稻米品質調控的分子基礎及非生物脅迫對稻米品質的影響
2022-05-26 16:43:20牛淑琳唐苗苗杜晨陽王增蘭謝先芝鄭崇珂
中國稻米 2022年3期
關鍵詞:水稻
牛淑琳 唐苗苗 杜晨陽 王增蘭 謝先芝* 鄭崇珂
(1山東師范大學 生命科學學院,濟南 250014;2山東省水稻研究所,濟南 250100;3山東農業大學 生命科學學院,山東 泰安271018;第一作者:nsl199811@163.com;*通訊作者:xzhxie2010@163.com;zhengck1983@163.com)
水稻是世界上重要的糧食作物。隨著人們生活水平的提高,優質稻米越來越受到消費者的喜愛,培育外觀優良,食味與營養兼具的稻米成為水稻研究中的重要課題[1-2]。現代生物技術和生物信息技術的發展,使一系列控制稻米優良品質的基因被發現并成功應用于優質水稻品種的選育。然而,頻發的極端天氣極大阻礙了優質水稻生產,對稻米的外觀品質、蒸煮食味品質與營養品質等均造成不良影響。本文在綜述調控稻米品質基因的基礎上對干旱、低溫、高溫、鹽脅迫對稻米品質的影響進行了概述,以期為改善非生物脅迫下的稻米品質提供指導。
1 稻米品質調控的分子基礎
稻米品質主要包括外觀品質、蒸煮食味品質、營養品質、加工品質和安全品質。精米是大米作為商品的主要形式,外觀是消費者重要的特征要求[3]。外觀品質主要包括水稻粒形、堊白粒率、堊白度和透明度等[4];香味、直鏈淀粉含量、膠稠度和糊化溫度等共同決定稻米的蒸煮食味品質[5];抗性淀粉含量、蛋白質含量、微量元素含量和其他營養物質含量等作為大米的主要營養成分決定了稻米的營養品質[6];加工品質又稱碾磨品質,主要通過糙米率、精米率、整精米率來進行評價[7];稻米安全品質主要通過稻米中農藥、重金屬元素(如砷、鎘、汞、鉛)、硝酸鹽等有害成分的殘留狀況來進行評價[8]。近年,調控稻米品質的多個基因被克隆(表1),其分子調控機制被解析并成功應用于優質水稻育種。
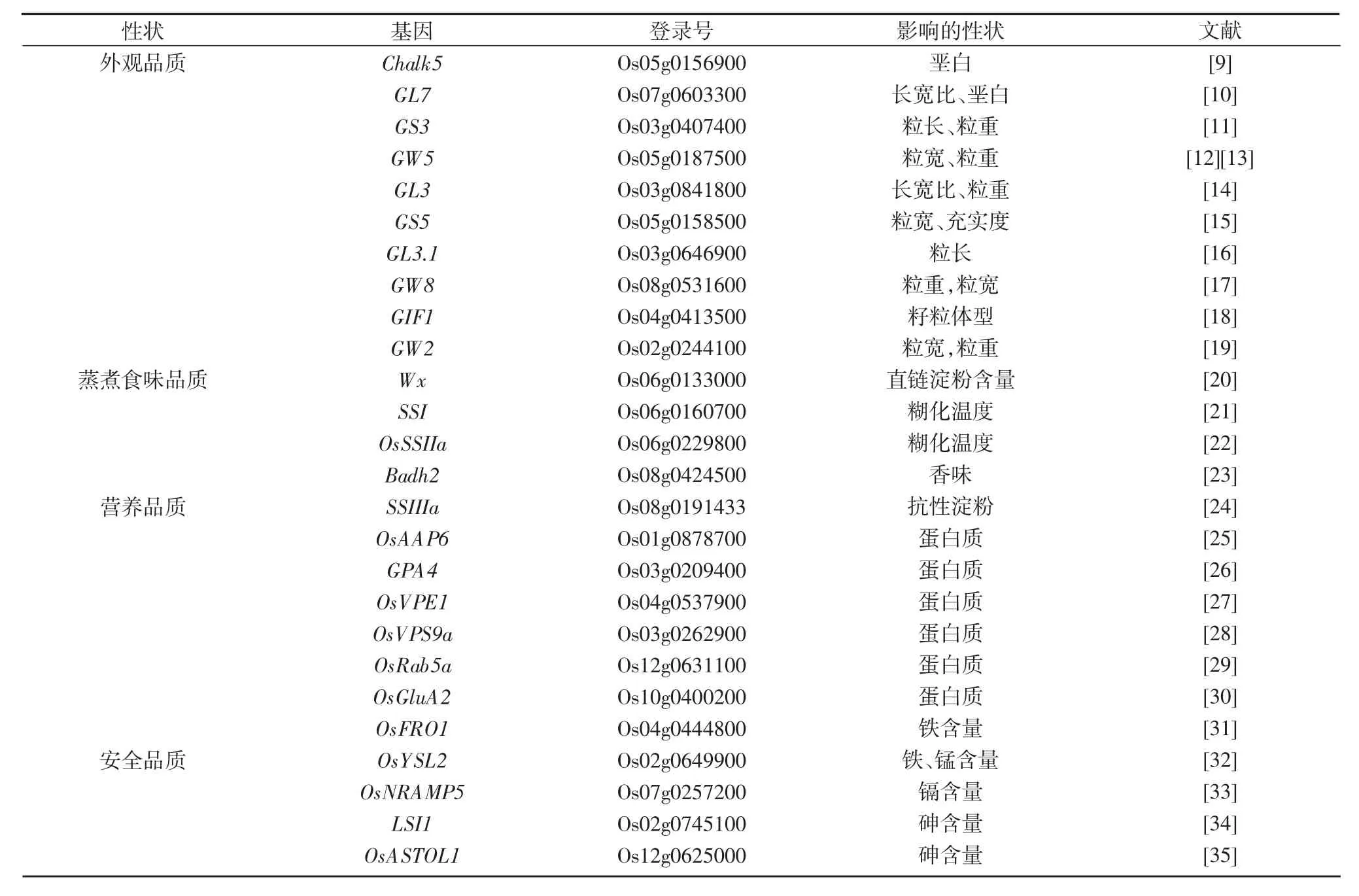
表1 水稻品質調控主要基因
1.1 外觀品質
優質的稻米一般要求外觀透亮,堊白面積少,堊白粒率低,長寬比合適[36]。堊白是種子胚乳內部灌漿不充實造成組織疏松而形成的不透明部分,會對碾磨、烹飪、食用和谷物外觀產生負面影響。LI等[9]利用秈稻H94(低堊白高品質)和珍汕97(高堊白低品質)雜交后代克隆了調控籽粒堊白的主效QTL,即Chalk5。Chalk5編碼1個液泡膜質子轉運焦磷酸酶,具有無機焦磷酸水解活性和質子轉運活性。Chalk5可能干擾了發育種子的內膜轉運系統的pH穩態,從而影響蛋白體的形成,造成囊泡類結構顯著增加,因此在胚乳儲存物質中形成了氣體空間,形成了籽粒的堊白。粒形相關基因在改良籽粒粒形的同時也會對籽粒的堊白性狀產生重要影響。秈稻地方品種Ping13(P13)具有較長的粒長和優異的外觀品質,在P13和日本晴(NPB)的F2群體中檢測到5個與粒長和/或粒寬相關的QTLs,其中1個位于第7染色體的QTL主要影響粒長,命名為Grain Length on Chromosome 7(GL7),GL7基因編碼擬南芥LONGIFOLIA蛋白的同源蛋白,提高GL7基因的表達不僅可以顯著增加谷粒長度,而且植株的堊白粒率和堊白度顯著降低,胚乳中的淀粉顆粒更大、更緊密[10]。
研究調控谷粒長和寬基因的分子機制以培育長寬比合適、外形優良的新品種,能更好的滿足當今消費者對于稻米外觀品質日益增長的需求。在眾多不同遺傳背景和環境的研究中,均在3號染色體的著絲粒附近檢測到1個影響谷粒大小的主效QTL,命名為Grain Size 3(GS3)。GS3編碼1個由232個氨基酸組成的跨膜蛋白,對粒質量起負調控作用[11]。GS3本身對籽粒大小無影響,但它與DEP1或GGC2競爭性結合Gβ,縮短粒長[37]。GW5(Grain Width and Weight on Chromosome 5)編碼一種由144個氨基酸組成的定位于細胞核的核蛋白,是水稻種子發育過程中通過調控細胞分裂來調控水稻籽粒寬度和質量的1個主效QTL,該基因的缺失可以增加籽粒的粒寬和粒質量[12]。TGW3為存在于第3染色體末端的控制粒長、粒質量的主效QTL,其在幼穗中表達,編碼1個類糖原合酶激酶GSK3(Glycogen Synthase Kinase 3)家族成員的激酶OsSK41/OsGSK5,定位于細胞核和細胞質中,能與生長素應答因子OsARF4(Auxin Response Factor 4)互作并將其磷酸化,OsARF4是生長素途徑中的轉錄抑制因子。Os-SK41-OsARF4可能通過負調控生長素信號,從而負調控水稻籽粒的大小和質量[14,38]。GS5(Grain Size on Chromosome 5)編碼1個絲氨酸羧肽酶,當其高表達時水稻籽粒變大,是1個控制水稻粒寬、充實度和千粒重的數量性狀基因[15,39]。GL3.1(Grain Length 3.1)編碼1個屬于蛋白磷酸酶PPKL家族的絲氨酸/蘇氨酸磷酸酶,能直接去磷酸化細胞周期蛋白T1;3,后者在水稻中的表達下調會使籽粒變短[16],同時GL3.1還可以抑制油菜素內酯信號的轉導進而使籽粒變短[40]。GW8(Grain Weight on Chromosome 8)編碼1個正調控細胞增殖的蛋白,該基因的高表達促進細胞分裂和籽粒灌漿,對水稻粒寬和產量有積極影響[17]。GIF1(Grf-Interacting Factor 1)編碼細胞壁蔗糖酶,蔗糖酶在把蔗糖轉化成淀粉的過程中起到重要作用,gif1突變體在形態和結實率上表現正常,但灌漿程度和粒質量比野生型低,籽粒淀粉粒排列松散,對籽粒良好體型造成不利影響[18]。GW2(Grain Width on Chromosome 2)編碼1個環型E3泛素連接酶,通過泛素蛋白酶體通路降解蛋白負調節細胞的分裂,GW2功能喪失的水稻穎殼變大,灌漿速率加快,粒寬和粒質量增加,產量提高[19]。
1.2 蒸煮食味品質
優質稻米除具有吸引力的外觀之外,更重要的是蒸煮之后的口感。目前,具有獨特香味的香稻越來越受到人們的喜愛,香味的產生主要歸功于2-乙酰基-1-吡咯啉。通過芳香型和非芳香型品種的雜交和回交獲得分離群體,在8號染色體精細定位到1個調控稻米香味的基因OsBADH2(Betaine Aldehyde Dehydrogenasea 2)[41]。OsBADH2編碼甜菜堿醛脫氫酶,具有醛脫氫酶活性,BADH2蛋白定位在細胞質中,除了在根中不表達外,在所有檢測的組織中包括葉、莖和穗部都有表達,其中在幼葉中表達量最高。BADH2蛋白催化2-乙酰基-1-吡咯啉的合成前體4-氨基丁醛的氧化,當4-氨基丁醛被BADH2氧化后,2-乙酰基-1-吡咯啉的合成受到抑制,因此稻米喪失了香味[23]。
飲食和烹飪質量主要受到直鏈淀粉含量(AC)、膠稠度(GC)和糊化溫度(GT)的影響。水稻Wx(Waxy)基因編碼顆粒淀粉合成酶(Granule-bound Starch Synthase,GBSS),是控制直鏈淀粉合成的主效基因,直接影響水稻胚乳和花粉中直鏈淀粉的含量。早在1990年,WANG等[42]以玉米蠟質基因DNA為探針從日本恒豐品種基因組文庫中篩選并鑒定了由14個外顯子和13個內含子組成的核苷酸序列,即Wx基因。通過關聯分析等手段對淀粉生物合成途徑中的18個基因進行系統研究[20],發現這些基因之間形成復雜的遺傳網絡來決定稻米蒸煮與食味品質,其中,Wx是唯一對AC和GC具有主效作用的基因,同時檢測到一些協同Wx調控稻米直鏈淀粉含量的微效基因。SS-III-2(Soluble Starch Synthase-III-2)、AGPlar[AGP(ADP-glucose Pyrophosphorylase)Large Subunit]、PUL(Pullulanase)和SSI(Soluble Starch synthase-I)4個基因只對AC有微效作用,AGPiso(AGPLarge Subunit Isoform)只對GC有微效影響;SSII-3(Soluble Starch Synthase-II-3)是唯一對GT具有主效作用、而對AC和GC具有微效性的基因。ISA(Isoamylase)和SBE3(Starch Branching Enzyme 3)同時對GC和GT有微效作用,而SS-IV-2(Soluble Starch Synthase-IV-2)只對GT有微效影響。同時還發現,淀粉合成相關基因在淀粉合成的不同階段對品質性狀產生影響并且形成一個精細調控網絡來影響稻米食用和蒸煮品質。由于Wx基因位點的重組率遠高于全基因組平均水平,因此產生了大量等位變異基因;除了Wx基因,目前已證明影響稻米食味品質的Wx等位基因有8個,分別為Wxlv、Wxa、Wxin、Wxb、Wxop/hp、Wxmq、Wxmp和Wxmw[43]。隨著對稻米淀粉合成途徑的了解,許多育種家利用CRISPR/Cas9技術和堿基編輯技術微調Wx基因表達進而適度調控AC以改良稻米品質[44-45]。也有研究表明,SSI蛋白表達的降低會通過改變支鏈淀粉的結構來降低稻米糊化溫度[21]。OsSSIIa(Starch SynthaseⅡa)編碼可溶性淀粉合成酶Ⅱ,當編碼產物中氨基酸發生改變會造成淀粉合成酶活性的變化,從而影響支鏈淀粉的中等長度分支鏈的合成,最終表現為糊化溫度的改變[22]。ALK(Alkali Degeneration)基因就是OsSSIIa基因,根據其編碼的酶是否有活性該基因存在2個主要的等位基因變異ALKa/ALKc[46],ALK基因變異主要影響糊化溫度,對直鏈淀粉含量無顯著影響,對膠稠度存在微效影響[47]。OsAAP6(Amino Acid Permease 6)編碼氨基酸通透酶,不僅能增加稻米中蛋白質和直鏈淀粉的含量,而且可以顯著降低膠稠度[25]。
1.3 營養品質
稻米營養品質指稻米中各種營養物質的含量,主要包括抗性淀粉含量、蛋白質含量、微量元素含量和其他營養物質含量[36]。隨著對不同代謝物在稻米中的生物合成途徑和對人類健康影響的認識不斷增加,營養價值的概念已經得到了廣泛的拓展。例如,抗性淀粉(RS)有助于預防Ⅱ型糖尿病和肥胖癥,同時還具有降低腹瀉、炎癥性腸病、結腸癌和慢性肝腎疾病發病率的潛力。ZHOU等[24]從經γ射線輻射過的雜交水稻恢復系R7954群體中篩選出了1個在煮熟的大米中具有高RS的突變體,并通過圖位克隆等方法克隆了編碼可溶性淀粉合成酶的SSIIIa(Soluble Starch Synthase IIIa)基因。該基因功能的缺失使得直鏈淀粉和脂類的合成增加從而累積抗性淀粉,而且SSIIIa/SSIIa雙抑制株系的籽粒呈現堊白外觀,直鏈淀粉含量增加,糊化溫度升高,黏度降低[48]。OsAAP6能夠促進水稻根對氨基酸的吸收和轉運,增加體內氨基酸的含量,使籽粒中的谷蛋白、醇溶蛋白、球蛋白和清蛋白等增加;更重要的是OsAAP6控制蛋白質含量,但對植物形態、開花時間和籽粒產量沒有影響[25]。OsGluA2編碼谷蛋白A2型前體,能夠通過增加籽粒存儲蛋白含量和氨基酸總量正調控水稻籽粒的蛋白含量,提高稻米的營養品質[30]。谷蛋白是水稻主要貯藏蛋白,約占稻米蛋白質含量的60%[49],易被人體消化吸收,改變胚乳中谷蛋白含量已成為當今改善稻米營養品質的一大趨勢。GPA4(Glutelin Precursor Accumulation 4)將編碼谷蛋白的RNA定位到皮層內質網,并將谷蛋白前體由內質網運輸進高爾基體[26];采用抑制消減雜交(SSH)[50]和篩選突變體[51]的方法均鑒定到1個編碼小分子GTP酶的基因OsRab5a(Small GTP-binding Protein),Rab5a是高爾基體中谷蛋白前體向蛋白質儲存泡(PSV)細胞內輸送所必需的[29];OsVPS9A(Vacuolar Sorting Protein 9)編碼1個鳥苷酸交換因子(GEF),GEF是Rab5a GTP酶的催化劑,OsVPS9A和OsRab5A協作共同發揮協助谷蛋白前體在體內運輸的作用[28]。OsVPE1(Vacuolar Processing Enzyme1)編碼1個液泡加工酶,將谷蛋白前體加工成谷蛋白晶體結構并儲存于PSV以快速提供幼苗生長階段對氨基酸源的需求[27]。谷蛋白合成及運輸途徑分子機制的研究,對于培育適宜于不同消費群體的高/低谷蛋白含量稻米具有重要意義,例如,腎病患者更偏愛成本較低的低蛋白稻米來代替其他低蛋白用品進行食療[52]。OsFRO1(Ferric Chelate Reductase1)編碼能將Fe3+還原為Fe2+的鐵還原酶氧化酶,在水稻獲取鐵的過程中發揮著重要作用[53],突變體fro1不僅在營養階段對鐵耐受,而且其籽粒的鐵含量增加21%~30%[31]。OsYSL2(Yellow Stripe-like 2)編碼1個水稻金屬-尼克酰胺轉運蛋白,在韌皮部細胞和發育中的種子中表達,是Fe和Mn長距離運輸所必需的,包括將其運輸到籽粒中[32]。Zn是水稻生長發育所必需的營養元素之一,能促進水稻生長,改善稻米品質,目前在第1、3、5、6、7、8、11和12號染色體上均已定位到控制水稻籽粒Zn含量的QTLs[54],但對于這些基因的功能與相關遺傳分析仍缺乏研究。稻米中富集有生理效應的功能性因子Se,具有保健、防病、優質和營養等多種特點,而對于水稻籽粒含Se量的QTL定位與遺傳機制鮮有報道。孫明茂[55]在第1、2、7和10號染色體檢測到7個與Se含量相關的QTLs。張現偉等[56]在第5號染色體檢測到3個與Se含量相關的QTLs。受籽粒Zn、Se等營養元素含量檢測方法的制約,不僅檢測出的調控籽粒養分元素含量的QTLs較少,而且在分子標記輔助育種中的應用也受到制約。
1.4 安全品質與加工品質
目前稻米安全品質越來越受到公眾關注,但這方面的研究還很欠缺,尚未有關于調控稻米中農藥殘留、硝酸鹽含量基因的報道。水稻籽實中重金屬主要來源于土壤、灌溉水、大氣、收割、翻曬和加工設備等,其中起主要作用的是土壤和灌溉水[57]。OsNRAMP5(Natural Resistance-associated Macrophage Protein)編碼1個自然抗性相關的巨噬細胞蛋白。ISHIKAWA等[33]通過碳離子束輻射獲得了osnramp5突變體,其無法編碼具有正常功能的轉運蛋白,因此極大降低了根部對鎘(Cd)的吸收,從而降低了籽粒中的Cd含量。當種植在被Cd污染的水田中,突變體籽粒中Cd的含量接近檢出下限,也沒有明顯的不利農藝和經濟性狀。Lsi1(Low Silicon Rice 1)編碼1個硅轉運蛋白,研究表明,水稻吸收和轉運亞砷酸鹽也通過此蛋白,lsi1突變體的根部和地上部積累的亞砷酸鹽減少,而谷粒中的亞砷酸鹽含量沒有變化。通過對突變體lsi2的研究發現,砷在水稻谷粒中的積累增加[34,58]。OsASTOL1(O-Acetylserine(thiol)Lyase 1)編碼1個半胱氨酸合酶,可以正調控S和Se的吸收和同化,提高水稻體內S和Se含量,可以使稻米呈現降砷富硒表型[35]。唐紹清[59]通過種植于重金屬污染嚴重稻田里的367份不同材料共檢測到10個控制重金屬Cu、Cd和Pb含量的QTLs,3個影響Cu含量的QTLs位于第3、9和12號染色體,3個影響Cd含量的QTLs位于第3、7和11號染色體,4個影響Pb含量的QTLs位于第4、5和12(2個)號染色體。
目前尚未有關于精米率、整精米率等調控加工品質的相關基因的報道。但隨著分子生物學技術的發展,會有越來越多的調控稻米品質的基因被克隆,稻米品質形成的分子網絡機制將不斷完善,為育種家進行稻米品質改良提供了豐富的基因基礎。同時利用已經解析的稻米品質調控機制,培育了眾多優質的水稻新品種,豐富人們的餐桌,提高人們的生活水平。
2 非生物脅迫對稻米品質的影響
近年來,非生物脅迫頻發,干旱、低溫、高溫、鹽脅迫等不僅降低了水稻產量,也影響了稻米品質,具體影響如圖1所示。提高多變氣候下、鹽漬化土壤上的水稻品質對于民生意義重大。不同的非生物脅迫對于稻米品質的影響不盡相同,通過了解不同非生物脅迫對于稻米品質的具體影響從而作出防御措施對減少資源浪費,培育耐非生物脅迫的優質水稻品種具有重要意義。

圖1 干旱、高溫、低溫和鹽脅迫對稻米品質的影響
2.1 干旱對稻米品質的影響
水稻是耗水量極大的農作物,干旱脅迫嚴重影響水稻的產量和品質。研究表明,對水稻灌漿期進行干旱脅迫處理后,胚乳中Wx基因的相對表達量減小,并且其表達模式與直鏈淀粉積累模式基本一致,造成籽粒中直鏈淀粉含量的降低,表明稻米食味品質的變化受基因表達量影響[60]。王成璦等[61]以農大3號(粳稻)為材料研究發現,不同生育階段的干旱脅迫對加工品質的影響存在顯著差異:灌漿期至蠟熟期干旱脅迫糙米率和整精米率都有所下降,其中對整精米率的影響主要表現在抽穗后11~40 d;孕穗中期和孕穗后期干旱脅迫也會使糙米率、整精米率明顯下降。而對揚兩優6號和漢優113開花期干旱脅迫除顯著提高堊白粒率和堊白度外,對外觀和營養品質均無顯著影響[62]。HAIDER等[63]通過對20個水稻品種進行干旱脅迫后發現,參試品種的粒長和粒寬均因干旱而減少,粒厚均略有增加。張家亮[64]以川秈955和9311為材料,發現干旱脅迫后GS3基因在幼穗分化第11天急劇升高,第18天時表達水平有所下降但仍高于對照,推測干旱脅迫下谷粒的縮小是由于對谷粒大小起負調作用的GS3的過表達引起。除此之外,水分脅迫下Mn、Fe、Cu等元素含量與正常培養條件下相比也有所下降[65]。王平榮等[66]對13個雜交水稻組合及恢復系自抽穗前7 d持續干旱處理,結果表明,干旱降低了稻米的加工品質,糙米率、精米率和整精米率都比未處理的對照降低,其中整精米率降低最明顯;干旱對稻米的外觀品質也有影響,主要表現在堊白粒率和堊白度顯著增加,而對于粒型的影響并不明顯;稻米的蒸煮和食味品質也受到了干旱的影響,如糊化溫度提高、膠稠度變硬、直鏈淀粉含量降低等。李國生等[67]以汕優63(雜交秈稻)和武育粳3號(常規粳稻)為材料,自抽穗至成熟期進行不同程度的干旱處理,結果表明,當汕優63結實期土壤水勢為-15kPa和-30kPa時,武育粳3號土壤水勢為-15kPa時,有利于改善稻米的外觀品質和食味品質,如堊白粒率和堊白度降低,最高粘度和崩解值增加,消減值降低等。段驊[68]對黃華占、雙桂1號、兩優培九和揚粳4038于抽穗和灌漿早期進行干旱處理后,發現兩優培九SSSI基因顯著下調,而其他品種則表現為顯著上調。不同時期的干旱脅迫對不同水稻品種品質產生的影響不盡相同,目前對于造成這種結果的分子機制尚不清楚,探究稻米品質下降的深層次原因至關重要。
2.2 低溫對稻米品質的影響
低溫脅迫是影響植物生長的嚴重環境脅迫之一,雖然耐冷品種的出現解決了水稻低溫脅迫下的產量問題,然而低溫對于稻米品質的影響正日益受到育種家的關注。任紅茹[69]對4個水稻品種在孕穗期進行不同低溫處理,發現低溫降低了加工品質,糙米率、精米率和整精米率均不同程度下降,其中整精米率的降幅最大;對外觀和營養品質的影響表現為降低了籽粒的粒長、粒寬,降低了籽粒的直鏈淀粉含量,增加了蛋白質含量。而王士強等[70]研究發現,孕穗期低溫處理后稻米直鏈淀粉含量增加。褚春燕等[71]發現,低溫脅迫后直鏈淀粉含量在孕穗期和抽穗期顯著降低,灌漿期逐漸增加。武琦[72]對寒地粳稻灌漿期低溫脅迫后發現,淀粉合成過程關鍵酶ADPG焦磷酸羧化酶和籽粒淀粉分支酶活性降低,造成籽粒總淀粉含量下降。SIDDIK[73]對秈稻萬象優華占和榮優華占低溫脅迫后發現,抽穗后第2周籽粒中Wx基因表達量升高,顆粒結合淀粉合成酶-1(GBSS1)活性降低。低溫脅迫后直鏈淀粉含量的變化情況目前尚無定論,這可能與直鏈淀粉含量性狀的復雜性有關,還有待進一步深入研究。堊白的產生與不同時期遭受低溫脅迫有關:孕穗期低溫脅迫下堊白粒率和堊白度下降[69],拔節期低溫脅迫下堊白粒率顯著減少,抽穗期低溫脅迫下堊白粒率顯著增加[74],灌漿前期低溫脅迫下堊白度、堊白粒率降低[75]。所以在特定的時期適當低溫脅迫有助于提高稻米的外觀品質。
2.3 高溫對稻米品質的影響
由于全球變暖,稻米受到高溫脅迫的可能性增加,在營養生長期和生殖生長期受到高溫脅迫會導致秧苗死亡[76]和敗育[77],從而降低產量。而在灌漿結實期高溫脅迫會導致稻米品質下降[78]。對南粳46和嘉58在灌漿期高溫處理,結果發現大米的食用和烹飪質量與正常溫度生長的大米相比明顯下降,同時在高溫條件下,稻米中長鏈直鏈淀粉和短鏈支鏈淀粉明顯減少,結晶性和糊化性能均高于正常條件種植的稻米[79]。CHUN等[80]對水稻Ilpum和Chucheong灌漿期高溫處理,發現直鏈淀粉含量和短枝鏈數量減少,而中間枝鏈數量增加,導致淀粉的糊化溫度和糊化焓升高。這些結果表明,在較高溫度下種植水稻需要增加蒸煮溫度和時間。在28℃/20℃(晝/夜)以上生長的水稻顯示出蒸煮質量的下降,在感官測試中有降低口感的趨勢。在稻米品質的各項指標中,受高溫條件影響最為顯著的方面為堊白性狀和RVA譜,其他指標變化較為遲鈍[81]。滕中華等[82]對西農優30和富優1號灌漿結實期進行高溫脅迫,發現成熟稻米總淀粉及直鏈淀粉含量降低,蛋白質含量升高,堊白度上升。黃福燈[83]的研究也有同樣的結果。張桂蓮等[84]對耐熱品種996和熱敏感品種4628在抽穗開花期進行高溫處理,結果表明高溫顯著降低了稻米的精米率、整精米率、堊白粒率和堊白度,且耐熱品種降幅小于熱敏感品種;另外,高溫下稻米堿消值、膠稠度和直鏈淀粉含量也有所下降,但蛋白質含量提高,推測是因為高溫促進了氨基酸向籽粒的運輸[85]。LIN等[86]對秈稻臺中在來1號和粳稻臺農67灌漿期高溫處理,發現在灌漿前期高溫(白天35℃/夜間30℃)增強了谷蛋白、脯氨酸、球蛋白和蛋白二硫異構酶基因的轉錄,但在灌漿后期降低了這些基因的表達。大米的硬度主要是由蛋白質含量決定,硬度越大,蛋白質含量越高,透明度也越高,但高溫脅迫下淀粉顆粒排列松散形成的堊白抵消了透明度高這一優點[87]。謝曉金等[88]研究發現,抽穗期高溫脅迫降低了揚稻6號與南粳43稻米中可溶性糖和蛋白質含量,而堊白度依舊大大上升。稻米在灌漿期受高溫脅迫后對堊白這一品質影響最大[89]。堊白的產生主要與淀粉合成途徑有關,在灌漿期進行高溫處理(白天33℃/夜間28℃),發現與淀粉合成相關的基因GBSS1表達明顯下調,因此導致直鏈淀粉含量下降,堊白度增加[90]。查曼[91]對秈稻3611和粳稻2661背景下的2個近等基因系材料在灌漿結實期進行高溫處理,發現秈粳背景下Wxb基因表達水平均受高溫誘導下調,Wxa基因總mRNA表達量不受高溫影響,但Wxa基因成熟mRNA受高溫誘導的表達趨勢完全相反,表現為3611背景上調表達,2 661背景下調表達。SIDDIK[73]對秈稻萬象優華占和榮優華占高溫脅迫后發現,抽穗后第2周籽粒中Wx基因相對表達量下降,OsAAP6基因相對表達量升高。曹珍珍[92]的研究結果表明,高溫脅迫加速了GluA2、GluA3和GluBl等谷蛋白亞基基因在水稻灌漿前中期的表達,抑制了淀粉合成類基因GBSS1和SSIIa的表達。段驊[68]對黃華占、雙桂1號、兩優培九和揚粳4038于抽穗和灌漿早期進行高溫處理,發現揚粳4038 SSSI基因表達顯著上調,其余品種顯著下調。
2.4 鹽脅迫對稻米品質的影響
隨著耐鹽水稻的培育,土壤鹽漬化這一困擾全世界的耕地資源浪費問題有望解決,然而鹽脅迫對于稻米品質的影響同樣不可小覷。羅成科等[93]對吉粳105研究發現,低濃度鹽分(0.1%)有利于提高水稻加工品質,略增加直鏈淀粉含量和蛋白質含量;而高濃度鹽分(0.2%~0.4%)易引起品質變劣,籽粒堊白明顯增加。余為仆[94]通過對揚稻6號和淮稻13研究得到類似結論。也有研究表明,在8 dS/m或更高的鹽水平下直鏈淀粉含量下降,然而2 dS/m和4 dS/m鹽水平下直鏈淀粉含量增加[95-96]。而肖丹丹等[97]以南粳9108和鹽稻12號為材料研究發現,低鹽濃度(0.10%~0.15%)下稻米的碾磨品質提高,米飯的外觀、黏度、平衡度和食味值總體上有所增加,稻米的膠稠度也有所提高。馬凌霄等[98]對水稻在不同濃度(0.15%、0.20%、0.25%、0.30%、0.35%、0.40%)鹽脅迫下的品質變化進行研究,結果表明,鹽分濃度與糙米率、精白度、粒型(長寬比)呈顯著負相關;而0.25%、0.30%鹽分濃度有提高水稻蒸煮品質的作用。翟彩嬌等[99]以常農粳8號和南粳9108為材料研究發現,隨著鹽脅迫強度的增加,食味值逐漸下降;而當土壤鹽濃度達到0.6%時,食味值與對照組相比顯著增加。SALEETHONG等[100]通過測量耐鹽品種Pokkali和鹽敏感品種KDML105在鹽脅迫下的籽粒營養元素,發現鹽脅迫導致兩種水稻籽粒N、P、K、Mg顯著減少,Na、Cu上升;鹽對Fe和Zn的影響不同,即在KDML105中增加,在Pokkali中下降。VERMA等[101]發現,隨著鹽度的增加,IR10198-66-2和IR28籽粒中Na、Fe、Zn含量增加,P、Mn含量降低,而N、Mg、Cu、K、Ca含量不受鹽脅迫的影響。陳景陽[102]對耐鹽型紅米HH11、HH12、JX99和白米YSXD、XZ3、TXZ共6個材料鹽脅迫后測量微量元素與營養因子變化,發現低鹽脅迫后紅米的花青素含量、總黃酮含量、色氨酸和賴氨酸含量均高于對照;紅米籽粒中可溶性糖積累較少,可能其植株大部分可溶性糖用于滲透壓調節。
3 展望
水稻品質是受多基因控制的復雜農藝性狀,系統解析決定品質的遺傳性狀十分具有挑戰性。雖然眾多調控稻米品質的基因已被定位與克隆,然而基因間往往牽一發而動全身,基因間是如何相互關聯影響稻米品質的?在遭受非生物脅迫后基因又是如何調節的?我們將如何利用已鑒定的基因提高非生物脅迫下稻米的品質?國內外有很多關于非生物脅迫影響稻米品質的報道,但是非生物脅迫影響稻米品質的分子機制仍不很清楚。這些都是目前亟待解決的問題。了解非生物脅迫下對于稻米品質具體某一方面或者某幾方面的影響從而進行定制的分子設計對于育種意義非凡,將分子與生理研究齊頭并進有機結合,相信必將產量與品質齊收,為農業注入源源不斷的活力。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
