非經典Hippo信號通路調控免疫穩態
2022-05-20 07:09:04史一然章津嘉陳蘭芬
廈門大學學報(自然科學版) 2022年3期
趙 昊,史一然,章津嘉,陳蘭芬
(廈門大學生命科學學院,細胞應激生物學國家重點實驗室,福建 廈門 361102)
生物體需要持續維持其內環境的穩態才能完成正常的生命活動.Hippo信號通路最早在果蠅(Drosophilamelanogaster)中發現,在進化上保守,是在控制器官大小和維持組織穩態中發揮重要作用的信號通路[1-3].哺乳動物中,Hippo信號通路的核心成員包括激酶Mst1/2(Ste20-like kinases 1/2,果蠅中Hippo的同源分子)、支架蛋白WW45(Salvador 同源物 SAV1)、激酶Lats1/2(large tumor suppressor 1/2)、支架蛋白MOB1(MOB kinase activator 1)和2個下游轉錄共激活因子YAP(Yes-associated protein)和TAZ(transcriptional coactivator with PDZ-binding motif).經典的Hippo信號通路是通過活化Mst1/2-WW45復合物后,進一步磷酸化和活化Lats1/2-MOB1A/B復合物,最終磷酸化YAP或TAZ,而磷酸化的YAP和TAZ會被降解或滯留在細胞漿中,無法進入細胞核與轉錄因子TEAD(transcriptional enhanced associate domain)結合,啟動下游基因轉錄(圖1(a)).很多研究表明Hippo信號通路在組織穩態、再生和腫瘤發生發展中起重要作用[4-11],如:在肝臟組織中,Hippo通路失活(敲除Mst1/2基因或過表達Yap基因)會導致肝臟增大和最終肝癌發生,而敲除Yap基因則會造成肝細胞死亡和肝臟衰竭.簡而言之,經典Hippo信號通路主要通過抑制YAP的功能來調控細胞增殖和死亡,維持組織器官的穩態平衡.最近,越來越多研究表明Hippo信號通路在維持免疫穩態平衡中也發揮重要作用.

FAT4.非典型鈣黏蛋白4;GPCR.G蛋白偶聯受體;FGFR4.成纖維細胞生長因子受體4;TLR.Toll樣受體;TCR.T細胞受體;BCR.B細胞受體;CaMKⅡ.鈣調蛋白依賴的蛋白激酶Ⅱ;IRAK1/4.白介素1受體相關激酶1/4;NFκB.核因子κB;PKC.蛋白激酶C;RAP1.Ras相關蛋白1;RAPL.Rassf5b,Ras相關結構域家族5b;LFA-1.淋巴細胞功能相關抗原1;DOCK8.胞質分裂作用因子8;Rac1.Rac家族小三磷酸鳥苷酶(GTPase)1;Keap1.Kelch類ECH相關蛋白l;AKT.蛋白激酶B;Sirt1.Sirtuin1;Stat5.信號轉換和轉錄激活蛋白5;ROS.活性氧;Nrf2.核紅細胞樣相關因子2;IRF3.干擾素調節因子3;Foxo1/3.叉頭框蛋白O1/3;Foxp3.叉頭框蛋白P3;RORγt.視黃酸相關孤兒受體γt.
1 Hippo信號通路與免疫穩態調控
免疫系統是機體識別和清除異己的重要防御系統,對于維持機體內環境穩態至關重要;在正常生理條件下,免疫系統本身的穩態維持對于其發揮正常免疫應答和免疫功能是必需的.免疫系統通過各種免疫細胞,包括天然免疫細胞和獲得性免疫細胞來持續性感受外環境,區分自己和異己,保護機體免受感染、自身免疫性疾病和腫瘤的侵擾[12-13].在外來抗原或自身細胞發生突變等刺激下,機體發生免疫應答,通過一系列合適的免疫反應使得機體盡快恢復內環境穩態平衡[13-14].當免疫應答過強或過弱時,都會造成機體穩態失衡,導致疾病的發生.免疫細胞可以通過相關受體感應外界環境、啟動相應信號轉導通路、產生下游效應分子,以及通過可能的反饋調節機制來進行免疫應答和維持機體的免疫穩態平衡.探究機體在生理和病理條件下維持免疫穩態平衡的分子機制,將有助于人們更好地理解自身免疫性疾病、感染、代謝失常和腫瘤等疾病的發病機制,為防治這些疾病提供新的思路和分子靶標.
最近研究發現,Hippo信號通路在維持免疫穩態平衡中也發揮重要作用,但與調控器官大小和組織穩態的經典Hippo信號通路不同,調控免疫系統功能的是以激酶Mst1/2為核心的非經典Hippo信號通路(圖1(b))[15].早期研究發現,Hippo信號通路中Mst1-Nore1b(又稱RAPL或Rassf5b)復合物是TCR或趨化因子激活后,LFA-1介導的T細胞黏附和遷移運動所必需的[16-17].激酶Mst1/2和其結合分子Nore1b以及底物MOB1在免疫組織和細胞中高表達[9].有意思的是,Mst1和Mst2雙基因敲除小鼠為胚胎致死表型,Mst1單基因敲除小鼠主要表現為免疫缺陷綜合征,而Mst2單基因敲除小鼠無明顯的免疫缺陷表型,表明Mst1在免疫系統中發揮著更為主導的功能,且Mst1和Mst2在功能上有一定互補性[9,18].臨床研究發現,激酶MST1功能缺失突變的病人表現為免疫缺陷綜合征,極易發生細菌、病毒和真菌感染,易得自身免疫性疾病等,且存在體內T細胞和B細胞減少、天然免疫細胞功能異常等臨床癥狀[19-20].在小鼠和病人中觀察到的表型和癥狀都暗示Hippo信號通路在天然免疫和獲得性免疫功能調節中發揮重要作用,而Mst1缺失導致的抗感染免疫缺陷,同時又有打破免疫耐受發生自身免疫性疾病的表型,也暗示Hippo信號通路在不同免疫細胞和不同免疫應激條件下可能發揮截然不同的調控作用.進一步研究發現,在免疫系統中,激酶Mst1/2主要通過與其他一些調控免疫功能的信號通路互作來調控機體的免疫應答,包括整合素(integrin)信號[16-17,21-22]、TCR和BCR信號[18,23-25]、細胞因子受體信號[26-27]、TLR信號[28-32]、有絲分裂原活化蛋白激酶(MAPK)信號[33-34]和抗病毒信號[35-37]通路等,統稱為非經典Hippo信號通路.
本研究團隊近年圍繞Hippo信號通路在免疫系統功能調控和穩態維持方面取得了一些進展.早期研究發現Mst1缺失會導致淋巴細胞減少,進一步研究發現在抗原或anti-CD3抗體刺激下,Mst1是調控T細胞增殖的內在抑制分子,對于維持機體T細胞數目非常重要[18,25].最近研究還發現在T細胞中,Mst1/2和其下游的TAZ分子能夠抑制輔助性T細胞17(Th17)和增強調節性T細胞(Treg)的分化,因此在T細胞中,恰當的Hippo信號通路調控對于防止自身免疫性疾病、維持免疫耐受至關重要[38].此外,在天然免疫細胞中,本研究團隊發現機械力受體Piezo1與TLR信號互作活化激酶Mst1/2,通過調控下游小G蛋白Rac1促進ROS產生,調控吞噬細胞對于細菌的吞噬和殺傷作用[29-30];同時激酶Mst1/2還可以感應細胞內的ROS水平,通過Mst1/2-Keap1-Nrf2信號轉導途徑來調控細胞的抗氧化活性,從而維持細胞的氧化還原穩態[39].
2 Hippo信號通路在T細胞中的功能
骨髓中的淋巴樣祖細胞在遷移到胸腺后,經過系列選擇過程,經歷CD4-CD8-雙陰性(DN)胸腺細胞、CD4+CD8+雙陽性(DP)胸腺細胞、CD4+或CD8+單陽性(SP)胸腺細胞階段,最終發育為成熟T細胞.成熟T細胞隨后進入血液循環,遷移到次級淋巴組織,接受抗原的刺激和活化.沒有接觸過抗原的T細胞稱為初始T細胞(na?ve T cell),在淋巴管、外周血和組織液中反復循環遷移,這有助于其廣泛接觸到可能的抗原,該過程對于加強免疫應答反應和維持長期的免疫記憶至關重要.大量研究表明Mst1在T細胞的遷移、歸巢、分化和活化中都發揮重要作用[16-18,21-22,25,27,38,40-42].
2.1 激酶Mst1/2調控T細胞的活化和遷移
在胸腺中,Mst1和Mst2蛋白在DP階段表達極低,而隨著發育進程表達量逐漸升高,最終在CD4+和CD8+的SP胸腺細胞中高表達;Mst1和Mst2的底物MOB1蛋白在DP和SP胸腺細胞中的表達量相當,但是磷酸化的MOB1只在SP胸腺細胞中被檢測到,該結果表明Mst1和Mst2在早期胸腺細胞發育中(DP胸腺細胞階段之前)的作用不大[25].在外周血中,剛從胸腺中成熟遷出的初始T細胞高表達Mst1和Mst2;而與CD4+CD62Lhigh初始T細胞相比,在CD4+CD62Llow效應/記憶T細胞中,Mst1和Mst2的蛋白和mRNA水平約下降90%[25].結合本研究團隊在Mst1敲除小鼠中觀察到的T細胞高度活化和CD4+CD62Llow效應/記憶T細胞比例增加的表型,可以推斷Mst1和Mst2蛋白水平的下降是初始T細胞轉變為活化效應/記憶T細胞所必需的,T細胞中Mst1的活性水平決定了初始T細胞活化的閾值;與Mst1單基因敲除的表型不同,Mst2單基因敲除小鼠并不影響各組織中的T細胞數目;然而在Mst1敲除小鼠中,進一步在造血細胞系中條件性敲除Mst2則會加重T細胞減少的表型,該結果表明激酶Mst2在Mst1敲除小鼠的淋巴組織中發揮代償性功能;其中,激酶Mst1活性對于T細胞數目和功能穩態是必需的,因為只有野生型Mst1蛋白而非激酶缺陷型的Mst1蛋白的轉基因小鼠,才可以恢復Mst1缺失導致的T細胞缺陷表型[18].
與野生型小鼠相比,Mst1-/-Mst2fl/flVav-Cre小鼠(全身敲除Mst1,在造血系統中條件性敲除Mst2)的CD4+和CD8+SP胸腺細胞比例增多,這些SP胸腺細胞發生大量的凋亡事件,雖已成熟卻無法遷移出胸腺[18].Qa淋巴細胞抗原2(Qa-2)分子在胸腺細胞發育晚期高表達,在成熟胸腺細胞離開胸腺和成為外周成熟T細胞時表達量繼續升高;而CD24分子則相反,在胸腺細胞發育期高表達,而在成熟胸腺細胞離開胸腺時表達量下降,在外周成熟T細胞中不表達[43].因此Qa-2和CD24可以用來指示胸腺細胞和外周循環T細胞的表面標記.Mst1-/-Mst2fl/flVav-Cre小鼠的CD4+和CD8+SP胸腺細胞主要表現為Qa-2highCD24low表型,同時高表達S1P(sphingosine 1-phosphate)受體(主要表達于胸腺細胞發育晚期),說明這些細胞都是被滯留于胸腺中的成熟T細胞[18].這些Mst1/2缺失的SP胸腺細胞表面的趨化因子受體(如CCR7、CXCR3、CXCR4和CCR5)和整合素(CD11b和LFA-1)的表達水平與野生型細胞相當,但如果將這些細胞放于外周循環中,同樣無法遷移進入二級淋巴組織,表明Mst1/2缺失的胸腺細胞可以發育成熟為初始T細胞,但這些細胞無法遷出胸腺組織;進一步研究發現,在成熟的胸腺細胞中,趨化因子CCL19或S1P刺激下,Mst1/2通過磷酸化MOB1,促進MOB1與DOCK8的相互作用,增強其鳥苷轉換因子(GEF)活性,促進小G蛋白Rho家族成員(如Rac1)的活化,從而促進胸腺細胞遷出胸腺[18].
在Mst1-/-Mst2fl/flVav-Cre小鼠和Mst1-/-Mst2fl/flLck-Cre小鼠(全身敲除Mst1,在胸腺發育期條件性敲除Mst2)都出現成熟胸腺細胞遷出胸腺缺陷、外周T細胞減少和高度活化的表型[18].為了進一步研究Mst1/2在外周T細胞中的功能,構建了僅在成熟和活化的T細胞中敲除Mst1和Mst2的Mst1fl/flMst2fl/flOx40-Cre小鼠,結果顯示這些小鼠的胸腺細胞發育正常,外周組織中T細胞數目也正常,但與野生型小鼠相比,在脾臟中效應/記憶T細胞比例增加,說明Mst1/2缺失促進了T細胞活化[38].在TCR刺激下,Mst1缺失的初始T細胞的增殖水平和細胞因子白介素-2(IL-2)、IL-4和干擾素γ(IFNγ)的表達水平都明顯比野生型細胞的高,但TCR下游的CD3、ZAP70(zeta chain of TCR associated protein kinase 70)、Lck(lymphocyte-specific protein tyrosine kinase)和PLCγ1(phospholipase C-γ1)的磷酸化水平卻與野生型細胞的相當[25].在敲除Mst1的T細胞中,Lats1/2的磷酸化水平沒有變化,而MOB1的磷酸化完全消失,該結果暗示了MOB1可能在T細胞活化中發揮重要作用[25],而本研究團隊發現敲除YAP并不能緩解Mst1缺失導致的缺陷表型,進一步暗示Mst1在T細胞中不是以經典Hippo信號通路來發揮作用的(未發表數據).在TCR刺激下,Mst1缺失的效應/記憶CD4+T細胞的凋亡比例大幅增加,說明Mst1在這些T細胞中具有抗凋亡活性,Mst1缺失會增強活化誘導的細胞死亡(AICD)效應[25].總之,這些結果表明Mst1/2在維持T細胞數目、遷移和活化方面都發揮重要的調控作用.
2.2 Mst1/2-TAZ信號調控CD4+ T細胞的分化命運
在TCR/抗原刺激活化和細胞因子等不同因素的調控下,初始CD4+T細胞會分化為不同亞型的T細胞,在免疫應答和調控中發揮不同的作用[44].其中,接受TCR/抗原刺激的初始CD4+T細胞在轉化生長因子β(TGF-β)誘導下可分化為Treg,分泌TGF-β,發揮調控免疫耐受和免疫抑制的功能;而在TGF-β和IL-6的共同誘導下則分化為CD4+Th17,分泌IL-6和IL-17,參與防御胞外細菌和真菌感染、炎癥反應和自身免疫反應.Treg和Th17比例的恒定對于維持機體免疫系統的穩態起著非常重要的作用,初始CD4+T細胞向Th17分化過多或向Treg分化過少,是導致多種自身免疫性疾病的主要原因.Treg和Th17亞型的分化是交互調控的,其中TGF-β在調控CD4+初始T細胞分化為Treg和Th17中發揮重要作用,而在TGF-β培養條件下添加IL-6會抑制Treg的分化,促進Th17的分化,這有助于機體在應對感染等外界侵擾時,促進機體從免疫耐受到免疫炎癥反應的轉化[44].研究發現,與野生型小鼠相比,6~8周Mst1-/-小鼠的胸腺和脾臟中CD4+Treg明顯減少,調節功能缺陷[41,45].進一步研究發現Mst1通過多種方式調控和促進Treg中轉錄因子Foxp3的表達和功能,包括直接磷酸化轉錄因子Foxo1/3[41]、激酶AKT[41]和去乙酰化酶Sirt1[45]或調控Rac1-DOCK8信號通路[27].
最近,本研究團隊發現[38]:Mst1-/-小鼠的Th17分化比例顯著升高,小鼠易患干燥綜合征和大腸炎.用鑰孔血藍蛋白(KLH)抗原和弗氏完全佐劑(CFA)誘導小鼠的免疫應答,在Mst1fl/flMst2fl/flOx40-Cre小鼠的引流淋巴結中,Th17比例顯著高于野生型小鼠中,且Treg比例也明顯下調.分析Hippo信號通路主要成員在幾種CD4+T細胞亞型中的表達水平,發現激酶Mst1/2下游的轉錄共激活因子TAZ在Th17和Treg中表達水平都明顯高于初始CD4+T細胞中,且Th17的高表達更為顯著.臨床類風濕性關節炎等病人的外周血記憶T細胞中TAZ的表達水平高于健康人中,且與Th17的核心轉錄因子RORγt水平呈正相關.機制研究顯示,在TCR刺激初始CD4+T細胞時,TGF-β單獨刺激能夠大幅提升TAZ mRNA的表達水平;IL-6單獨刺激雖無法提高TAZ mRNA的表達水平,但能夠進一步加強TGF-β誘導TAZ mRNA的表達,且TGF-β和IL-6通過其下游轉錄因子Smad3和Stat3來調控TAZ的轉錄表達.
在調控Th17和Treg分化方面,本研究團隊發現TAZ能夠與Th17及Treg中的核心轉錄因子RORγt和Foxp3形成復合物[38]:首先,作為轉錄共激活因子,TAZ可以直接增強RORγt的轉錄活性;其次,由于Foxp3會被乙酰轉移酶Tip60(HIV-1 TAT-interactive protein,60 ku)乙酰化而抑制其泛素化和蛋白酶體降解[46],TAZ通過與Tip60競爭性結合Foxp3,導致Foxp3乙酰化降低、泛素化增強和蛋白降解增加,從而削弱Treg的分化、促進Th17的分化.因此,Mst1/2-TAZ信號在調控初始CD4+T細胞分化為Th17和Treg的過程中起關鍵作用(圖2),也證明了Hippo信號通路在免疫系統穩態調控中的重要性[38].
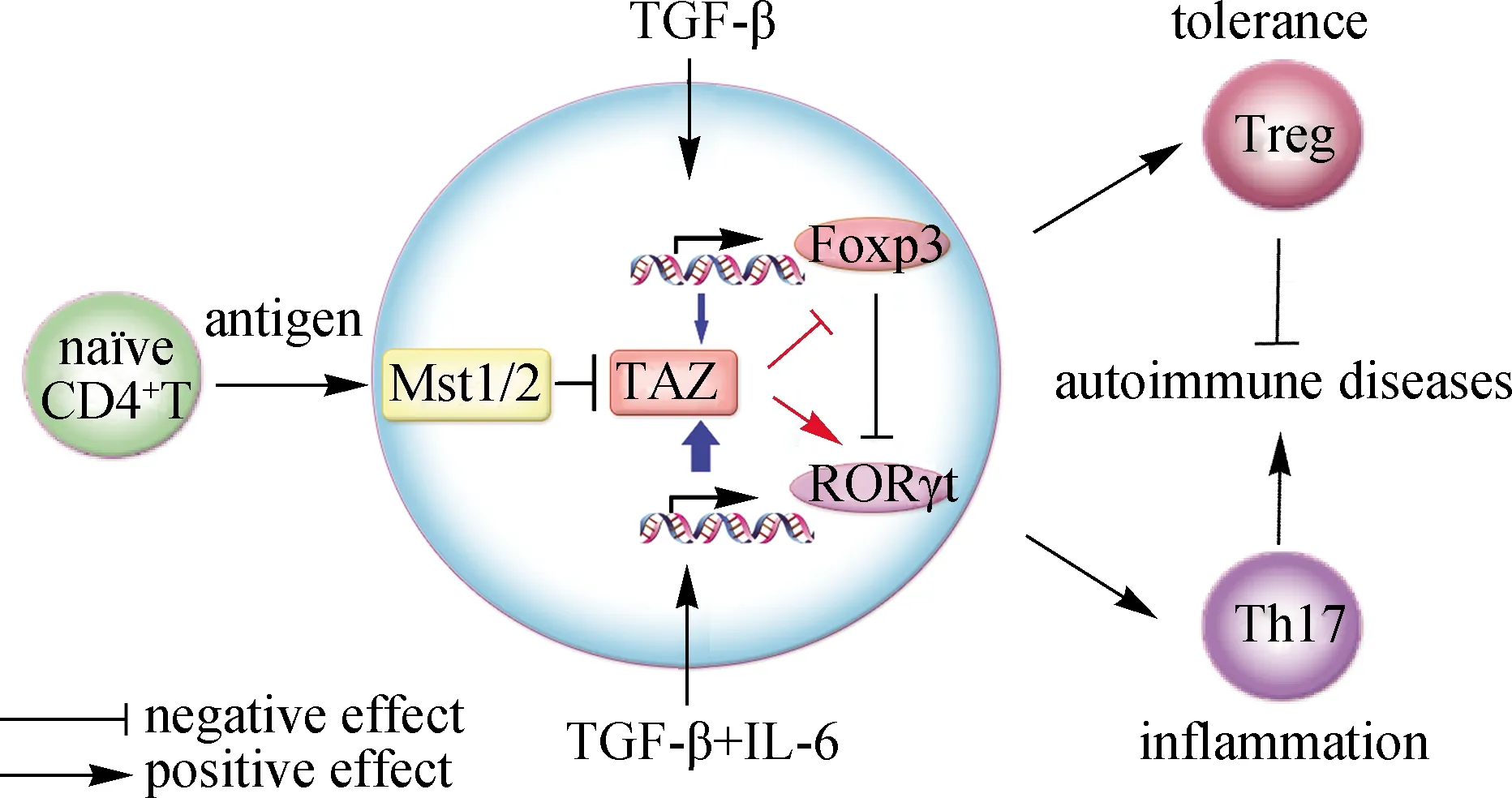
在Th17和Treg的分化過程中,TGF-β誘導TAZ的表達,而IL-6進一步增加TAZ的表達水平;作為轉錄共激活因子,TAZ會增強Th17中核心轉錄因子RORγt的轉錄活性以促進Th17的發育分化;TAZ也可通過調控Treg核心轉錄因子Foxp3的乙酰化和泛素化水平,促進Foxp3的降解,抑制Treg的分化.因此,Hippo信號通路可以調控機體免疫炎癥和免疫耐受的穩態平衡.
3 Hippo信號通路在天然免疫細胞中的功能
天然免疫系統是機體區分自我和非我,以應對外環境侵擾(如各種病原感染)并在第一時間作出免疫反應和應答的重要防線.單核巨噬細胞是最重要的引發炎癥反應的天然免疫細胞,其與另一類重要天然免疫細胞中性粒細胞均屬吞噬型細胞,主要發揮吞噬和清除病原體的功能.當病原體入侵機體時,天然免疫細胞通過模式識別受體,如TLR、C型凝集素樣受體、清道夫受體和補體受體等,來識別表達于病原體中的抗原相關的模式分子,如脂多糖(LPS)、脂蛋白、多肽和核酸分子等[47].
TLR可以識別多種不同的病原相關模式分子,在天然免疫細胞廣泛表達,是抵御病原入侵的第一道防線,也是連接天然免疫應答和獲得性免疫應答的橋梁[12].本研究團隊用各種TLR的配體刺激野生型小鼠骨髓來源的巨噬細胞(BMDM),發現表達于細胞表面的TLR1、TLR2和TLR4在相應配體或激動劑刺激活化后,都會導致Mst1/2底物MOB1的磷酸化水平明顯升高;表達于內體上的TLR7、TLR8和TLR9在相應配體或激動劑刺激后,MOB1的磷酸化水平沒有變化;而同樣表達于內體上的TLR3在聚肌胞苷酸poly(I:C)刺激下,MOB1的磷酸化水平略有升高[30].利用敲除MyD88(myeloiddifferentiationprimaryresponsegene88)的單核巨噬細胞系RAW264.7,進一步研究發現LPS(TLR4激動劑)、Pam3CSK4(TLR1和TLR2激動劑)和LTA(TLR2激動劑)激活Mst1/2依賴于MyD88信號,Mst1/2缺失并不影響TLRs介導的MAPK(包括p38、Jnk和Erk)的活化;然而與野生型BMDM相比,在Mst1/2缺失的BMDM中,LPS刺激導致更高的IKKα/β(IκB kinase α/β)和IκBα(inhibitor of NFκB kinase)磷酸化水平,以及相應更高的IL-6和腫瘤壞死因子α(TNFα)表達水平[30].此外,Mst1可以通過磷酸化和降解IRAK1調節TLR信號通路,抑制慢性炎癥反應和肝癌發展[31].同樣,結核桿菌(Mycobacteriumtuberculosis)也可以通過TLR2-IRAK1/4信號通路來調控Mst1/2的活性,從而調控趨化因子CXCL1/2的表達和IRF3介導的免疫應答[48].而在果蠅中的研究發現:Hippo信號是果蠅抵御革蘭氏陽性菌和真菌感染所必需的,果蠅脂肪體(果蠅的免疫器官)中Hippo信號缺失的免疫表型與Toll受體信號缺失的表型非常類似,說明在果蠅脂肪體中經典Hippo信號通路在調控天然免疫功能中發揮重要作用;當Hippo信號通路失活時,下游轉錄共激活因子Yorkie活化,可以直接促進Cactus(果蠅中的NFκB抑制蛋白IκB)的轉錄表達,從而抑制Dorsal(果蠅中的NFκB)和Dorsal相關免疫分子(Dif)入核以及抗菌肽的表達[32].而在哺乳動物中,研究發現在LPS和IFNγ刺激下,YAP在巨噬細胞中的表達水平升高促進巨噬細胞向M1型極化以及促炎癥因子IL-6的表達,而IL-4和IL-13刺激則抑制YAP的表達,因此巨噬細胞中特異性敲除YAP可明顯減輕葡聚糖硫酸鈉鹽誘導的腸炎發生[49].這些結果都表明Hippo信號通路在進化上是保守的,但在高等哺乳動物中有著更為復雜的調控機制來調節天然免疫應答.
3.1 激酶Mst1/2調控吞噬細胞的細菌吞噬和殺傷作用
Mst1功能缺失的病人易發生細菌、病毒和真菌的多發感染性疾病[19-20].本研究團隊前期發現Mst1-/-Mst2fl/flVav-Cre小鼠也經常發生肺炎、肺膿腫等多發感染性.與Mst1-/-Mst2fl/flVav-Cre小鼠不同,Mst1fl/flMst2fl/flLyz2-Cre小鼠(在髓系細胞中條件性敲除Mst1和Mst2)在正常SPF(specific pathogen free)條件下飼養到7個月,均未發現有炎癥或感染癥狀;但在Mst1fl/flMst2fl/flLyz2-Cre小鼠中構建盲腸穿刺誘導的細菌性腹膜炎(CLP)模型時,與野生型小鼠相比,模型小鼠的死亡率更高,各組織中的細菌載量更多,血清中的炎癥因子水平也更高[30].進一步研究發現TLR-MyD88信號介導的Mst1/2活化是吞噬型細胞(如巨噬細胞和中性粒細胞)吞噬和殺滅細菌所必需的[30];在大腸桿菌(Escherichiacoli)或李斯特菌(Listeriamonocytogenes)感染時,Mst1/2缺失的BMDM或中性粒細胞中吞噬的細菌比野生型細胞中少,但在感染后期胞內留存的細菌數卻明顯更多,這表明Mst1/2缺失影響了吞噬型細胞對細菌的吞噬和殺傷作用.
ROS在吞噬細胞的細菌殺傷作用中扮演著重要的角色,這些ROS主要來源于吞噬體上還原型輔酶Ⅱ(NADPH)氧化酶復合物產生的ROS和募集的線粒體產生的線粒體ROS(mROS).在Mst1/2缺失的吞噬細胞中,細菌感染后誘導產生吞噬體ROS和mROS的能力有明顯缺陷,是導致細菌殺傷能力下降的主要原因;此外,膜表面的TLR1、TLR2和TLR4活化后可以誘導產生大量mROS和胞內總ROS,而表達于內體上的TLR3、TLR7、TLR8和TLR9活化后則無法誘導相應的ROS產生;有意思的是,用TLR1、TLR2和TLR4相應的激動劑刺激Mst1/2缺失的BMDM或中性粒細胞,均無法誘導mROS和胞內總ROS的產生,這與之前研究發現刺激這些膜上TLR才會激活Mst1/2的結果一致[30].在機制上,Mst1/2的磷酸化激活PKC,活化的PKC進而磷酸化其底物二磷酸鳥苷(GDP)解聚抑制劑LyGDI,活化小G蛋白Rac1,促進TRAF6-ECSIT(TNF receptor associated factror 6-evolutionarily conserved signaling intermediate in Toll pathway,mitochondrial)復合物形成,最終促進募集線粒體靠近吞噬體,并釋放mROS殺滅吞噬體中的細菌;而同時Rac1是NADPH氧化酶復合物的主要成員之一,因此活化的Rac1也會促進吞噬體ROS的產生,進一步增強吞噬體中ROS水平以殺滅細菌[30].該研究揭示了一條全新的信號轉導通路,即TLR-Mst1/2-PKC-Rac1-TRAF6-ECIST介導吞噬細胞中細菌誘導ROS產生的分子機制[30],表明TLR下游信號并非只有激活相關轉錄因子、促進炎癥因子基因轉錄表達的作用,還可以通過蛋白質翻譯后修飾作用活化小G蛋白,影響細胞骨架快速調控細胞器(吞噬體和線粒體)的遷移,從而實現抗感染天然免疫應答.
3.2 機械力受體Pizeo1調控Mst1/2促進巨噬細胞抗感染免疫
感染、炎癥和腫瘤發生會導致組織中細胞間質的硬度、結構和成分等物理微環境的改變,研究發現細胞微環境中這些物理和機械信號可以調節多種細胞類型的分化、增殖和遷移[50].作為重要的天然免疫細胞,巨噬細胞持續穿梭于體液和各組織中以應對這些復雜的微環境,通過細胞本身硬度和彈性的內在變化來感知物理微環境變化.已有研究發現,貼壁細胞的生長密度和培養基硬度可以調控Hippo信號通路活性,細胞生長密度較低時或細胞培養在硬度高的介質上,都會抑制Hippo信號通路活性,導致下游分子YAP/TAZ進入細胞核,促進細胞生長,因此YAP/TAZ是兩個極為關鍵的受機械力調控的轉錄效應分子,可以將物理信號最終轉化為基因表達[51].
本研究團隊前期發現,TLR活化信號會激活下游的Mst1/2,且這一活化過程依賴于MyD88分子,但TLR-MyD88復合物如何激活Mst1/2并不明確[30].最近本研究團隊發現,培養在軟基質上的巨噬細胞的吞噬和殺菌功能被嚴重削弱,外環境機械硬度信號是巨噬細胞實現細菌吞噬和殺傷所必需的[29].機械敏感離子通道(MSC)中Piezo1離子通道是近年發現的在血管發育、紅細胞體積調節、細胞遷移等方面發揮重要作用的Ca2+通道[52].本研究團隊的結果顯示[29]:Piezo1受體在多種髓系免疫細胞中高表達,在細菌感染或LPS等一些TLR配體刺激下,其表達量會進一步升高.與野生型小鼠相比,天然免疫細胞中敲除Piezo1的Piezo1fl/flLyz2-Cre小鼠在CLP模型中生存率明顯降低,各組織中細菌載量顯著增加;而與Mst1/2缺失的BMDM表型類似,Piezo1缺失的BMDM的細菌吞噬和殺傷清除能力明顯低于野生型細胞.使用Yoda1(Piezo1激活劑)處理能夠顯著增強野生型BMDM的細菌吞噬和清除能力,而對Piezo1缺失的BMDM則沒有影響,說明活化Piezo1能夠促進巨噬細胞的抗感染能力;細菌感染或LPS刺激會促進TLR4和Piezo1發生組裝,從而增強依賴于Piezo1的鈣內流和CaMKⅡ活化,CaMKⅡ再通過磷酸化Hippo信號通路的Mst1/2介導Rac1活化,促進絲狀肌動蛋白重塑、增強吞噬作用、促進線粒體和吞噬體ROS的產生,從而增強殺菌能力.該結果表明機械力感受器Piezo1是激活Mst1/2-Rac1抗感染信號和誘導殺傷性ROS產生的關鍵上游分子[29].近期也有研究發現Piezo1可以調控巨噬細胞吞噬清除衰老紅細胞和鐵代謝[53],以及巨噬細胞通過Piezo1感受周期性的靜水壓變化[54],從而將胞外信號轉化為胞內信號調控細胞的炎癥反應.這些研究揭示了機械力感知和應答信號在巨噬細胞抗感染中的作用和機制,表明Piezo1所介導的機械力傳導信號對于宿主抗病原體的天然免疫應答至關重要.
3.3 Mst1/2調控巨噬細胞的氧化還原穩態
ROS在維持細胞正常的生理功能中發揮重要功能,是免疫吞噬細胞殺傷病原和激活炎癥反應的主要武器,但過量的ROS也會對細胞造成氧化應激損傷而導致衰老和死亡,因此調控ROS的產生和清除維持細胞的氧化還原穩態至關重要.在體外研究中,常使用過氧化氫溶液來激活Mst1/2,表明ROS可以有效激活激酶Mst1/2[25].早期研究發現在線蟲(Caenorhabditiselegans)中敲除CST-1(Hippo激酶的同源分子)會加速衰老和縮短壽命[55].本研究團隊前期發現,Mst1/2是巨噬細胞在細菌感染時誘導產生大量殺傷性ROS所必需的,然而同時Mst1/2缺失的巨噬細胞中ROS本底水平明顯高于野生型細胞[30].在這些Mst1/2缺失的巨噬細胞中,蛋白的羰基化水平(氧化應激標記)、組蛋白H2A變體H2A.X的磷酸化水平(DNA損傷標記)、PARPγ(poly ADP-ribose polymerase γ)和Caspase3切割分子的水平升高,同時細胞出現提早衰老的特征,而這些現象都可以在ROS清除劑N-乙酰半胱氨酸(NAC)的處理下得到緩解,表明Mst1/2在調節巨噬細胞氧化應激穩態維持中也發揮重要作用[39].研究結果顯示[39]:Mst1/2缺失的巨噬細胞中抗氧化酶系統存在缺陷,在LPS、抗霉素(atimycin A)、魚藤酮(rotenon)、過氧化氫處理或大腸桿菌感染下,Mst1/2缺失的BMDM中抗氧化酶基因(如Nqo1、Ho-1、Gclc和Gclm等)的表達水平都明顯低于野生型BMDM中.這些抗氧化基因多由抗氧化效應轉錄因子Nrf2調控表達,而Nrf2在巨噬細胞中高表達;與野生型細胞相比,Mst1/2缺失的巨噬細胞中Nrf2泛素化水平明顯升高,蛋白水平顯著降低.進一步研究表明Mst1/2能夠感應ROS,并被募集到吞噬體和線粒體的ROS產生部位而被釋放的ROS活化,進而調控Nrf2來維持胞內的氧化還原穩態,從而在巨噬細胞對病原體清除期間保護細胞免受氧化損傷,延緩衰老與死亡.
細胞中的Nrf2蛋白主要受泛素-蛋白酶體系統嚴格調控:在正常生理條件下,多聚化的Keap1與Cul3-E3 泛素連接酶靶向Nrf2,促進Nrf2泛素化和蛋白酶體降解;而當細胞處于氧化應激條件下,Keap1發生解聚,Keap1-Cul3-E3 泛素連接酶無法穩定結合到Nrf2上,導致Nrf2泛素化水平下降,蛋白水平升高,進入細胞核中與ARE(antioxidant response element)序列結合以激活下游抗氧化應激酶的表達[56-57].在調控機制上,本研究團隊證實Mst1/2可直接結合Keap1,并對Keap1的4個位點進行磷酸化修飾,Mst1/2介導的Keap1磷酸化阻止了Keap1分子的多聚化,從而阻斷了Nrf2的泛素化和蛋白降解;進一步利用病毒載體回補Nrf2,可明顯降低Mst1/2缺失的巨噬細胞中ROS水平、DNA損傷和細胞凋亡比例,證實在巨噬細胞中Nrf2是Hippo激酶下游的關鍵分子[39].上述研究揭示在巨噬細胞參與宿主防御過程中存在一條新的抗氧化和抗衰老信號通路,闡明了Mst1/2參與調節巨噬細胞維持氧化應激穩態以實現清除病原體的同時,還能避免ROS誘導的自身損傷和細胞衰老的關鍵機制(圖3),為進一步研究氧化應激如何促進衰老相關炎癥和感染的發生和發展提供了重要的理論基礎.

在細菌感染條件下,巨噬細胞中的TLR聯合機械敏感受體Piezo1激活Mst1/2,活化Rac1,促進線粒體募集到吞噬體周圍產生殺傷性ROS,同時增強吞噬體細胞膜上的NADPH氧化酶活性而產生更多殺傷性ROS,最終殺滅細菌;大量ROS會進一步活化Mst1/2,通過磷酸化Keap1來調控Nrf2的穩定性,促進抗氧化基因表達,實現細胞的抗氧化應激反應.因此,Hippo信號通路在調控巨噬細胞的氧化還原穩態中發揮著重要作用.
4 總結與展望
Hippo信號通路在進化上高度保守,是調控細胞增殖和死亡、維持機體組織器官穩態的重要通路,一直被認為是重要的腫瘤抑制信號通路.近年來,本研究團隊和其他課題組的工作揭示了Hippo信號通路在免疫穩態調控中也發揮著重要功能.Mst1/2是Hippo信號通路的核心激酶,在巨噬細胞抗感染過程中,作為重要的分子開關協同調控兩種重要信號轉導,即誘導產生殺傷性ROS的信號和活化Nrf2以清除過量ROS的信號,以維持巨噬細胞的氧化還原穩態[29-30,39].這些結果揭示了天然免疫細胞在感染和不同外周環境刺激下,合適的免疫應答需要胞內多種細胞器和免疫效應分子在時間和空間上的協同作用.一些環節的調控缺陷會影響吞噬細胞對病原體的吞噬和殺傷,無法維持胞內的氧化還原穩態,導致細胞早衰和死亡.而在獲得性免疫調控方面,Hippo信號通路抑制促炎性Th17的分化,促進免疫抑制性Treg的分化,在正常生理條件下對于維持免疫系統耐受和穩態至關重要,闡釋了當Hippo信號通路失調時病人易發自身免疫性疾病的病理機制[38].這些研究表明非經典Hippo信號通路成員可能是治療感染性疾病、自身免疫性疾病、免疫細胞衰老以及腫瘤等免疫失調疾病的潛在靶點.
作為腫瘤抑制通路,靶向促進Hippo通路活性治療腫瘤的策略不能忽略Hippo信號通路活化對于Treg分化和功能的促進作用,因為Treg會抑制免疫系統對腫瘤細胞的清除;然而這種靶向治療卻可能會通過抑制Th17的分化和增強Treg的抗炎癥效應而有利于治療自身免疫性疾病.Hippo信號通路調控腫瘤發生發展以及組織再生和穩態,同時也調控免疫應答和功能,如果直接靶向Hippo通路的核心節點分子治療疾病,可能會帶來一些副作用,因此在設計小分子藥物時要考慮到作用位點、作用效率,以及作用是否可逆等因素,例如:本研究團隊發現的小分子藥物XMU-MP-1能夠可逆且特異性地抑制激酶Mst1/2的活性,因此可以通過精準的用藥窗口促進組織再生和修復而避免造成組織異常增生,已在肝臟和腸道損傷再生動物模型中取得了很好的治療效果[58].此外,只有深入研究Hippo信號通路在免疫系統中的精準調控功能,特別是該通路如何協同調控其他經典免疫信號通路,及其在調控腫瘤和免疫細胞互作中的作用,闡明在不同組織微環境和不同免疫應答事件中的具體效應分子,才能更好、更安全地將靶向Hippo信號通路作為治療腫瘤、損傷再生、感染和自身免疫病等相關疾病的治療策略.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年3期)2019-02-01 06:12:26
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
