瞬時過表達MnERF2基因對桑樹耐鹽性的影響
2022-05-14 02:52:28董亞茹聶玉霞李云芝趙東曉耿兵王照紅
山東農業科學 2022年4期
關鍵詞:植物
董亞茹,聶玉霞,李云芝,趙東曉,耿兵,王照紅
(山東省蠶業研究所,山東 煙臺 264002)
土壤鹽漬化是自然界常見的逆境,全世界約有20%的耕地受到鹽漬化威脅,嚴重影響了植物的生長、光合作用及生物量積累、產量等[1,2]。植物對非生物脅迫的反應受到多種因素的影響,涉及大量抗性基因的表達與調控,因此要提高植物對逆境的抗性就要從一些關鍵的調節轉錄因子著手。AP2/ERF類轉錄因子是植物特有的與脅迫應答有關的轉錄因子超家族。該類轉錄因子都具有1個或2個由60~70個氨基酸殘基組成的非常保守的AP2/ERF結構域。Sakuma等[3]依據AP2/ERF轉錄因子的AP2結構域相似性將其劃分為5大類,即AP2、ERF、DREB、RAV和Soloist。不同植物中AP2/ERF轉錄因子及其各亞家族成員的數量存在差異。桑樹中共鑒定到116個AP2/ERF轉錄因子家族成員,其中ERF 58個,AP233個,DREB 21個,RAV 3個,Soloist 1個[4]。
20世紀90年代,第一個AP2/ERF轉錄因子從擬南芥中分離出來,研究表明其參與調控了花的發育過程[5]。事實證明,AP2/ERF轉錄因子不僅參與調控植物的生長發育,在響應低溫、干旱、高鹽、缺氧、高溫等逆境脅迫方面也起到非常重要的作用[6,7]。擬南芥中AtCBF3基因可以誘導編碼脯氨酸合成關鍵酶的表達,增加體內游離脯氨酸含量,提高植株的耐寒性[8]。過表達AtERF1基因的轉基因擬南芥植株同時表現出對高溫、高鹽及干旱的耐受性,形態學觀察發現植株通過減小葉片氣孔孔徑以降低水分流失[9]。覃利萍[10]在對剛毛檉柳的研究中發現ThCRF1轉錄因子可以促進海藻糖和脯氨酸的生物合成以及提高SOD和POD活性,從而提高植物對鹽脅迫的耐受性。Yao等[11]發現擬南芥ERF74和ERF75受缺氧脅迫誘導,通過調控缺氧響應基因的表達來提高植株的缺氧耐受能力,同時敲除ERF74和ERF75的突變體植株表現出對淹水脅迫較高的敏感性。高溫脅迫下擬南芥AtCBF3顯著表達,與野生型相比,轉AtCBF3基因的馬鈴薯植株ROS積累量減少,光合作用顯著提高,表現出更強的耐高溫性[12]。此外,AP2/ERF類轉錄因子也參與乙烯的生物合成及信號傳導。
本研究將MnERF2基因過表達載體(pROKⅡ-MnERF2)、抑制表達載體(pROKⅡ-MnERF2-SRDX)瞬時轉入桑樹中并通過NaCl脅迫使其表達,比較分析了不同轉基因植株的耐鹽生理指標,初步鑒定MnERF2的功能,以期為桑樹及木本植物耐鹽分子機制研究奠定理論基礎。
1 材料與方法
1.1 試驗材料
供試桑樹品種為雜交種桂桑優12,種子購于廣西蠶業技術推廣總站。試驗于2021年3月在山東省蠶業研究所組培實驗室進行。挑選飽滿一致的種子,在超凈臺中用75%乙醇消毒1 min,然后用5%次氯酸鈉消毒10 min,最后用無菌水沖洗5次,鋪種到裝有MS培養基的培養皿中。一周后挑選萌發無菌的種子移栽到裝有MS培養基的組培瓶中,每瓶4顆種子,以此獲得大量無菌桑樹植株,用于瞬時侵染試驗。植物過表達載體(pROKⅡ-MnERF2)、抑制表達載體(pROKⅡ-MnERF2-SRDX)由本實驗室構建保存。MnERF2屬于ERF亞家族,GenBank登錄號為XM_010106513,由本實驗室從桂桑優12中克隆獲得。
1.2 MnERF2基因過表達和抑制表達載體瞬時轉化桑樹
選擇生長一致的1個月左右的桑樹無菌苗,將構建好的MnERF2基因過表達載體(pROKⅡ-MnERF2,OE)、抑 制 表 達 載 體(pROKⅡ-MnERF2-SRDX,RNAi)和空載體(pROKⅡ,WT)分別瞬時侵染桑樹組培苗,侵染方法參照Ji等[13]的描述。桑樹植株在1/2MS培養基中培養48 h后移至含有100 mmol·L-1NaCl的1/2MS培養基中,在脅迫處理0、6、12、24、48 h取整株桑樹材料,經液氮速凍后保存于-80℃冰箱,用于后續試驗。
1.3 MnERF2基因瞬時表達桑樹的抗逆生理指標測定
1.4 實時熒光定量PCR
用植物RNA試劑盒提取桑樹植株總RNA,并反轉錄成cDNA。將反轉錄產物稀釋10倍,選擇MnRPL15和β-actin作為內參基因,引物見表1,由上海生工生物有限公司合成。RT-PCR反應體系為:TB Green PremixEx Taq10μL,基因特異性上下游引物各1μL(10μmol·L-1),模板2 μL,補ddH2O至20μL。每個測試樣品進行3次重復試驗。反應程序按照TB Green PremixEx Taq(TaKaRa No.RR820Q)說明進行,在CFX96 Real-Time PCR Detection System儀器(Bio-Rad Bio.CA.USA)上完成。RT-PCR數據利用2-△△Ct法[21]進行分析。

表1 RT-PCR引物及其序列
2 結果與分析
2.1 MnERF2基因瞬時表達桑樹植株的獲得
NaCl脅迫48 h后對MnERF2基因瞬時表達量進行RT-PCR分析,結果(圖1)顯示,MnERF2基因在瞬時過表達(OE)株系中的表達量最高,在抑制表達(RNAi)株系中最低。表明已成功獲得了瞬時過表達和抑制表達MnERF2基因的桑樹植株。

圖1 MnERF2基因瞬時轉化桑樹植株qRT-PCR檢測
2.2 MnERF2基因瞬時表達桑樹植株的抗性生理研究
2.2.1 ROS及MDA含量的變化 NaCl脅迫后,MnERF2基因OE株系中的、H2O2、·OH和MDA含量均低于野生株,而RNAi株系中的含量高于野生株,且隨著處理時間的延長,各株系中的、H2O2、·OH和MDA含量均不斷增加。脅迫48 h時,OE株系的、H2O2、·OH和MDA含量分別比野生株降低了17.67%、34.42%、30.24%和27.40%,而RNAi株系分別比野生株升高了27.45%、46.17%、21.74%和9.71%(圖2)。
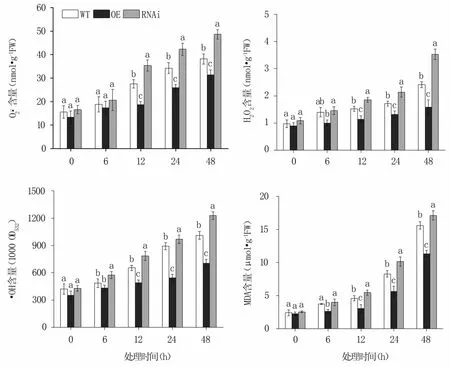
圖2 NaCl脅迫不同時間轉基因和野生桑樹中ROS和MDA含量的變化
2.2.2 抗氧化酶活性 NaCl脅迫后,MnERF2基因OE株系中SOD、POD、CAT、GST活性均強于對照,RNAi株系中則均弱于對照,且隨著處理時間的延長各株系中的四種保護酶活性均不斷增加。脅迫48 h時,OE株系的SOD、POD、CAT、GST活性分別比野生株提高了36.05%、23.86%、37.98%、24.46%,RNAi株系的則比野生株降低了28.16%、29.32%、23.74%、22.24%(圖3)。
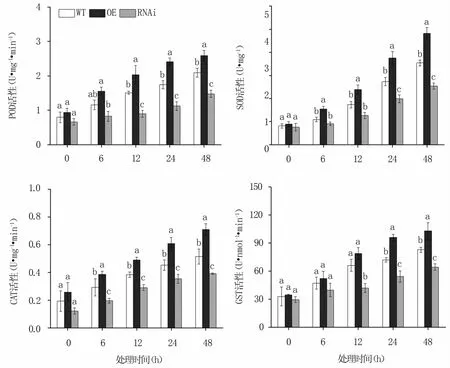
圖3 NaCl脅迫不同時間轉基因和野生桑樹中SOD、POD、CAT和GST活性的變化
2.2.3 滲透調節物質含量 NaCl脅迫后,MnERF2基因OE株系中AsA、GSH、脯氨酸含量均高于對照,而RNAi株系中均低于對照,隨著處理時間的延長均不斷增加。脅迫48 h時,OE株系的AsA、GSH、脯氨酸含量分別比野生株提高41.73%、22.47%、34.47%,RNAi株系則分別比野生株降低25.18%、18.32%、31.03%(圖4)。
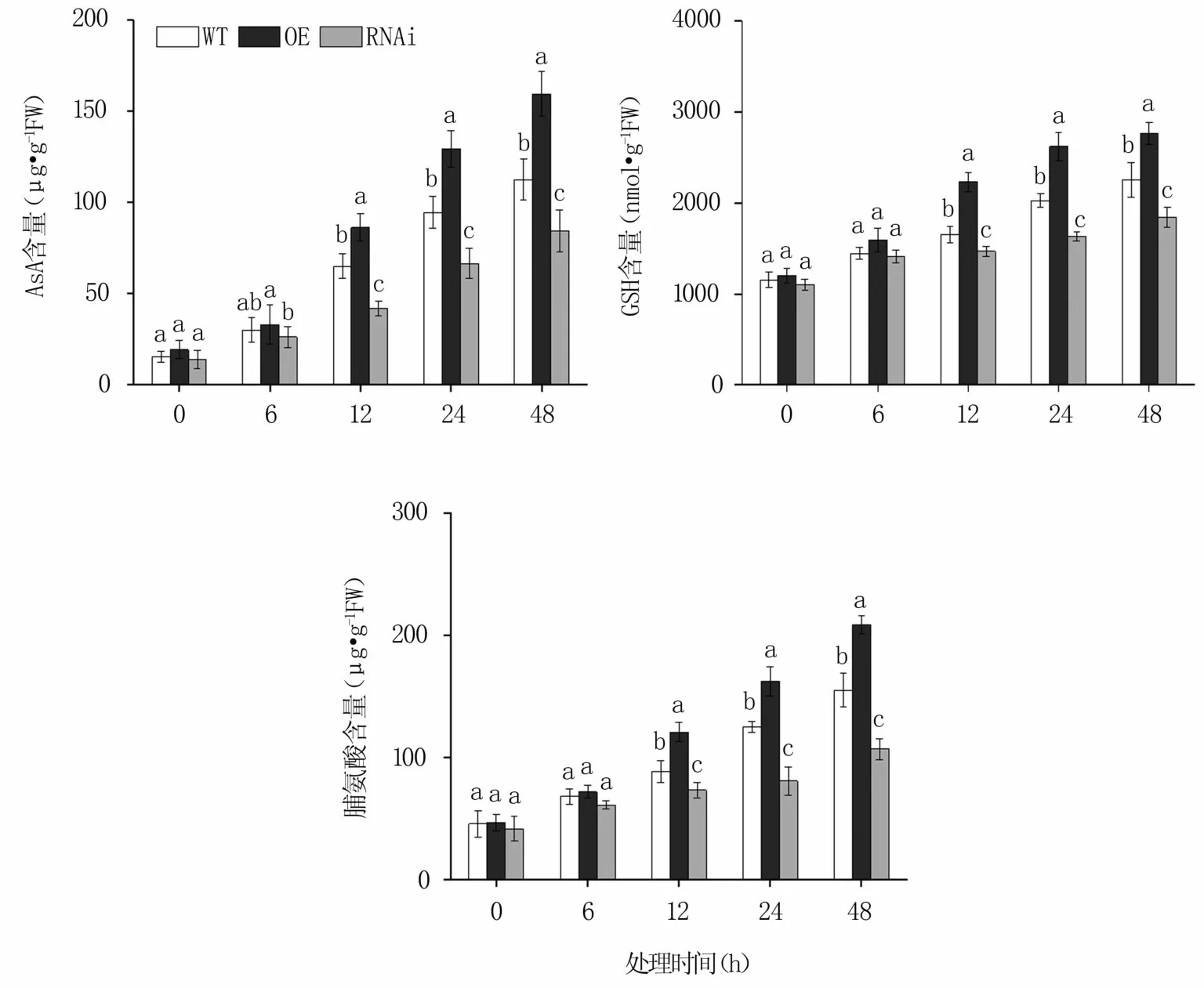
圖4 NaCl脅迫不同時間轉基因桑樹和野生桑樹中AsA、GSH和脯氨酸含量的變化
2.3 抗氧化酶基因表達分析
由于脅迫48 h的抗氧化酶活性最高,因此對該時間SOD、POD、CAT三種酶基因的表達量進行分析,結果(圖5)顯示,MnERF2基因的OE株系中三種酶基因的表達量均顯著高于對照,而RNAi株系中三種酶基因的表達量均顯著低于對照。
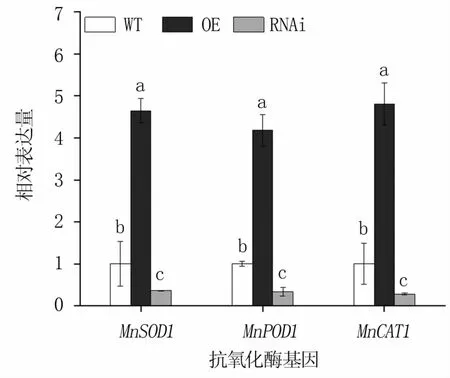
圖5 NaCl脅迫48 h轉基因和野生桑樹中抗氧化酶基因的表達量
3 討論與結論
轉錄因子可以與啟動子內的順式元件相互作用,調節下游基因的轉錄,在植物抵抗非生物脅迫中具有重要作用[22]。因此,發掘抗非生物脅迫相關轉錄因子,研究其抗逆分子機制,能夠為植物抗逆分子改良提供重要數據和材料。已有研究顯示,不同植物的ERF轉錄因子通過調控脅迫響應基因,在高鹽度、干旱和低溫等非生物脅迫響應中發揮重要作用[23-26]。本研究前期在桑樹鹽脅迫轉錄組數據分析中發現ERF基因響應鹽脅迫的誘導,并從中篩選到一個上調表達倍數較高的ERF基因,命名為MnERF2,推測其在桑樹耐鹽響應中發揮一定功能。
為了驗證MnERF2基因的耐鹽功能,分析桑樹響應鹽脅迫的生理生化變化,本研究分別構建植物過表達載體(pROKⅡ-MnERF2)及抑制表達載體(pROKⅡ-MnERF2-SRDX),通過農桿菌介導的高效瞬時遺傳轉化獲得了MnERF2基因瞬時過表達(OE)、抑制表達(RNAi)桑樹植株,RT-PCR檢測結果顯示,MnERF2基因在OE植株中的表達量顯著高于在RNAi植株中,表明獲得了瞬時表達桑樹植株,這與在擬南芥[21]、白樺[27]、檉柳[28]上的研究結果類似,可用于后續抗逆功能的研究。
活性氧(ROS)是有氧代謝的副產物[29]。當植物遭遇鹽和干旱脅迫時,ROS會過度積累,導致DNA、脂類、蛋白質的損傷,最終導致氧化應激[30]。通過抗氧化系統清除ROS,可將ROS保持在較低的穩態水平[31]。鹽脅迫可以顯著影響植物細胞質膜脂質過氧化,從而改變質膜滲透性,調節離子的滲透模式,鹽脅迫引起的這種膜損傷也與ROS的增加有關[32]。MDA是脂質過氧化作用的最終產物,其積累量反映了氧自由基引起的細胞膜損傷程度[13]。為此,本研究分別對NaCl脅迫條件下MnERF2基因過表達、抑制表達及野生桑樹植株的ROS和MDA含量進行比較分析,結果顯示,隨著脅迫時間的延長,過表達MnERF2基因可以降低桑樹在鹽脅迫下的O2·-、H2O2、·OH和MDA含量,說明MnERF2基因可以降低鹽脅迫下的氧化應激,從而保護細胞膜、減輕損傷,具有一定的耐鹽性。
為了避免ROS過度積累造成的氧化損傷,植物自身抗氧化系統會通過提高抗氧化酶活性和非酶物質來清除ROS。谷胱甘肽S-轉移酶(GST)是一種多功能酶,在外源性物質的解毒和應激代謝中起著至關重要的作用,是植物在逆境條件下的重要因子,可防止氧化損傷[33]。擬南芥AtGSTU19在鹽度和干旱條件下表達上調,檉柳ThGSTZ1基因在干旱和鹽脅迫下表達上調[34,35]。本研究中,過表達MnERF2基因可以顯著提高桑樹在鹽脅迫下的SOD、POD、CAT、GST活性,具有正向調控作用,從而增強ROS清除能力,進而提高桑樹的耐鹽性。這與前人[28,36]的研究結果類似。
抗壞血酸(AsA)具有抗氧化作用,能有效清除羥基自由基和過氧化物[37]。谷胱甘肽(GSH)是一種低分子量的硫醇,廣泛分布在動植物中[38,39],可以直接去除活性氧,是一種重要的抗氧化劑[40]。同時,GSH作為GST的底物,具有活性氧清除作用[38]。游離脯氨酸(Pro)可以調節細胞膜的滲透性,在穩定膜和蛋白質等生物大分子結構方面發揮作用,起到對鹽、干旱和污染物脅迫的抗性作用[41]。本研究中,過表達MnERF2基因可以使鹽脅迫桑樹中的AsA、GSH、脯氨酸含量顯著高于對照和抑制表達植株。由此可見,MnERF2能夠通過控制鹽脅迫下AsA、GSH、脯氨酸的生物合成來增加植物的抗氧化物質含量,提高滲透勢,最終提高植物的耐鹽性。
綜上所述,鹽脅迫下過表達MnERF2基因可通過增強抗氧化酶SOD、POD、CAT、GST活性,增加抗氧化物質和滲透調節物質含量,增強ROS清除能力,減少細胞受損或死亡,從而提高桑樹的耐鹽能力。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
