水平,而在室溫貯藏的5~9 d就顯著上調(diào)并出現(xiàn)轉(zhuǎn)錄高峰。這與桃(Prunnuspersia)果實MnSODs基因(PpaSOD3和PpaSOD4)在低溫貯藏下始終保持低表達水平[21]相似,也與獼猴桃Cu/ZnSODs基因(AdCSD1和AdCSD2)在低溫和室溫貯藏中的轉(zhuǎn)錄模式[23]類似。陳金印[35]對獼猴桃在低溫和室溫貯藏過程的SOD酶活性研究亦表明,低溫貯藏過程中果實的SOD酶活性一直保持在較低水平,僅在貯藏后25 d出現(xiàn)小高峰,之后呈下降趨勢;而室溫貯藏過程中SOD酶活性上升快速,貯藏后10 d出現(xiàn)高峰,且峰值是低溫貯藏的2倍。可見,不同溫度貯藏下獼猴桃果實的SOD在轉(zhuǎn)錄水平和蛋白水平呈現(xiàn)相似的變化規(guī)律。前人研究表明,果蔬中活性氧的積累是導(dǎo)致其衰老和品質(zhì)下降的主要原因之一[20]。宦晨[21]對桃果實的研究發(fā)現(xiàn),超氧陰離子(活性氧種類之一)含量由SOD在蛋白和轉(zhuǎn)錄水平共同調(diào)控。而SOD催化超氧陰離子發(fā)生歧化反應(yīng),生成的產(chǎn)物H2O2是重要的信號因子。H2O2可以調(diào)控葡萄果實中細胞壁去乙酰化基因(GDSL)、細胞壁降解基因(XTH)等表達,影響果實的成熟[36];H2O2也參與番茄果實成熟相關(guān)基因ACO5、ACS6和多聚半乳糖醛酸酶基因PG的表達調(diào)控,影響果實的成熟軟化[37]。與室溫貯藏相比,獼猴桃果實在低溫貯藏下的硬度下降緩慢,乙烯合成酶關(guān)鍵基因(ACO1和ACS1)和淀粉酶基因(BAM1和BAM3)等的表達受到抑制,果實軟化減緩、耐貯性提高[16,38]。因而,獼猴桃果實在采后可能通過SOD(MnSODs和Cu/ZnSODs)等相關(guān)基因轉(zhuǎn)錄水平的改變,從而調(diào)控SOD等酶活性,參與果實后熟軟化進程的活性氧清除,進而影響其采后軟化速度與貯藏品質(zhì)。
外源ABA和GA3調(diào)控植物MnSOD基因的表達。丹參(Salviamiltiorrhiza)SmMSD1和SmMSD3在ABA處理后表達下調(diào),SmMSD2在ABA處理6 h時表達顯著上調(diào)[6]。茶樹(Camelliasinensis)CsMSD1基因在GA3處理后下調(diào)表達,在24 h時達最低值,而在48 h時顯著上調(diào)表達[31]。相關(guān)研究表明,不同MnSOD基因成員在外源ABA和GA3處理后的表達差異可能與其啟動子區(qū)的脫落酸響應(yīng)元件ABRE、赤霉素響應(yīng)元件GARE的差異有關(guān)[5-6]。ABA處理5 d顯著誘導(dǎo)獼猴桃果實AdMSD1表達,抑制AdMSD2表達;而GA3處理抑制AdMSD1表達,在處理1 d時誘導(dǎo)AdMSD2表達。AdMSD1和AdMSD2對外源ABA和GA3的不同響應(yīng),是否與其啟動子區(qū)所含響應(yīng)元件的差異有關(guān)仍待試驗驗證。
4 結(jié)論
本研究克隆獲得2個獼猴桃錳超氧化物歧化酶基因(AdMSD1和AdMSD2),2個基因編碼的蛋白均具有植物MnSOD家族的基本特征,但聚類在進化樹的不同分支,屬于MnSOD家族的不同成員。AdMSD1在葉中的表達量最高,而AdMSD2在花中的表達量最高。AdMSD1和AdMSD2均參與獼猴桃果實后熟軟化過程,且在低溫貯藏的轉(zhuǎn)錄下調(diào),2個基因在果實中的表達均受ABA和GA3的調(diào)控。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
