渤海灣浮游纖毛蟲豐度和生物量的周年變化
2022-05-26 10:03:04張博倫徐曉甫張武昌
生態學報 2022年9期
于 瑩,王 宇,張博倫,王 碩,徐曉甫,張武昌
1 泰山學院,泰安 271000 2 天津市水產研究所,天津 300457 3 中國科學院海洋生態與環境科學重點實驗室,中國科學院海洋研究所,青島 266071 4 青島海洋科學與技術(試點)國家實驗室,海洋生態與環境科學功能實驗室,青島 266237 5 中國科學院海洋大科學研究中心,青島 266071
作為海洋微食物網的重要組成類群[1—2],浮游纖毛蟲粒徑較小(5—200 μm),主要包括營浮游生活的無殼寡毛類纖毛蟲和砂殼纖毛蟲兩大類,分別隸屬于纖毛門(Ciliophora Doflein),旋毛綱(Spirotrichea Bütschli)下的寡毛亞綱(Oligotrichia Bütschli)及環毛亞綱(Choreotrichia Small &Lynn)[3]。
溫帶近岸海區的浮游纖毛蟲豐度和生物量呈現明顯的季節波動,一年之中往往呈現雙峰值,一般在春季和夏季[4]或春季和秋季[5]達到高峰。無殼纖毛蟲通常在浮游纖毛蟲中占絕對優勢,砂殼纖毛蟲占浮游纖毛蟲豐度的比例一般<10%[6—7],砂殼纖毛蟲偶爾在溫度較高的夏季或秋季超過無殼纖毛蟲豐度占優勢[8—10]。
渤海灣位于渤海西部,面積約為1.47×104km2,平均水深12.5 m,海水交換能力弱,是一個典型的半封閉性淺水海灣。于瑩等[11—12]對渤海灣浮游纖毛蟲豐度和種類的季節變化有過調查,但沒有完整的周年變化資料。本研究于2019年7月—2020年6月在渤海灣天津近岸海域進行浮游纖毛蟲豐度和生物量的周年調查,為渤海灣浮游生態系統研究提供基礎數據和參考資料。
1 材料和方法
2019年7月—2020年6月在渤海灣固定站位(117°59′E,39°02′N,水深9—12 m,圖1)進行每月1次(2019年7月22日、8月23日、9月23日、10月21日、11月25日、12月23日、2020年1月16日、2月24日、3月20日、4月19日、5月25日及6月22日)共12個航次纖毛蟲樣品的采集。
圖1 采樣站位圖Fig.1 Location of the sampling station
用YSI Professional Plus便攜式水質分析儀測定表層海水的溫度和鹽度。采集表層水樣500 mL,用GF/F濾膜過濾,濾膜置于-20℃冰箱保存。濾膜帶回實驗室用90%丙酮低溫(0 ℃)萃取24 h后,用分光光度計測量葉綠素a(Chl a)濃度。
采集表層水樣1 L,用Lugol′s液固定(終濃度1%)。樣品分析按照Uterm?hl方法[13]進行,自然沉淀至少48 h,虹吸后剩余約150 mL,于陰涼處保存。取16 mL放于沉降杯內,在Olympus IX71倒置顯微鏡下100倍或400倍鏡檢,記錄無殼纖毛蟲和砂殼纖毛蟲(不計空殼)的個數。測量纖毛蟲蟲體的體長、體寬等,按最接近的幾何形狀(如柱體、球體和錐體)計算體積。生物量由體積乘轉換系數(0.19 pg C/μm3)[14]得來,砂殼纖毛蟲體積直接按照肉體體積計算。
按照纖毛蟲細胞體積的大小將無殼纖毛蟲分成3種粒級:小型無殼纖毛蟲(ACI,103—104μm3)、中型無殼纖毛蟲(ACII,104—105μm3)及大型無殼纖毛蟲(ACIII,>105μm3)[15]。砂殼纖毛蟲根據文獻[16]鑒定到種,按照砂殼纖毛蟲種類出現的季節規律,將砂殼纖毛蟲分為周年出現種、冬季種、春季種、夏季種及秋季種五大類[17]。
根據徐兆禮等[18]的公式計算優勢度(Y),Y>0.02時,該種為優勢種。砂殼纖毛蟲群落的多樣性采用Shannon指數(H′)[19],均勻度采用Pielou指數(J)[20]。
采用R語言的vegan程序包進行砂殼纖毛蟲群落結構和環境因子的Mantel檢驗,經過9999次排列得到統計結果,置信度設為95%;采用SPSS 20.0進行纖毛蟲豐度和生物量與環境因子的相關性分析,運用Pearson相關分析方法,置信度設為95%。
2 結果
2.1 環境因子
表層海水溫度、鹽度和Chl a濃度呈現明顯的周年變化。年平均溫度為(14.15±9.79)℃(平均值±標準差,下同),1月最低(2.35 ℃),7月最高(29.60 ℃);年平均鹽度為31.53±0.44,夏秋季較低,其中7月最低(30.76);年平均Chl a濃度為(3.67±4.19)μg/L,一年之中出現兩個峰值,分別在3月(5.53 μg/L)和7月(15.29 μg/L,圖2)。
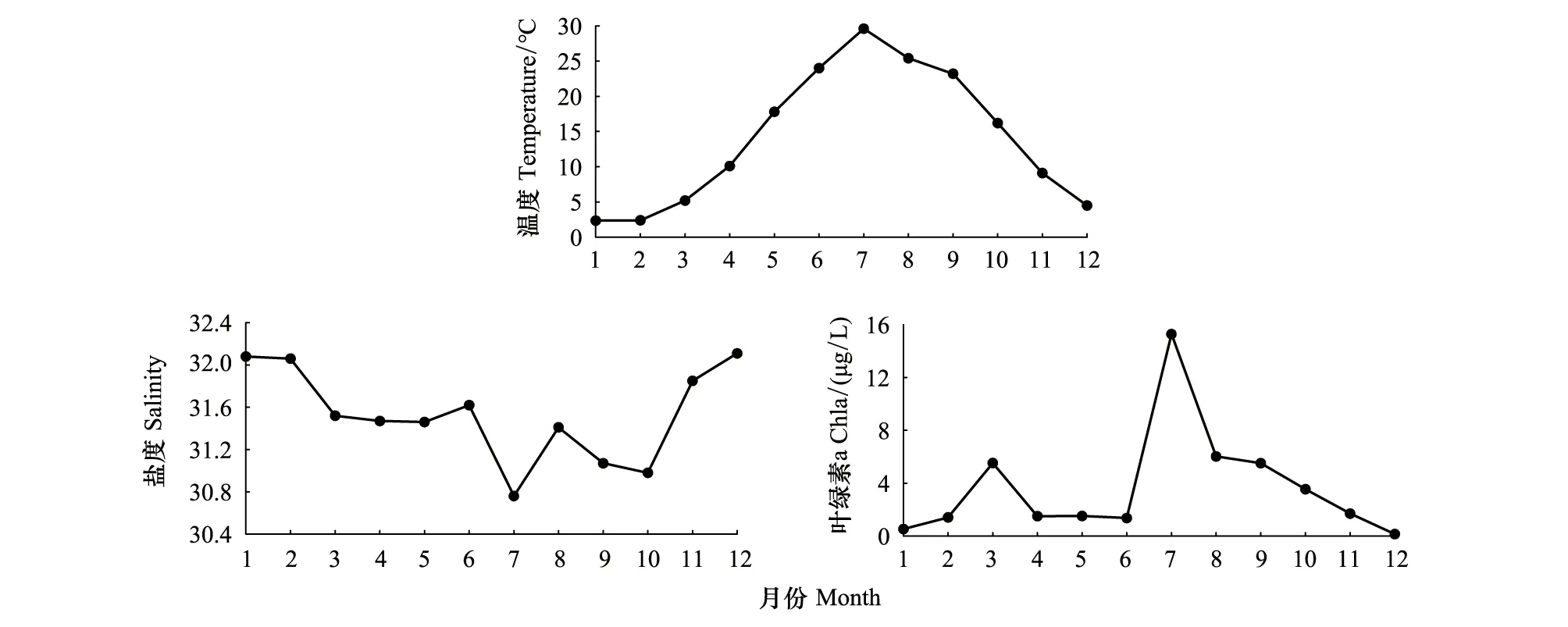
圖2 溫度、鹽度及Chl a濃度的周年變化Fig.2 Annual variation of water temperature,salinity and Chl a concentration
2.2 纖毛蟲豐度和生物量
無殼纖毛蟲年平均豐度為(1382±1929)個/L(137—6748 個/L),年平均生物量為(2.29±3.05)μg C/L(0.09—10.83 μg C/L),豐度和生物量均在4月和8月呈現雙峰值;砂殼纖毛蟲年平均豐度為(946±2384)個/L(0—8440 個/L),年平均生物量為(3.86±11.57)μg C/L(0.00—40.57 μg C/L),豐度和生物量均在7月出現最高值;總纖毛蟲年平均豐度為(2328±2735)個/L(143—8577 個/L),年平均生物量為(6.15±11.35)μg C/L(0.09—41.02 μg C/L),豐度和生物量均在4月和7月呈現雙峰值(圖3)。
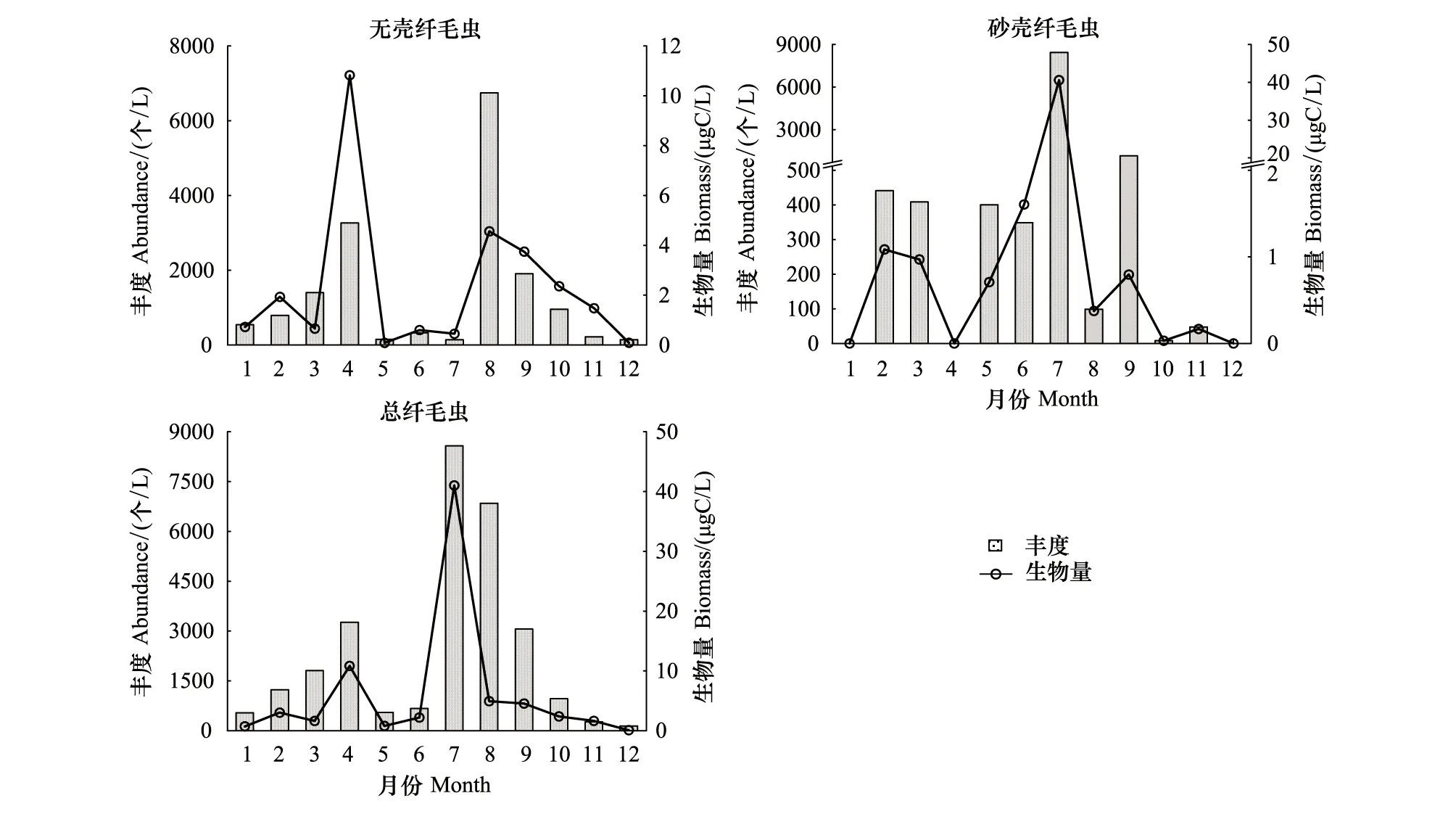
圖3 無殼纖毛蟲、砂殼纖毛蟲及總纖毛蟲豐度和生物量的周年變化Fig.3 Annual variation of abundance and biomass of aloricate ciliates,tintinnids and total ciliates
砂殼纖毛蟲占纖毛蟲總豐度的比例平均為(28.6±32.6)%,5—7月較高,占比均超過50%,其中7月最高(98.4%)。
2.3 無殼纖毛蟲粒級結構
無殼纖毛蟲粒級結構沒有呈現明顯的季節規律。ACI在無殼纖毛蟲豐度中占絕對優勢,占無殼纖毛蟲豐度的比例平均為(76.7±25.1)%;ACII占無殼纖毛蟲豐度的比例平均為(20.6±25.2)%;ACIII占無殼纖毛蟲豐度的比例最小,平均值為(2.7±4.9)%(圖4)。
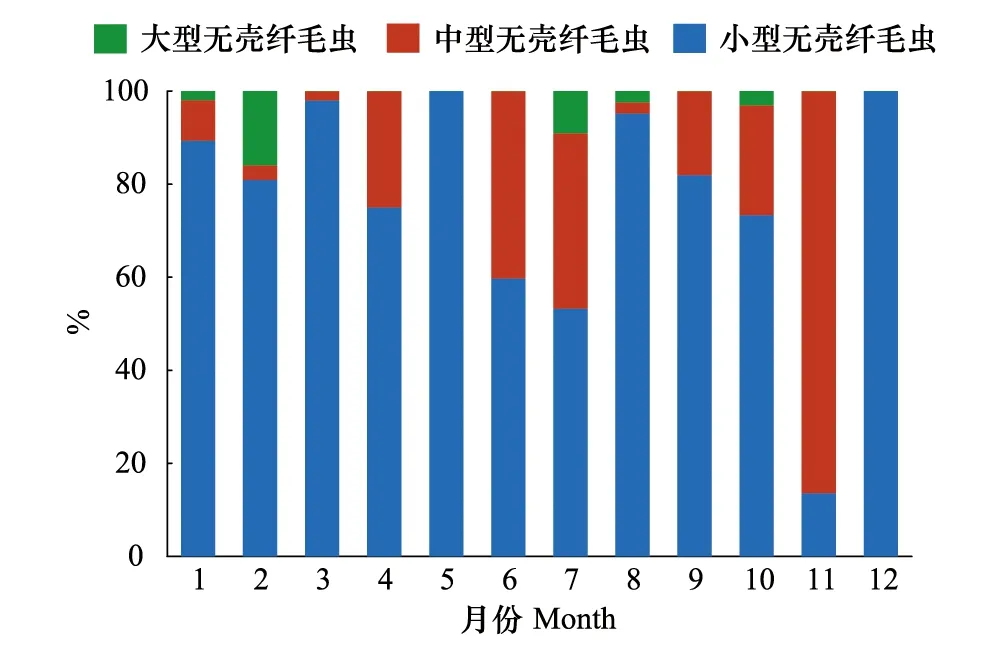
圖4 無殼纖毛蟲粒級組成的周年變化Fig.4 Annual variation of size classes of aloricate ciliates
2.4 砂殼纖毛蟲群落
2.4.1種類組成
共鑒定砂殼纖毛蟲6屬21種(表1),其中擬鈴蟲屬(Tintinnopsis)種類最多(13種)。優勢種為簡單薄鈴蟲(Leprotintinnussimplex,Y=0.08)、諾氏薄鈴蟲(Leprotintinnusnordqvisti,Y=0.04)和原始筒殼蟲(Tintinnidiumprimitivum,Y=0.03)。砂殼纖毛蟲種類數在6—8月較高,7月最高(10種)。
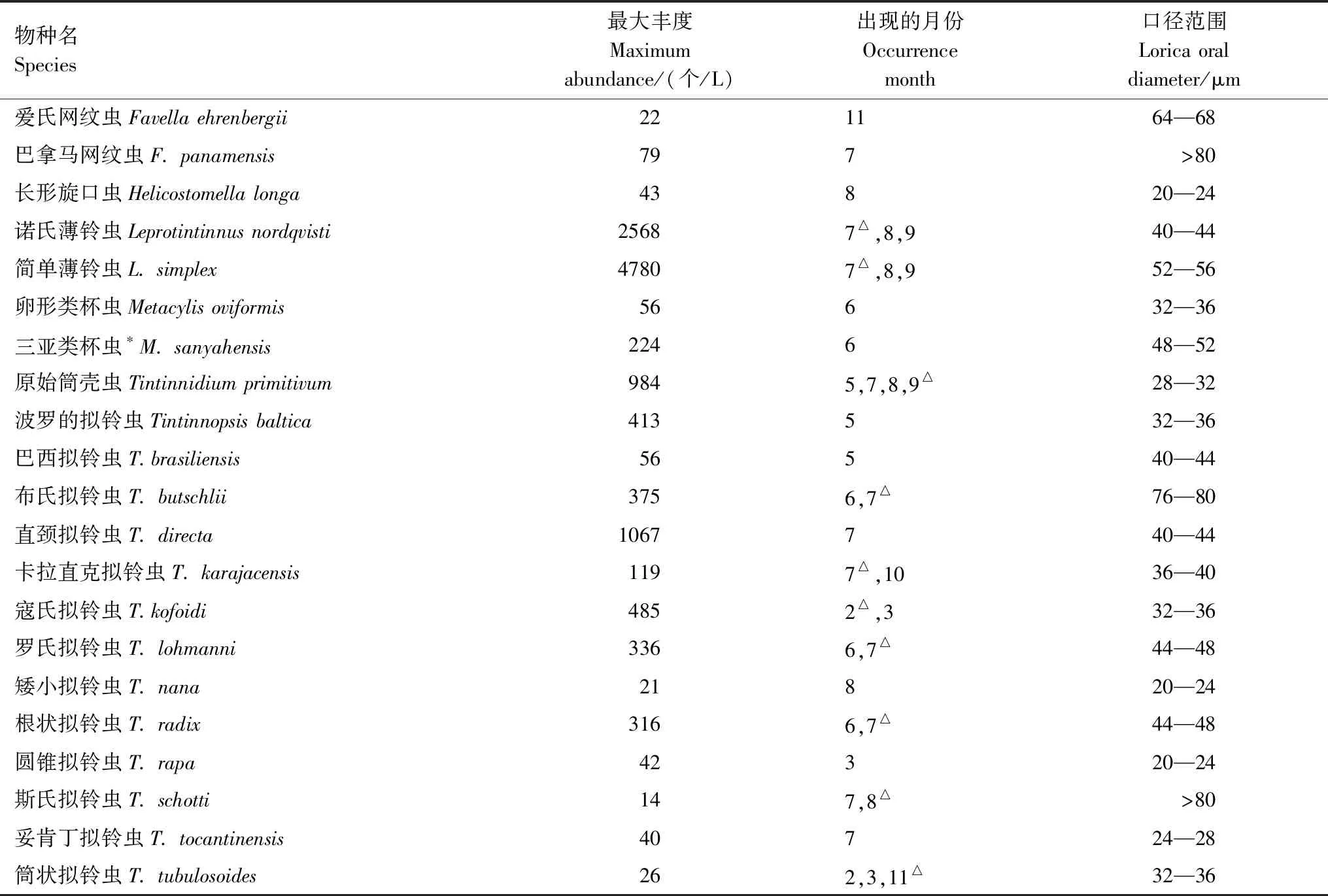
表1 渤海灣砂殼纖毛蟲種類Table 1 Species list of tintinnids in the Bohai Bay
豐度較高(豐度占砂殼纖毛蟲總豐度的比例>2%)的種類有10種,為簡單薄鈴蟲(36.9%)、諾氏薄鈴蟲(19.5%)、原始筒殼蟲(11.0%)、直頸擬鈴蟲(Tintinnopsisdirecta,8.2%)、寇氏擬鈴蟲(Tintinnopsiskofoidi,7.0%)、波羅的擬鈴蟲(Tintinnopsisbaltica,3.2%)、羅氏擬鈴蟲(Tintinnopsislohmanni,2.8%)、根狀擬鈴蟲(Tintinnopsisradix,2.8%)、布氏擬鈴蟲(Tintinnopsisbutschlii,2.6%)及三亞類杯蟲(Metacylissanyahensis,2.0%)。其中,寇氏擬鈴蟲只在溫度較低(2.4—5.2℃)的冬季出現,是冬季種。波羅的擬鈴蟲只在溫度為17.8℃的春季出現,是春季種。原始筒殼蟲在春季和夏季均有發現,在夏季(>23.2℃)豐度較高;三亞類杯蟲、布氏擬鈴蟲、羅氏擬鈴蟲、根狀擬鈴蟲、諾氏薄鈴蟲、簡單薄鈴蟲及直頸擬鈴蟲只在溫度較高(>23.2℃)的夏季出現,是夏季種(圖5)。
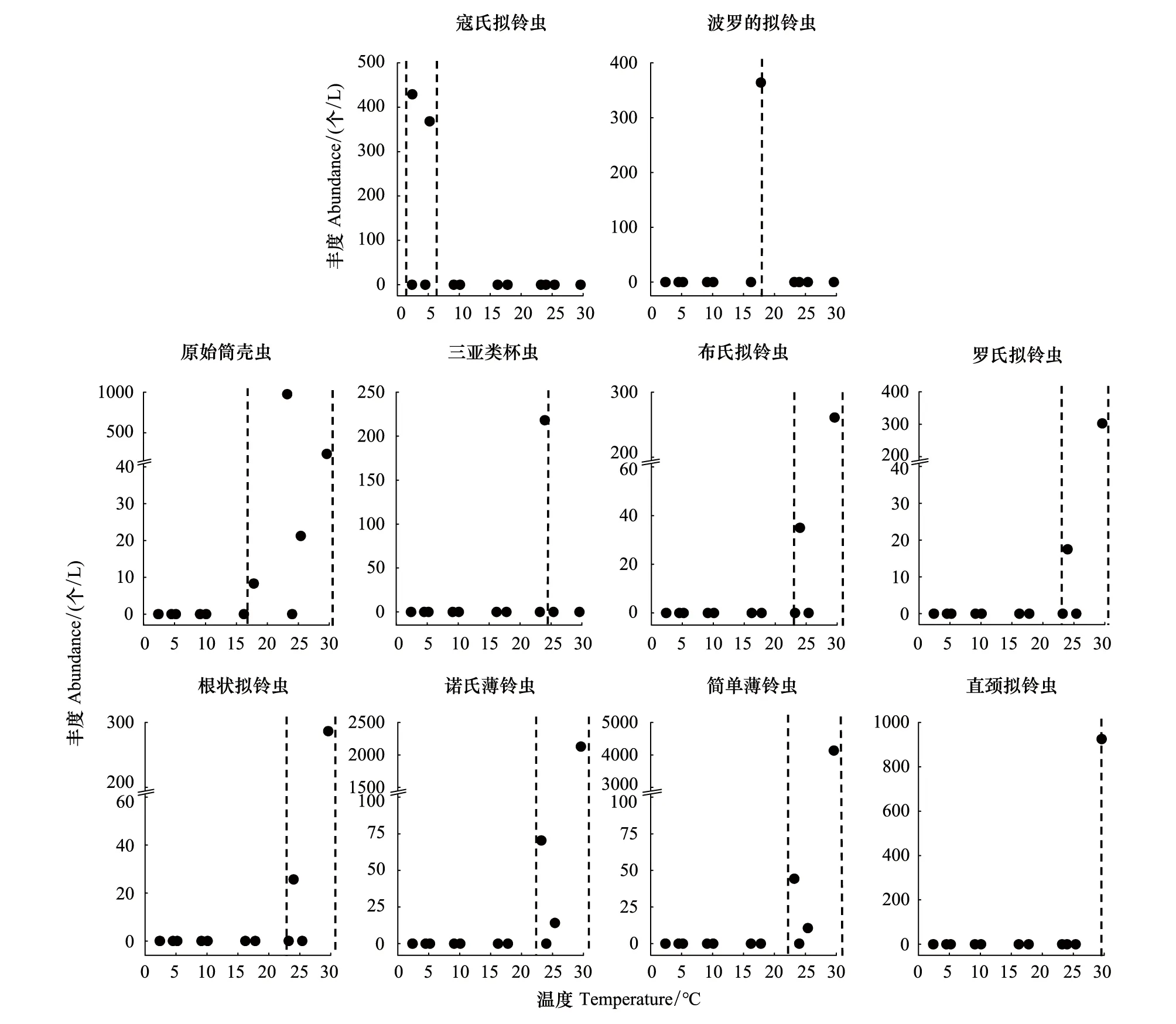
圖5 豐度較高砂殼纖毛蟲種類出現的溫度范圍Fig.5 Temperature range in which abundant tintinnids occurred
2.4.2群落結構
砂殼纖毛蟲種類組成周年變化明顯,不同種群出現季節更替。2、3月寇氏擬鈴蟲占優,5月波羅的擬鈴蟲占優,6月三亞類杯蟲占優,7月簡單薄鈴蟲、諾氏薄鈴蟲及直頸擬鈴蟲豐度較高,8月長形旋口蟲(Helicostomellalonga)及原始筒殼蟲豐度較高,9月原始筒殼蟲占優,10月卡拉直克擬鈴蟲(Tintinnopsiskarajacensis)占比100%,11月筒狀擬鈴蟲(Tintinnopsistubulosoides)及愛氏網紋蟲(Favellaehrenbergii)占優(圖6)。Mantel 檢驗表明,驅動砂殼纖毛蟲群落周年變化的主要環境因子是溫度,次要因子是鹽度(表2)。

表2 砂殼纖毛蟲群落結構(用Bray-Curtis相異度測量)和環境因子的Mantel檢驗Table 2 Mantel test between tintinnid community (measured as Bray-Curtis dissimilarity) and environmental factors

圖6 砂殼纖毛蟲種類組成的周年變化Fig.6 Annual variation of species composition of tintinnids
砂殼纖毛蟲群落Shannon指數的平均值為0.95±0.78,在6—8月較高,7月最高(2.11);Pielou指數的平均值為0.52±0.34,在6—8月及12月較高,12月最高(1.00)。
2.5 纖毛蟲與環境因子相關性分析
無殼纖毛蟲豐度與溫度、鹽度和Chl a濃度均沒有明顯相關性。砂殼纖毛蟲豐度、纖毛蟲總豐度及砂殼纖毛蟲占纖毛蟲總豐度的比例均與溫度呈極顯著的正相關;砂殼纖毛蟲豐度和纖毛蟲總豐度均與鹽度呈極顯著的負相關,砂殼纖毛蟲占纖毛蟲總豐度的比例與鹽度呈顯著的負相關;砂殼纖毛蟲豐度、纖毛蟲總豐度及砂殼纖毛蟲占纖毛蟲總豐度的比例均與Chl a濃度呈極顯著的正相關(表3)。

表3 纖毛蟲豐度及砂殼纖毛蟲占纖毛蟲總豐度的比例與溫度、鹽度及Chl a濃度的相關性分析Table 3 Correlation between ciliate abundance,and percentage of tintinnids to total ciliate abundance and water temperature,salinity and Chl a concentration
3 討論
近年來,多數研究主要關注纖毛蟲豐度、種類組成與環境因子的關系[21—23]及物理過程如水團等對纖毛蟲豐度分布的影響[24—27],只有少數研究關注纖毛蟲群落組成長時間序列的變化[28—29]。本文在渤海灣近岸海區開展纖毛蟲豐度和種類組成的周年調查,補充渤海灣纖毛蟲長時間序列的數據。
與其它海區相比,本次調查中纖毛蟲豐度和生物量均處在已有文獻的范圍之內(0—145000 個/L,0—400 μg C/L)[30];與之前渤海灣的研究[11—12]相比,本次調查中無殼纖毛蟲豐度相差不大,砂殼纖毛蟲豐度明顯較高;與獐子島海區[31]、膠州灣[9]周年資料對比(表4),渤海灣纖毛蟲豐度較低,砂殼纖毛蟲豐度占纖毛蟲總豐度的比例稍高。
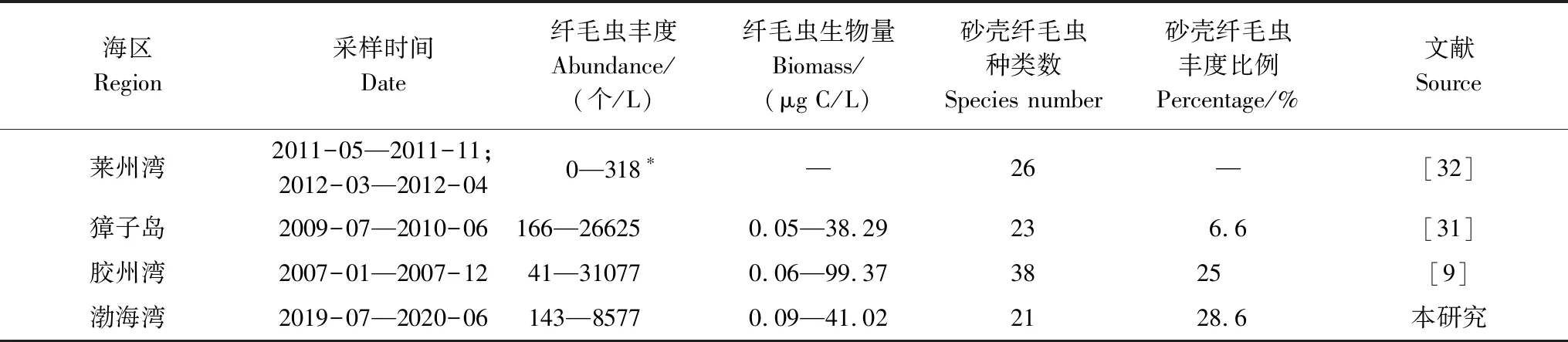
表4 我國海區浮游纖毛蟲群落的周年變化數據Table 4 Annual studies of planktonic ciliate communities in China seas
3.1 纖毛蟲豐度和生物量的周年變化
本研究中纖毛蟲豐度和生物量分別在春季和夏季出現峰值,表現出典型的溫帶近岸海區的周年變化規律。溫帶近岸海區如Gdansk Basin[33]、Ka?tela Bay[34]、膠州灣[9]等,纖毛蟲豐度和生物量通常在春、夏季或者春、秋季呈現雙峰型,這可能與餌料水平(Chl a濃度)的周年變化密切相關。與多數研究類似[35—37],渤海灣纖毛蟲豐度和生物量與Chl a濃度呈極顯著的正相關,其兩個峰值與Chl a濃度的峰值基本吻合,稍有滯后,這說明上行控制在渤海灣纖毛蟲豐度和生物量周年變化中扮演重要的角色。
渤海灣無殼纖毛蟲和砂殼纖毛蟲出現峰值的時間存在較大差異,這可能是由于無殼纖毛蟲和砂殼纖毛蟲在不同營養條件下的競爭力不同導致的。砂殼纖毛蟲攝食的餌料粒級較大(多為nano級浮游植物),且由于肉體外具殼需要的餌料更多,因此在寡營養條件下(pico級浮游植物占優)生長能力較低[38];在較高餌料條件下(nano級浮游植物占優),砂殼纖毛蟲具有更高的競爭力,豐度往往較高,有時會超過無殼纖毛蟲在豐度上占優勢[39]。本研究中7月砂殼纖毛蟲豐度高達8440 個/L,遠超無殼纖毛蟲占絕對優勢,這可能與7月較高水平的餌料條件(用Chl a濃度指示)有關。
3.2 砂殼纖毛蟲種類
渤海灣砂殼纖毛蟲各種類只在某些月份出現,不同種群更替有序,砂殼纖毛蟲群落組成呈現明顯的周年變化。與萊州灣的研究結果[32]不同,本次渤海灣調查中只記錄了冬季種、春季種及夏季種,沒有發現周年出現種。
本研究中大部分砂殼纖毛蟲豐度都偏向在溫度較高的夏季較高,值得注意的兩種砂殼纖毛蟲——波羅的擬鈴蟲和寇氏擬鈴蟲,其偏好的水溫較低(低于18℃)。波羅的擬鈴蟲只在水溫為17.8 ℃的5月出現,其他月沒有記錄,這說明波羅的擬鈴蟲偏好溫度適中的環境,在溫度較高或較低時不適合生長[40]。寇氏擬鈴蟲只在溫度低于5.2 ℃的2、3月出現,是一種喜低溫的砂殼纖毛蟲,這與膠州灣的研究結果[41]不同,膠州灣寇氏擬鈴蟲一般在溫度為5—15 ℃時豐度較高。這種差異可能是由于本研究采樣站位單一的局限性導致,需要在今后積累更大范圍的樣品來得出更準確的結論。喜低溫砂殼纖毛蟲種類的出現可以作為冷水團的指示種,在浮游纖毛蟲與物理過程的耦合關系研究中發揮重要的指示作用。
3.3 渤海的新紀錄種——三亞類杯蟲
本研究在渤海首次紀錄到三亞類杯蟲(圖7),該種類為南方種[42],一般在我國近岸36°N以南海區有發現,如膠州灣[42]、東海[43—44]及南海近岸[45—46],本研究第一次在我國36°N以北海區紀錄到三亞類杯蟲,且與其它海區相比,渤海灣三亞類杯蟲個體稍大(表5)。
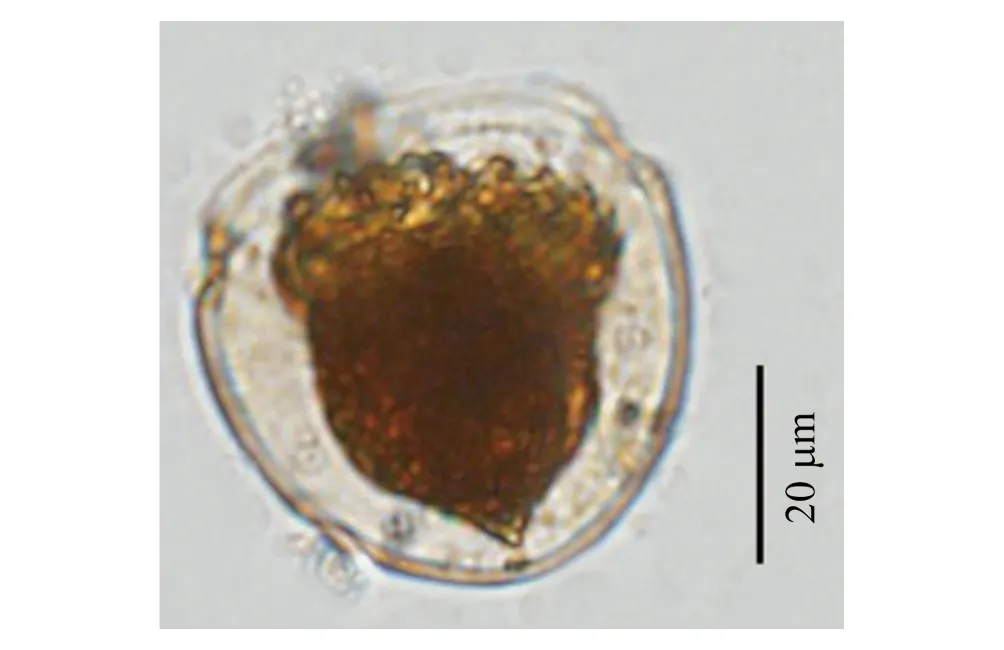
圖7 三亞類杯蟲顯微照片Fig.7 Micrograph of Metacylis sanyahensis
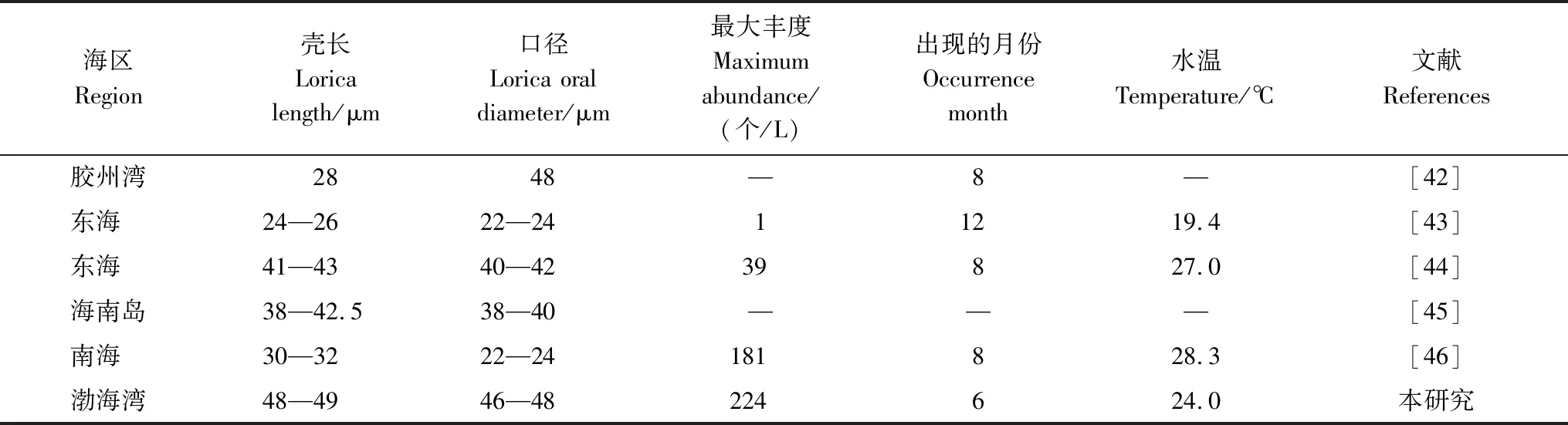
表5 我國海區的三亞類杯蟲Table 5 Metacylis sanyahensis in China seas
現有資料表明,三亞類杯蟲在我國海區豐度不高,出現頻率也較低,一般趨向分布在溫度較高的夏季,在其它季節很少觀測到。作為喜高溫的南方種,三亞類杯蟲北移,在渤海灣出現,可能與全球變暖有關,可以作為暖水團的指示種在微型浮游動物與水團的耦合關系研究中發揮重要的指示作用。
