轉小麥 TaPDI-A、 TaTRXh-A和 TaPP2Ac-D基因擬南芥植株的主要抗旱性研究
2022-05-20 08:15:50張云瑞楊燕寧田淑媛張雙星張曉科
麥類作物學報 2022年5期
張云瑞,魏 凡,楊燕寧,田淑媛,張雙星,張曉科
(西北農林科技大學農學院,陜西楊凌 712100)
小麥(L.)是世界上最重要的糧食作物之一,在生長發育過程中會面臨各種生物脅迫和非生物脅迫。干旱作為主要的非生物脅迫之一,嚴重影響作物的生長發育,是導致作物減產的重要原因。核氧還蛋白(nucleoredoxin,NRX)是硫氧還蛋白(thireodoxin,TRX)超家族的成員,在植物的生長發育、花粉萌發以及抗旱等方面發揮重要作用。研究表明,NRX可通過還原目標蛋白的二硫鍵來調控蛋白的生物活性。因此,研究NRX互作蛋白的抗旱性功能,對于解析NRX響應干旱脅迫的分子調控機制具有重要意義。
植物NRX蛋白根據TRX-like結構域的數量和氨基酸序列分為3大類:第Ι類NRX含有三個結構域,其中第一個和第三個結構域包含典型的WCG/PPC氧化還原位點,而第二個結構域是一個非典型的TRX結構域,沒有任何典型的TRX活性位點;第Ⅱ類和第Ⅲ類NRX包含兩個結構域,其中第Ⅱ類NRX包含非典型活性位點(WYP/AK/PC和W/R/HCL/A/V/RPC/G),第Ⅲ類NRX的第一個結構域包含一個高度保守的氧化還原位點(WCRPC),第二個結構域包含一個典型的TRX活性位點(WCPPC/F/S)。研究表明,小麥第Ι類NRX蛋白基因()受干旱脅迫誘導表達,與植物的抗旱性相關。程 潔等研究表明,TaNRX1蛋白具有體外TRX蛋白還原活性,并且其3個結構域均具有還原胰島素二硫鍵的活性。蛋白質并不是獨立在細胞中發揮其功能,通常與其他蛋白質相互作用形成復合體后才能完成特定的功能,比如某些賴于構象變化或翻譯后修飾的蛋白質功能只有在復合體形成后才能發揮出來。田淑媛等利用生物信息學方法篩選了TaNRX1的候選互作蛋白,然后經過酵母雙雜交和雙分子熒光互補驗證,發現小麥TaNRX1蛋白與蛋白質二硫鍵異構酶(protein disulfide isomerase,PDI)、h型硫氧還蛋白(h-type thioredoxin,TRXh)和蛋白磷酸酶2A催化亞基(protein phosphatase 2A catalytic subunit,PP2Ac)均存在互作關系,互作部位主要為細胞核和細胞膜,說明TaNRX1與其互作蛋白在生物體內廣泛存在,可行使多種生物學功能。研究表明,玉米和小麥基因在非生物脅迫下均上調表達,在硫氧還蛋白的氧化還原途徑中具有重要作用,并構成抗氧化防御系統的一部分;豌豆基因由氧化應激誘導表達,可以通過還原抗氧化蛋白參與豌豆組織中活性氧(ROS)的清除;小麥基因同樣受脅迫誘導表達,在植物對干旱脅迫的應答中起重要作用。
田淑媛等通過Wheat Exp數據庫預測了TaNRX1互作蛋白基因的表達量,的A基因組()和的A基因組()在干旱脅迫下表達量明顯升高。Xu等研究表明,基因具有抗旱功能。因此選擇、和三個基因進行后續試驗。田淑媛等發現,在酵母中過表達、和基因,緩解了干旱脅迫對酵母生長的抑制作用。但是蛋白的抗旱功能在微生物中和植物中可能存在一定差異,為了進一步驗證、和基因在植物中的抗旱功能,本研究在模式植物擬南芥()中分別對三者進行了抗旱功能驗證。通過農桿菌介導的遺傳轉化將普通小麥、和基因轉化擬南芥獲得過表達株系,以T代轉基因純合株系為材料進行抗旱性功能分析,以期為TaNRX1與其互作蛋白TaPDI、TaTRXh和TaPP2Ac的抗旱調控機制研究奠定基礎。
1 材料與方法
1.1 植物材料及培養條件
擬南芥()為Columbia生態型,由本實驗室保存。
1.2 過表達載體的構建
植物雙元表達載體pBI121由本實驗室保存。用限制性內切酶H I和I(NEB,美國)雙酶切線性化pBI121載體,依據前期克隆的小麥、和基因開放閱讀框(open reading frame,ORF)序列,設計帶有H I和I酶切位點的同源臂引物121-TaPDI-A-F/R、121-TaTRXh-F/R和121-TaPP2Ac-F/R(表1),利用ClonExpressII One Step Cloning Kit(諾唯贊,南京)將、和基因完整的ORF序列連接到線性化pBI121載體上。測序正確的重組載體分別命名為pBI121:、pBI121:和pBI121:,提取質粒保存備用。

表1 試驗所用引物
1.3 擬南芥的遺傳轉化與鑒定
將pBI121:、pBI121:和pBI121:重組載體轉化農桿菌GV3101,通過浸花法侵染擬南芥。T代擬南芥單株種子收獲后,利用卡那霉素(Kan)抗性篩選獲得T代轉基因擬南芥。T~T代擬南芥利用Kan抗性篩選和PCR檢測篩選陽性株系。PCR所用引物為121-TaPDI-A-F/R、121-TaTRXh-F/R和121-TaPP2Ac-F/R(表1)。PCR反應體系為20 μL,包括10 μmol·L的模板DNA 1.0 μL,10 μmol·L的上下游引物各0.5 μL,2 × Rapid Taq Master Mix 10.0 μL(諾唯贊,南京),ddHO 8.0 μL。PCR反應程序:95 ℃ 3 min;95 ℃ 15 s,70 ℃ 退火15 s,72 ℃ 延伸30 s,35個循環;72 ℃ 4 min。PCR產物在電壓120 V、電流250 mA條件下,利用1%的瓊脂糖凝膠電泳檢測是否含有目標條帶,用E.Z.N.A.Gel Extraction kit試劑盒(Omega,美國)膠回收純化后連接克隆載體pEAsy-Blunt Cloning Kit(全式金,北京),轉化大腸桿菌DH5α,挑單克隆進行測序。測序正確的陽性植株種子單株收獲,培養獲得轉基因擬南芥T代純合株系。本試驗涉及的引物合成和測序服務均由西安熱默爾生物科技有限公司完成。
1.4 轉基因擬南芥的抗旱性鑒定
幼苗期干旱脅迫后根長的測定:分別選擇3個T代轉、和基因擬南芥純合株系(用OE1~OE3表示),用甘露醇模擬干旱脅迫,以野生型作為對照鑒定干旱脅迫對根長的影響。擬南芥種子先用2%的NaClO溶液消毒10 min,蒸餾水沖洗3次后,再用70%酒精處理2 min,蒸餾水沖洗3次。將經過消毒處理的的擬南芥種子(野生型和轉基因株系)點播在1/2 MS培養基上,在光照培養箱內豎直培養3 d;挑選長勢一致的幼苗,移至1/2 MS培養基以及添加250 mmol·L甘露醇的1/2 MS培養基上;每個株系調查5株幼苗,每個處理重復3次,在光照培養箱(在光照強度為120 μmol·m·s,晝夜溫度為25 ℃/20 ℃,光暗周期為16 h/8 h)內豎直培養7 d后測量并記錄根長。
成株期抗旱性鑒定:擬南芥成株期抗旱性鑒定采用干旱脅迫復水法,以野生型為對照,將T代轉、和基因擬南芥株系OE1~OE3種子直接種植于營養土(基質∶蛭石=3∶1)中,置于光照培養箱中培養21 d后停止澆水,干旱脅迫21 d,待全部擬南芥都呈萎蔫狀態后恢復供水,復水5 d后計算存活率。使用IBM SPSS Statistics 20.0對數據進行統計分析。
1.5 轉基因擬南芥抗旱相關指標的測定
離體葉片失水率的測定:取在營養土中培養至4周齡的野生型和轉基因擬南芥,葉片離體后立即稱重(視為初始重量,W1),然后將樣品在室溫下放置0.5、1.0、2.0、4.0、5.0和6.0 h后稱重(W2),參照Srivastava等的方法進行測定,每個試驗設置3個生物學重復,計算相對于初始重量減輕的比例,失水率=(W1-W2)/W1×100%。
葉片MDA、脯氨酸及HO含量的測定:野生型和轉基因植株在營養土中生長至4周齡后,部分正常供水作為對照,部分停止澆水進行干旱脅迫,7 d后分別取正常供水和干旱脅迫的野生型和轉、、基因擬南芥測定葉片中MDA、脯氨酸和HO含量。MDA和脯氨酸含量的測定參照王學奎等的方法;HO含量的測定采用二氨基聯苯胺(DAB)染色法:取正常供水和干旱脅迫的擬南芥葉片浸沒在0.1 mg·mLDAB溶液中染色 4 h,用清水沖洗去除染色液后加入無水乙醇∶乙酸∶甘油=3∶1∶1(V/V/V),于沸水中脫色 15 min后記錄照相,根據DAB染色的深淺來推斷葉片中HO的含量。所有試驗至少重復3次,使用IBM SPSS Statistics 20.0對數據進行統計分析。
2 結果與分析
2.1 干旱脅迫對轉TaPDI-A、TaTRXh-A和TaPP2Ac-D基因擬南芥根長和存活率的影響
為了鑒定轉、和基因擬南芥的抗旱性,分別在苗期和成株期對擬南芥進行干旱脅迫處理。在不含甘露醇的1/2 MS固體培養基上,苗期野生型擬南芥的平均根長分別為3.08 cm,轉、和基因擬南芥的平均根長為 3.10、2.97和3.01 cm,野生型和轉基因擬南芥的根長之間無顯著差異;在含有250 mmol·L甘露醇的1/2 MS固體培養基上,野生型擬南芥的平均根長為1.52 cm,轉、和基因擬南芥的平均根長分別為2.24、2.42和2.37 cm,均極顯著大于野生型,增幅分別為47.4%、59.2% 和 55.9%(圖1)。
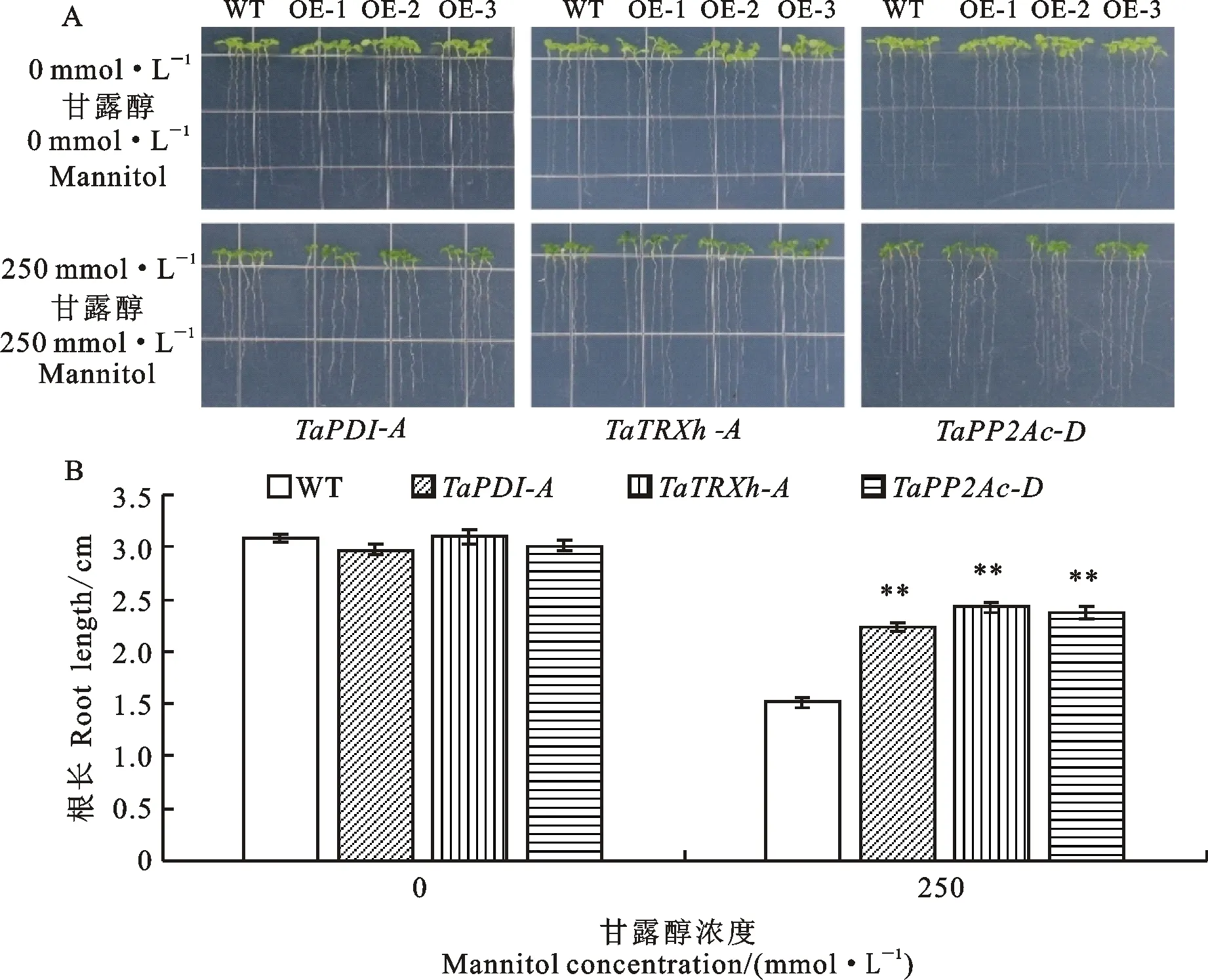
A:野生型和轉基因擬南芥的根長;B:根長的比較分析。WT:野生型擬南芥;OE-1~OE-3:轉基因擬南芥株系。圖4同。TaPDI-A:轉 TaPDI-A基因擬南芥;TaTRXh-A:轉 TaTRXh-A基因擬南芥;TaPP2Ac-D:轉 TaPP2Ac-D基因擬南芥。圖2~6同。**表示轉基因擬南芥與野生型擬南芥在0.01水平上差異顯著。圖2、圖3、圖5和圖6同。
成株期擬南芥在干旱脅迫前,野生型和轉基因擬南芥表型無明顯差異;干旱脅迫21 d后,野生型擬南芥萎蔫枯死嚴重,而轉、和基因擬南芥萎蔫枯死較輕;干旱脅迫復水后,轉、和基因擬南芥存活植株較多,存活率分別為93.3%、93.3%和66.7%,而野生型擬南芥的存活率僅為33.3%(圖2)。苗期和成株期的抗旱性鑒定結果均表明,過表達、和基因增強了擬南芥對干旱脅迫的耐受性。

A:野生型和轉基因擬南芥的干旱與復水表型;B:干旱脅迫復水后存活率。
2.2 干旱脅迫對轉 TaPDI-A、 TaTRXh-A 和 TaPP2Ac-D基因擬南芥離體葉片失水率的影響
從圖3可以看出,葉片離體0.5~6.0 h后,轉、和基因擬南芥離體葉片均表現出較低的失水率,顯著低于野生型,說明過表達、和基因降低了擬南芥在干旱脅迫下的葉片失水率,增強了擬南芥在干旱脅迫下的持水保水能力。

圖3 野生型和轉基因擬南芥離體葉片失水率
2.3 干旱脅迫對轉 TaPDI-A、 TaTRXh-A和 TaPP2Ac-D基因擬南芥葉片H2O2含量的影響
通過DAB對HO組織定位染色發現(圖4),正常供水條件下,野生型和轉基因擬南芥的葉片染色程度均較淺,兩者之間沒有明顯差異;干旱脅迫條件下,轉、和基因擬南芥株系的葉片染色程度比野生型擬南芥淺,表明轉基因擬南芥株系葉片中HO含量低于野生型。說明轉、和基因擬南芥在干旱脅迫下積累的HO較少。

圖4 正常和干旱脅迫條件下野生型和轉基因擬南芥的DAB染色結果
2.4 干旱脅迫對轉 TaPDI-A、 TaTRXh-A 和 TaPP2Ac-D基因擬南芥丙二醛(MDA)含量的 影響
從圖5可以看出,正常供水條件下,轉基因擬南芥與野生型擬南芥之間的MDA含量無顯著差異;干旱脅迫條件下,轉、和基因擬南芥的MDA含量均極顯著低于野生型。說明轉、和基因擬南芥在干旱脅迫下積累的MDA較少。

圖5 正常和干旱脅迫條件下野生型和轉基因擬南芥葉片的丙二醛(MDA)含量
2.5 干旱脅迫對轉 TaPDI-A、 TaTRXh-A 和 TaPP2Ac-D基因擬南芥脯氨酸含量的影響
從圖6可以看出,正常供水條件下,轉基因擬南芥與野生型擬南芥的脯氨酸含量之間無顯著差異;干旱脅迫條件下,轉、和基因擬南芥的脯氨酸含量均極顯著高于野生型。說明過表達、和基因增加了干旱脅迫后脯氨酸的積累,提高了擬南芥的滲透調節能力,對干旱脅迫的耐受性增加。

圖6 正常和干旱脅迫條件下野生型和轉基因擬南芥葉片的脯氨酸含量
3 討 論
3.1 小麥 TaPDI-A、 TaTRXh-A和 TaPP2Ac-D基因正調控擬南芥的抗旱性
植物可通過提高水分吸收率和減少水分流失率來提高抗旱能力,根系是植物吸收水分和養分的重要器官。本研究發現,干旱脅迫后,轉、和基因擬南芥的根長極顯著大于野生型,提高了轉基因擬南芥根系在干旱條件下的吸水能力,有助于植物抵御干旱脅迫。同時,轉、和基因擬南芥的離體葉片失水率顯著低于野生型,說明轉基因擬南芥可能通過減少葉片的水分流失率來應對干旱脅迫。植物適應干旱脅迫的另一個重要途徑是滲透調節,脯氨酸作為重要的滲透調節物質,當植物受到干旱脅迫后,體內會積累大量的游離脯氨酸,降低細胞內水勢,減少水分流失或增加水分吸收,從而有助于植物更好地抵御逆境脅迫。本研究結果表明,干旱脅迫后轉、和基因擬南芥植株中游離脯氨酸的含量增加,增強了對干旱脅迫的耐受性。前人研究表明,過表達基因增強了擬南芥幼苗對非生物脅迫的耐受性;擬南芥中基國在氧化脅迫條件下表達量明顯增加;小麥基因能夠被非生物脅迫因子誘導表達,且過表達的轉基因煙草比野生型具有更強的抗旱性。本研究發現,在擬南芥中過表達小麥、和基因,提高了擬南芥干旱脅迫下的存活率,正調控擬南芥的抗旱性。這與前人研究結果一致。
3.2 小麥 TaPDI-A、 TaTRXh-A和 TaPP2Ac-D調節活性氧的積累

4 結 論
本研究基于本課題組前期篩選的TaNRX1互作蛋白,通過農桿菌介導遺傳轉化擬南芥。干旱脅迫下,轉、和基因擬南芥的根長、存活率和脯氨酸含量均顯著大于野生型;葉片離體失水率、MDA含量和HO含量均低于野生型。說明TaNRX1的互作蛋白基因、和能夠顯著增強擬南芥的抗旱性。本研究豐富了對小麥TaNRX1互作蛋白基因抗旱功能的認識,為小麥抗旱遺傳改良提供了理論依據。
