蘋果梨果皮的7種酚類物質對鏈格孢的離體抑制作用
2022-05-14 08:24:28張婷婷王調蘭李永才
食品工業科技 2022年10期
關鍵詞:生長
張婷婷,王調蘭,李永才
(甘肅農業大學食品科學與工程學院,甘肅蘭州 730070)
鏈格孢(Alternaria alternata)是一種可以引起多數水果發生采后病害的真菌病原物,可對棗[1]、蘋果[2]、柑橘[3]、藍莓[4]以及蘋果梨[5]等造成嚴重的病害。由該病原菌引起的黑斑病是蘋果梨主要的采后病害。黑斑病不僅造成蘋果梨的品質劣變,而且在果實體內產生的鏈格孢毒素對人體健康造成嚴重威脅,制約著蘋果梨產業的健康發展。目前,合成殺真菌劑被廣泛應用于控制果實采后病害的發生[6],其效果雖然較好,但由于廣泛應用導致了病原菌耐藥性顯著增加,且對人類及環境造成危害[7]。因此研究開發控制果蔬采后病害的天然防腐劑已勢在必行。
酚類物質作為植物天然次級代謝產物具有抗氧化、清除自由基、抵御生物和非生物脅迫作用。已有研究表明酚類物質可通過限制病原體生長、增加植物抗性的化學屏障等抵御病原物的侵染[8]。近期研究發現蘋果、柑橘、山楂和櫻桃中提取的酚類物質可以有效清除果實體內的自由基[9]。同時果皮中的酚類物質對病原菌的侵染也具有一定的影響,如在未成熟蘋果中的綠原酸,桃果皮和晚熟甜櫻桃中的酚酸能有效抑制盤長孢(Gloeosporium perennans),桃褐腐病菌(Monilinia fructicola)和A. alternata侵入寄主[10?11]。進一步研究發現,果實中的酚類物質還可以通過損傷病原物的細胞膜進而延緩其致病性,如對香豆酸甲酯(乙酯)和水楊酸可以對A. alternata[12?13]和擴展青霉(Penicillium expansum)[14]的質膜造成損傷從而促進內溶物的泄露,降低其對果實的致病性。另外酚類物質影響A. alternata色素和毒素的產生[10]。但由于果皮酚類物質種類、含量及存在狀態具有較大差異,因此其調控機理也呈現復雜性[15]。雖然已有部分研究報道了酚類物質對病原真菌的調控作用,但蘋果梨果皮中的酚類物質對A. alternata生長以及致病性的調控機制鮮有報道,因此在我們前期發現的蘋果梨果皮中主要7種酚類物質的基礎上[16],本文進一步研究評價了蘋果梨果皮的7種酚類物質對A. alternata的生長以及黑色素產生的影響,以期為揭示酚類物質調控該病原物的致病機理提供有效的理論依據。
1 材料與方法
1.1 材料與儀器
鏈格孢(Alternaria alternata) 分離于貯藏中自然發病的蘋果梨果實,純化后PDA培養基保存待用;酚類物質標準品(綠原酸、咖啡酸、阿魏酸、對香豆酸、兒茶素、表兒茶素、熊果苷) 上海源葉生物科技有限公司;黑色素標準品 Sigma公司;瓊脂、Tween-80 柯意哲(上海)機電工程有限公司;75%酒精 山東利爾康醫療科技股份有限公司。
1.2 實驗方法
1.2.1 病原物分離、純化與孢子懸浮液的配制 將有黑斑病的蘋果梨果實,以75%酒精進行果實表面消毒,再用無菌水沖洗后切取病健交界處的組織,在無菌操作條件下移至馬鈴薯葡萄糖(PDA)培養基上,然后于28 ℃恒溫培養,待其長出分生孢子之后再進行分離、純化,并通過回接試驗確定其致病性。分離純化的病原物在馬鈴薯葡萄糖(PDA)培養基上保存備用[16]。
取PDA上培養5 d的A. alternata菌種,加入含有0.05%的Tween-80的適量無菌水,用無菌涂布器輕輕刮涂平板上的病原菌孢子,然后用4層已滅菌的紗布進行過濾,將過濾后的孢子懸浮液轉移到無菌三角瓶中,并立即封口。隨后將配制好的孢子懸浮液在漩渦振蕩器上振蕩20 s使孢子分散均勻,并用血球計數板在光學顯微鏡下進行計數,添加無菌水進行稀釋,使其達到實驗所需不同濃度。最后將配制好的A. alternata孢子懸浮液在6000 ×g條件下離心10 min,并棄去上清液,沉淀用于后續試驗。
1.2.2 蘋果梨果皮的酚類物質處理對A. alternata生長的影響
1.2.2.1 外源酚類物質對A. alternata孢子萌發的影響 無菌條件下,用打孔器取直徑為9 mm的2%的水瓊脂培養基,然后將其置于已滅菌的載玻片上,在1.2.1中得到的孢子懸浮液(1×106個/mL)的沉淀中分別加入等量的綠原酸(1、2、4 mg/mL)、咖啡酸(0.2、0.4、0.6 mg/mL)、阿魏酸(0.5、1、2 mg/mL )、對香豆酸(0.3、0.6、1.2 mg/mL)、兒茶素(0.5、1、2 mg/mL)、表兒茶素(0.2、0.4、0.8 mg/mL)以及熊果苷(7.5、15、30 mg/mL)溶液,混合均勻后,分別在水瓊脂培養基上用移液槍滴加20 μL的混合溶液,此濃度來源于前期實驗研究得到的基于蘋果梨果皮的7種酚類物質含量[16]。置于28 ℃下培養,分別在萌發2、4、6、8 h時鏡檢觀察A. alternata萌發率,直至對照完全萌發。以無菌水處理作為對照。每次鏡檢100個孢子,重復3次。
1.2.2.2 外源酚類物質對A. alternata菌絲生長的影響 分別將不同濃度的上述酚類物質溶液加入無菌的PDA培養基中,混合均勻,倒入無菌的培養皿中,分別接種28 ℃培養7 d的直徑為8 mm的A.alternata菌餅,然后于28 ℃條件下恒溫培養5 d,每天測量菌落直徑。以無菌水作為對照。每個處理3個平行,重復3次。
1.2.2.3 外源酚類物質對A. alternata菌絲生物量的影響 上述酚類物質溶液加入無菌的PDA培養基中,混合均勻,倒入無菌的培養皿中,待凝固后在培養基表面鋪上無菌的玻璃紙,分別接種28 ℃培養7 d的直徑為8 mm的A. alternata菌餅,然后于28 ℃條件下恒溫培養5 d后取下玻璃紙,稱重。以無菌水作為對照。每個處理3個平行,重復3次。
1.2.3 外源酚類物質對A. alternata黑色素的影響
1.2.3.1 黑色素標準曲線的制作 分別配制濃度為0、10、20、30、40、50 mg/L的黑色素標準溶液,在400 nm處測定吸光值,制作標準曲線。
伊匹單抗是一種抑制CTLA-4的完全人源化IgG1單克隆抗體[47],FDA 最終于2011年批準伊匹單抗用于治療轉移性黑色素瘤。伊匹單抗治療過程中出現的孤立性心臟不良反應已見報道,除了發生率<1%的心肌炎,心包積液和罕見心臟不良反應(心律失常,例如心房顫動等)也可能發生[48]。1例使用伊匹單抗治療的患者出現了心肌纖維化伴肝炎[1],另1例由伊匹單抗導致的心臟驟停也被報道[49]。1例接受4個標準劑量伊匹單抗治療的83歲女性轉移性黑色素瘤患者被診斷出患Takotsubo綜合征,這與CTLA-4失活所致的自身免疫性心肌炎有關[50]。
1.2.3.2 黑色素的提取 按照1.2.2.3中的方法,將培養5 d的A. alternata菌絲取下后,稱取0.25 g置于研缽中,加入液氮,研磨,置于30 mL 1 mol/L NaOH的錐形瓶中,進行沸水浴5 h提取,每30 min左右晃蕩一次,注意補充NaOH。?20 ℃冷卻20 min,雙層濾紙過濾,濾液用7 mol/L HCl調節pH2.0,10000 ×g離心15 min,得到沉淀為黑色素粗提物。
1.2.3.3 黑色素的純化與測定 向1.2.3.2中得到的黑色素粗提物加入5 mL 7 mol/L HCl,充分混勻,沸水浴2 h,10000 ×g離心15 min,沉淀用1 mol/L NaOH溶解,再用7 mol/L HCl調pH至2.0,10000 ×g離心15 min,重復3次。沉淀用1 mol/L NaOH溶液定容。以1 mol/L NaOH為空白,用紫外分光光度計于400 nm處測定溶液的吸光值。A. alternata菌絲中黑色素含量(y,mg/g),x為吸光值,計算公式為:
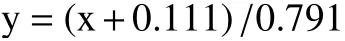
1.3 數據處理
所有試驗數據采用Microsoft Excel 2019計算標準偏差,Origin 2021作圖,用SPSS 23.0軟件對得到的數據進行方差分析,采用Duncan’s多重差異顯著分析。圖中豎線代表標準誤,相同字母表示差異不顯著(P<0.05)。
2 結果與分析
2.1 蘋果梨果皮酚類物質對A. alternata孢子萌發的影響
2.1.1 酚酸類物質處理對A. alternata孢子萌發的影響A. alternata孢子萌發率隨培養時間的延長而增加,不同濃度的酚酸類物質處理后A. alternata萌發率均顯著低于對照(P<0.05),且存在濃度依賴性(圖1),對照組和處理組的孢子萌發率隨著培養時間的增加逐漸增加,且對照組與處理組之間存在顯著性差異(P<0.05)。在培養8 h后,四個酚酸類物質對A.alternata的孢子萌發均有一定的抑制作用(圖1),其中2 mg/mL阿魏酸、1.2 mg/mL對香豆酸、4 mg/mL綠原酸和0.6 mg/mL咖啡酸處理組的孢子萌發率分別較對照降低了98.5%、86.0%,41.5%和6.5%。
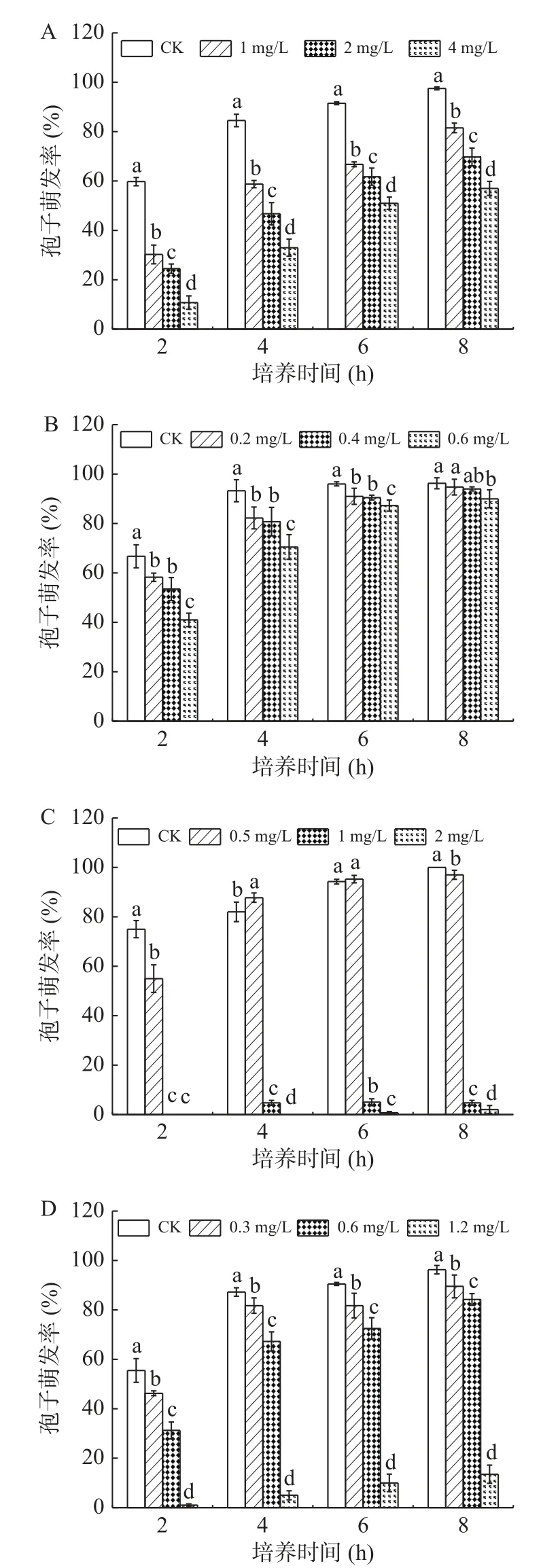
圖1 酚酸類物質處理對A. alternata孢子萌發的影響Fig.1 Effect of phenolic acids treatment on spore germination of A. alternate
2.1.2 黃酮類物質處理對A. alternata孢子萌發的影響 由圖2可知,在整個培養時間內,孢子萌發率隨著培養時間的延長逐漸增加,不同濃度的表兒茶素和兒茶素處理對A. alternata孢子萌發率均有一定的抑制作用,且濃度越大抑制作用越強。在培養2~6 h時,抑制作用明顯,培養8 h時,抑制作用減弱,0.8 mg/mL表兒茶素和2 mg/mL兒茶素處理組與對照之間存在顯著性差異(P<0.05),孢子萌發率分為對照的11.9%和6.7%。
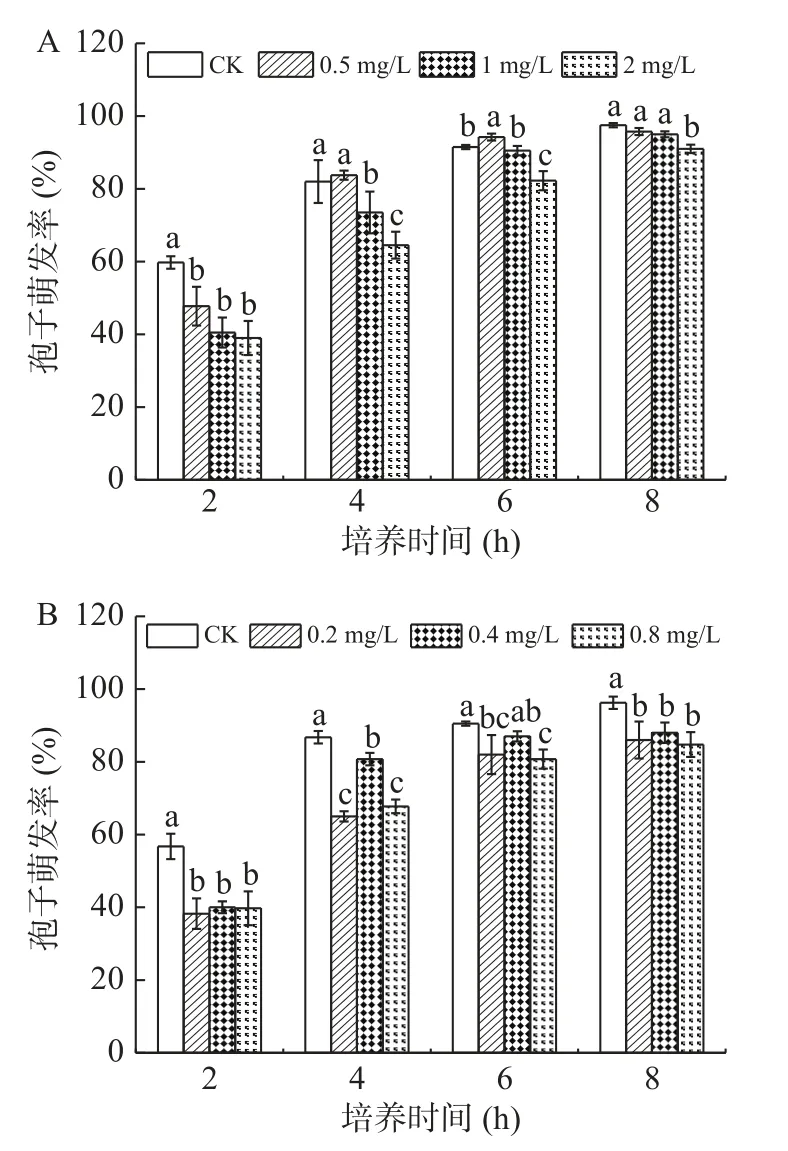
圖2 黃酮類處理對A. alternata孢子萌發的影響Fig.2 Effect of flavonoids treatment on spore germination of A. alternata
2.1.3 熊果苷處理對A. alternata孢子萌發的影響 熊果苷是蘋果梨果皮中含量最豐富的一種酚類物質。由圖3可知,在整個培養時間內,不同濃度熊果苷處理A. alternata后,對其孢子的萌發無抑制作用,與對照相比無明顯差異。
圖3 熊果苷處理對A. alternata孢子萌發的影響Fig.3 Effect of arbutin treatment on spore germination of A. alternata
2.1.4 酚酸類物質處理對A. alternata菌絲生長的影響 由圖4可知,在整個培養期間,A. alternata的菌絲生長受到酚酸類物質的顯著抑制,且隨著處理濃度的增加,抑制作用越強,在培養4 d后,高濃度酚酸類物質處理A. alternata后,其菌絲生長狀況與對照組相比,均有顯著地抑制作用,其中阿魏酸處理抑制作用最強,以4 mg/mL綠原酸、0.6 mg/mL咖啡酸、2 mg/mL阿魏酸和1.2 mg/mL對香豆酸處理組分別較對照降低了24.8%、6.5%、8.2%和29.4%。圖5表示A. alternata由4種酚酸類物質處理之后的菌落顏色為墨綠色,邊緣白色為菌絲,且圖片顯示4種酚酸類物質均不同程度得抑制了A. alternata的菌落生長,且統計結果為圖4所示。
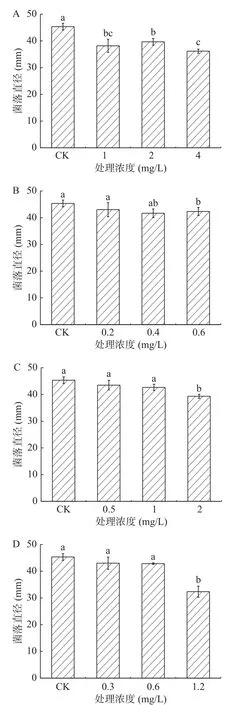
圖4 酚酸類物質處理對A. alternata菌絲生長的影響Fig.4 Effect of phenolic acids treatment on mycelia growth of A. alternata

圖5 酚酸類物質處理對A. alternata菌絲生長的影響Fig.5 Effect of phenolic acids treatment on mycelia growth of A. alternata
2.1.5 黃酮類物質處理對A. alternata菌絲生長的影響 黃酮類物質表兒茶素處理A. alternata后,培養4 d后,由圖6可知,其菌絲生長狀況與對照相比無明顯差異,而兒茶素處理后高濃度具有顯著地抑制作用,2 mg/mL處理后較對照降低了8.2%(P<0.05)。圖7則分別為兒茶素和表兒茶素處理之后A. alternata的菌落生長4 d的形態,通過圖片結果顯示,2種黃酮類物質處理之后,其中2 mg/mL的兒茶素處理抑制了A. alternata的菌落生長,表兒茶素處理之后對其抑制作用不明顯,但2種黃酮類物質處理之后,A.alternata的孢子產生均被抑制其中兒茶素抑制性較表兒茶素強,且具有濃度效應。
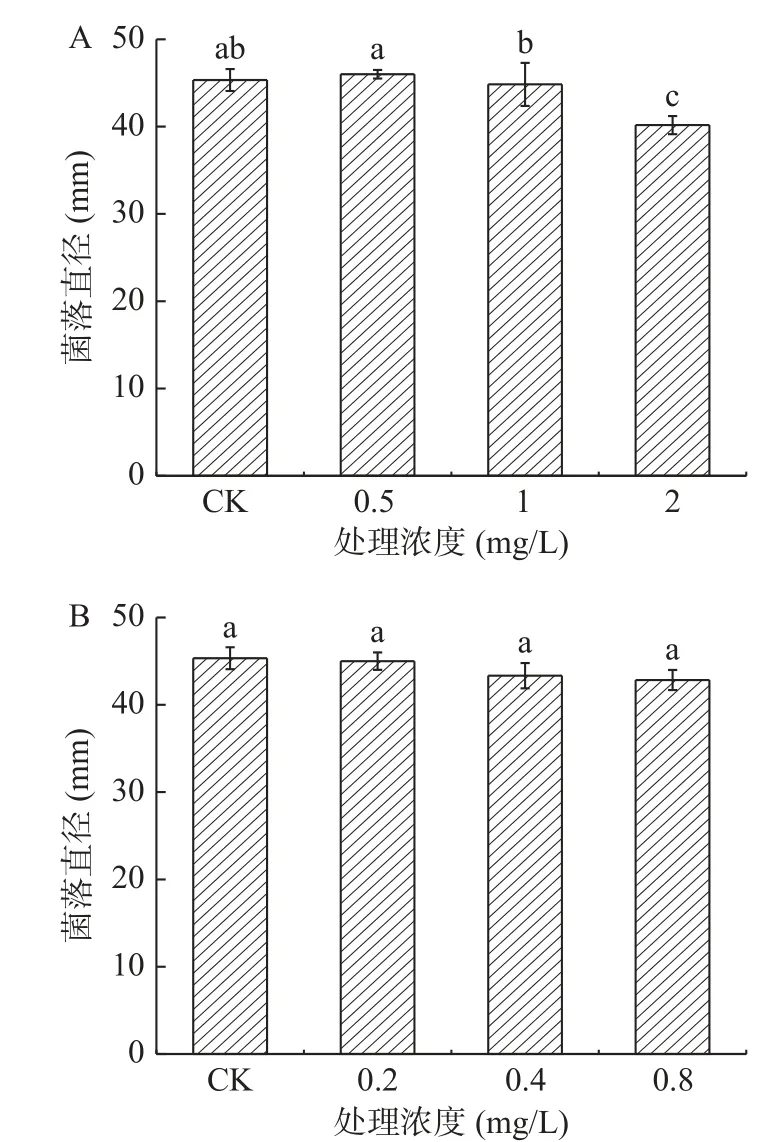
圖6 黃酮類物質處理對A. alternata菌絲生長的影響Fig.6 Effect of flavonoids treatment on mycelia growth of A. alternata
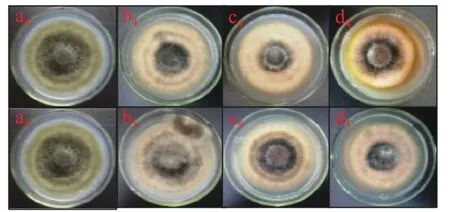
圖7 黃酮類物質處理對A. alternata菌絲生長的影響Fig.7 Effect of flavonoids treatment on mycelia growth of A. alternata
2.1.6 熊果苷處理對A. alternata菌絲生長的影響 由圖8可知,不同濃度外源熊果苷處理A. alternata后,培養4 d時,其菌絲生長狀況與對照相比無顯著性差異(P>0.05)。表明外源熊果苷處理后對A.alternata的菌絲生長無抑制作用。由圖9可以看出,熊果苷處理A. alternata后,菌落直徑與對照相比無明顯差異,但菌落顏色對照為墨綠色,處理組為褐色。
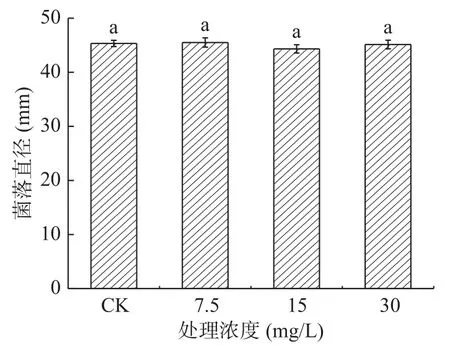
圖8 熊果苷處理對A. alternata菌絲生長的影響Fig.8 Effect of arbutin treatment on mycelia growth of A. alternata
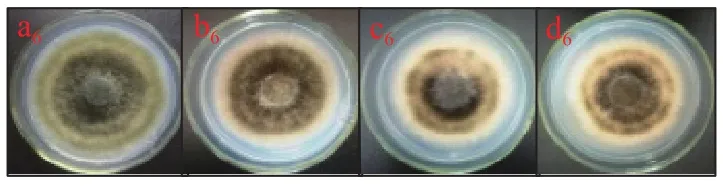
圖9 熊果苷處理對A. alternata菌絲生長的影響Fig.9 Effect of arbutin treatment on mycelia growth of A. alternata
2.2 蘋果梨果皮酚類物質對A. alternata生長的影響
2.2.1 酚酸類物質對A. alternata菌絲生物量的影響 不同濃度的外源酚酸類物質處理A. alternata后,均一定程度的降低了A. alternata的菌絲干重,且隨著濃度的增加菌絲生物量被降低的程度越強。在培養4 d后(圖10),以4 mg/mL綠原酸,0.6 mg/mL咖啡酸,2 mg/mL阿魏酸和1.2 mg/mL對香豆酸處理組較對照分別降低了52.9%、42.5%、22.2%和32.8%。
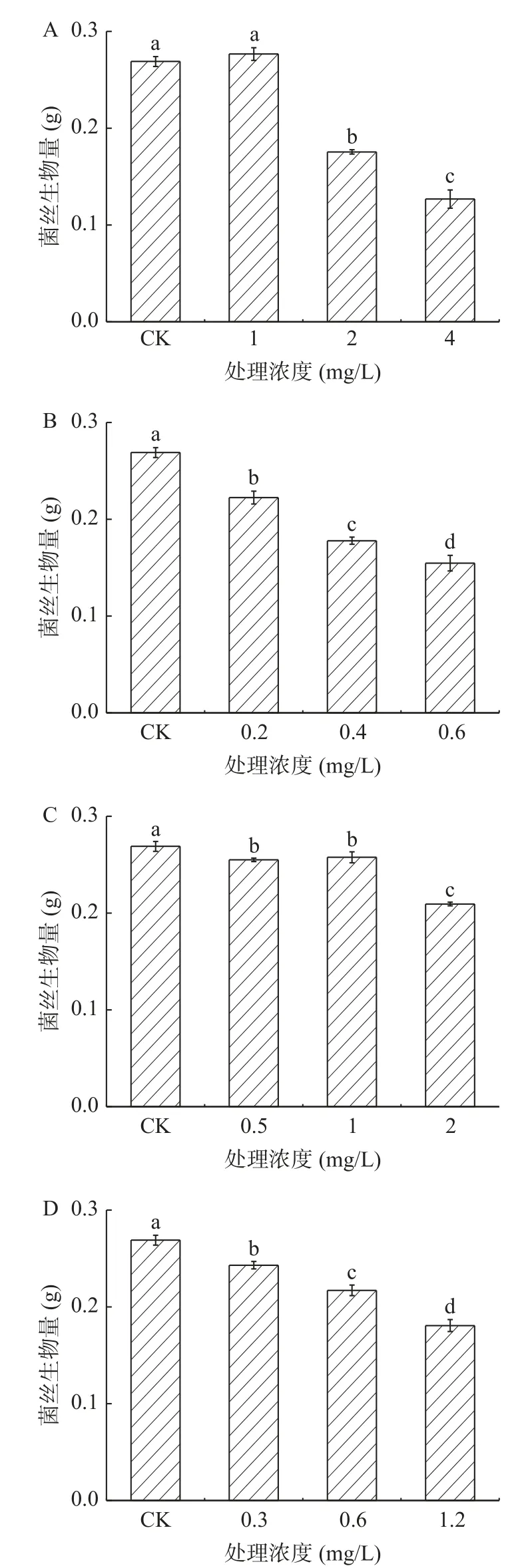
圖10 酚酸類物質處理對A. alternata菌絲生物量的影響Fig.10 Effect of phenolic acids treatment on mycelia biomass of A. alternata
2.2.2 外源黃酮類物質對A.alternata菌絲生物量的影響 由圖11可知,不同濃度兒茶素處理A. alternata后(圖11A),對其菌絲干重的影響程度不同,其中2 mg/mL處理后與對照相比菌絲生物量降低了18.1%。然而,不同濃度表兒茶素處理A. alternata后(圖11B),與對照相比菌絲生物量無明顯差異。
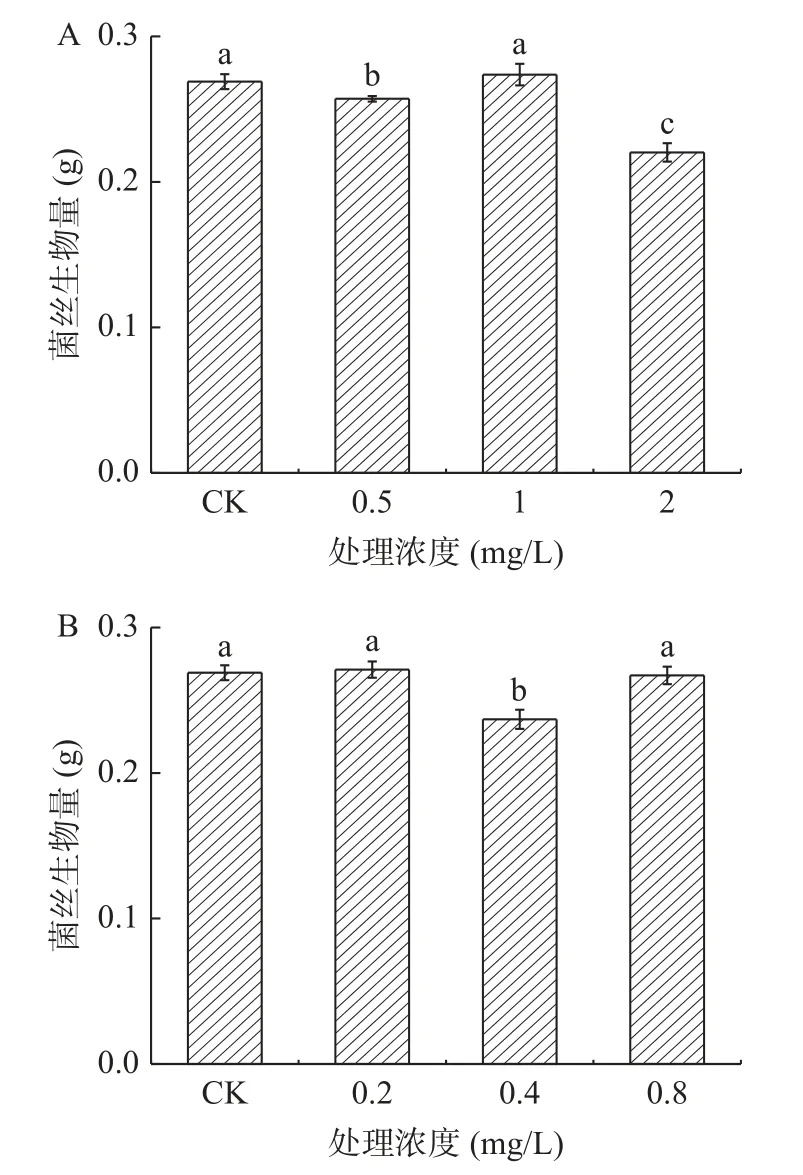
圖11 黃酮類物質處理對A. alternata菌絲生物量的影響Fig.11 Effect of flavonoids treatment on mycelia biomass of A. alternata
2.2.3 外源熊果苷處理質對A. alternata菌絲生物量的影響 在蘋果梨果皮酚類物質中,熊果苷含量最高。由圖12可知,培養4 d時,以不同濃度外源熊果苷處理A. alternata后,隨著熊果苷濃度增加,菌絲生物量含量逐漸下降。其中30 mg/mL熊果苷處理后較對照降低了19.6%。
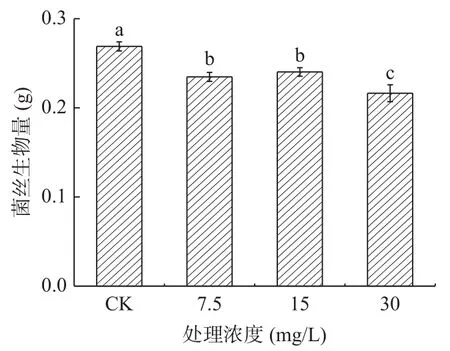
圖12 熊果苷處理對A. alternata菌絲生物量的影響Fig.12 Effect of arbutin treatment on mycelia biomass of A. alternata.
2.3 蘋果梨果皮酚類物質對A. alternata黑色素含量的影響
由表1可知,培養4 d時,不同濃度外源酚類物質處理A. alternata后,其黑色素含量均不同程度的增加,且總體上隨著酚類物質濃度的增加黑色素含量呈現增加的趨勢。其中兒茶素、表兒茶素和熊果苷對黑色素形成的促進作用最強,0.4 mg/mL 咖啡酸、2 mg/mL兒茶素、0.6 mg/mL表兒茶素和30 mg/mL熊果苷處理后,A. alternata黑色素含量分別是對照的2.53倍、3.00倍、2.05倍和1.53倍。但綠原酸、阿魏酸和對香豆酸處理A. alternata后,黑色素的含量略有增加。
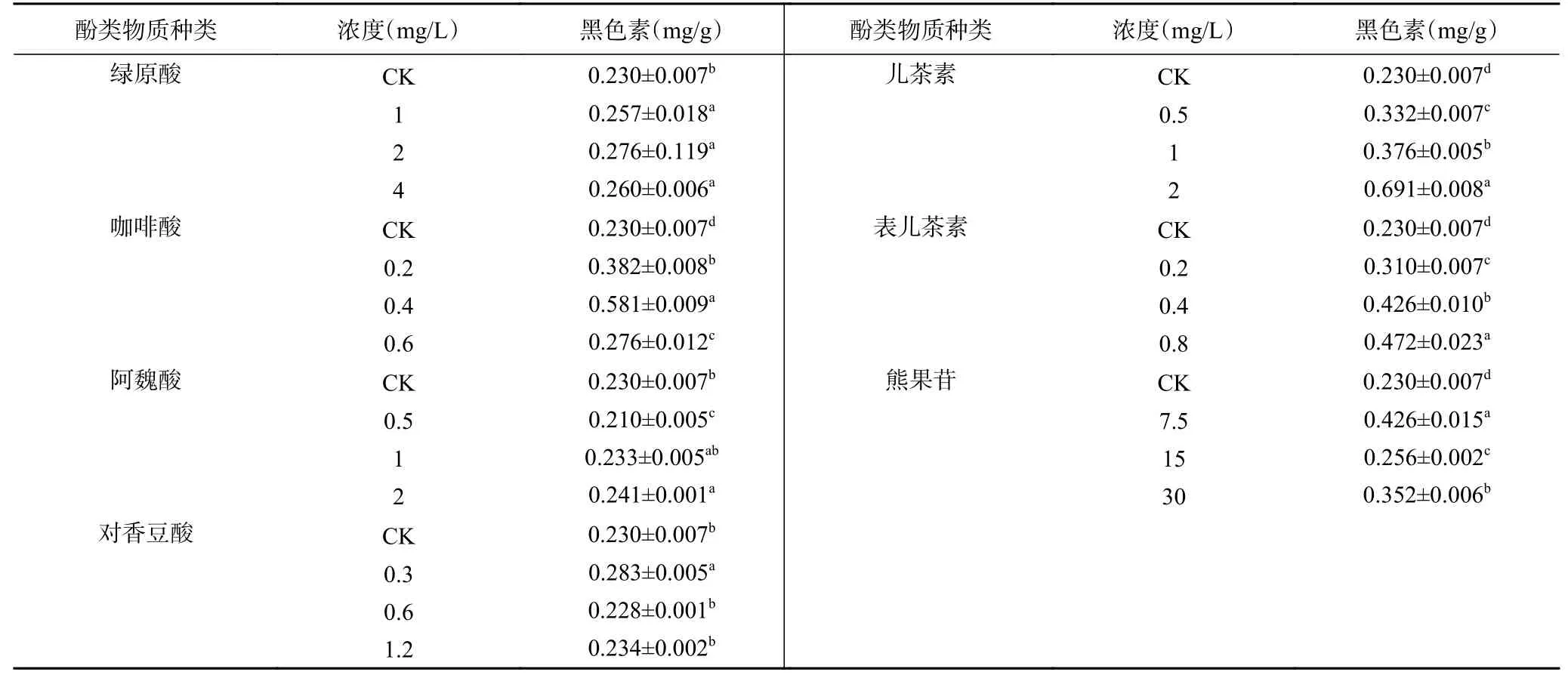
表1 不同酚類物質對A. alternata黑色素含量的影響Table 1 Effect of different phenolic compounds on the content of melanin in A. alternata
3 討論
本研究發現蘋果梨果皮的酚酸類物質顯著地抑制了A. alternata的孢子萌發、菌絲生長及生物量,這與Lee等[11]發現在未成熟的桃果實皮中的綠原酸對M. fructicola作用結果相一致。類似的如Sanzani等[3]和白雪娜等[17]研究發現傘形花內酯能有效抑制蘋果P. expansum、Botrytis cinerea和M. fructicola的生長。Li等發現800 μg/mL的對香豆酸甲酯(乙酯)顯著地延緩了A. alternata孢子萌發以及菌絲生長,進而延緩棗果實病害的發生[12?13]。本文研究結果顯示,阿魏酸和對香豆酸對A. alternata生長的抑制效果最佳,在處理8 h后發現,2 mg/mL阿魏酸和1.2 mg/mL對香豆酸處理的A. alternata孢子萌發率分別較對照降低了98.5%以及86.0%;培養4 d時,這兩個酚類物質處理組的A. alternata菌落直徑與對照相比分別降低了29.4%和8.2%,相類似的研究發現阿魏酸處理可有效控制西瓜采后病原真菌Fusarium oxysporum引起的枯萎病[18]。此外,本研究發現,2 mg/mL的兒茶素對A. alternata的生長同樣具有抑制作用,Pane等[19]在辣椒中也有類似發現。根據本研究的具體結果,分析可能的原因與酚類物質處理病原菌后破壞了其細胞膜完整性有一定的關聯,由于植物酚類物質的化學結構式上存在的羥基(-OH)連接在苯環或其復雜的芳香環上[20],因此其更具有親脂性,而羥基可作用于細胞膜通過氧化磷酸化解偶聯進而優先進入膜內[21]。由此酚類物質的抗真菌機制在于其影響病原物細胞脂蛋白膜的功能,損傷細胞膜完整性,細胞離子穩態以及液泡,胞質pH的變化[22-23],類似的研究發現水楊酸處理后損傷P. expansum細胞膜完整性,導致質膜蛋白質,糖以及核酸的滲透泄露[24]。
黑色素是病原真菌致病過程中重要的毒力因子,具有抗氧化、抗溶菌酶、吸收紫外線、抵御宿主免疫攻擊等生物學功能[25],雖然對真菌生長和繁殖的影響較弱,但能夠降低病原菌對寄主所分泌的代謝產物的敏感性,從而提高其在寄主中的生存能力[26],如Bacillus anthraci、Magnaporthe grisea、Acanthosporium cucurbitum、Acanthopanax senticosus和Acanthopanax graminearum等入侵寄主時產生的黑色素能激發附著胞產生膨壓,有利于病原菌在寄主體內定殖[27],同時,Pihet等[28]和Chai等[29]研究發現Cryptococcus neoformans黑色素缺失突變菌株喪失了致病性,在M. grisea[30]和A. alternata[31]中也發現了類似結果。而黑色素增強病原真菌的生命力主要是由于其具有菌體胞外氧化還原緩沖劑的作用,可以中和病原物侵入寄主時造成的由于環境壓力產生的氧化劑,并且研究觀察到,黑色的C. neoformans細胞在氮和氧衍生的氧化劑作用下比非黑化的細胞存活率高出約10倍,這表明黑色素通過保護病原體免受免疫產生的自由基而有助于其致病,同時在對Wangiella dermatidis以及A. alternata的氧化損傷研究時發現,黑色素在這兩種病原菌中的氧化還原性質與C.neoformans中發現的結果一致[32]。然而,酚類物質對病原菌黑色素的研究鮮有報道,我們研究發現,梨果皮的7種酚類物質處理A. alternata后,不同程度的刺激了該病原菌黑色素的產生,有趣的是,酚酸類物質處理后顯著地延緩A. alternata的生長,然而其對黑色素的產生無明顯影響,相反的是兒茶素、表兒茶素和熊果苷處理后使A. alternata生長過程中黑色素明顯增加,其高濃度處理后黑色素含量分別是對照的3.00倍、2.05倍和1.53倍。其具體調控機理尚需進一步研究。
4 結論
除熊果苷外,蘋果梨果的酚類物質處理對A.alternata的生長均具有不同程度的抑制作用,其中酚酸類物質的抑制效果較明顯,阿魏酸的抑制作用最強。此外,酚類物質處理后,均顯著地激發了A.alternata致病過程中黑色素的產生量,兒茶素、表兒茶素和熊果苷對A. alternata產生黑色素的促進作用較強。表明蘋果梨果皮酚類物質對A. alternata的生長以及致病性具有重要的調控作用。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
