溫度升高對長江口蘆葦濕地細根形態和生長的影響
2022-05-26 10:02:02仲啟鋮張桂蓮王開運鄭思俊王小玲何晶李德志
生態學報 2022年9期
關鍵詞:生長
丁 一,仲啟鋮,張桂蓮,王開運,鄭思俊,王小玲,何晶,李德志
1 上海市園林科學規劃研究院,城市困難立地生態園林國家林業和草原局重點實驗室,上海城市困難立地綠化工程技術研究中心,上海 200232 2 華東師范大學生態與環境科學學院,上海市城市化生態過程與生態恢復重點實驗室,上海 200241
據IPCC預測,到21世紀末,全球地表平均溫度將較工業革命前上升1.5℃以上(IPCC 2014)。空氣、土壤和水體的溫度升高將會對人類賴以生存的多種生態系統的結構和功能產生深遠影響[1]。濱海濕地位于陸地和海洋生態系統交界處,植被生產力高、有機碳分解速率低、土壤碳積累速率快[2],具有保育水生生物多樣性、向近岸水域輸送營養物質、過濾降解污染物、保護海岸線[3]以及作為藍色碳匯[4—6]吸收和固持大氣中的CO2等重要生態系統服務。目前已有一些研究采用室內培養、模擬增溫和氣候梯度等方法研究了升溫對濱海濕地植物的影響及其反饋[3,7—9]。這些研究發現升溫可能會增強外來入侵植物的擴散[9],改變植物群落結構[8],影響植物的生理和生長指標等。如Charles和Dukes[3]發現升溫提高了北美鹽沼優勢植物互花米草(Spartinaalterniflora)的株高、基徑和生物量。Kirwan等[10]預測2—4℃的升溫將使互花米草群落的地上生產力提高10%—40%。李沙沙等[11]發現升溫顯著提高了崇明東灘蘆葦(Phragmitesaustralis)和互花米草的單株葉面積、頂葉面積和葉面積指數。但是,與地上部分相比,目前關注全球變暖對濱海濕地植物地下部分影響的研究相對較少,而對地下部分最為活躍、最為敏感的濕地植物細根會對升溫條件產生何種響應則更不清楚。
根系是植物最重要的功能器官,超過一半光合作用生產的有機碳會被植物分配到地下部分用于根系生長[12—13],并通過根系周轉產生的殘體和根系沉積輸入根際土壤[14]。盡管直徑≤2 mm的細根在總根生物量中僅占不到20%,但由于其處于不斷的周轉過程中,每年約1/3的全球凈初級生產力被消耗用于生產和維持細根[15—16]。細根還是土壤中有機碳的重要來源,植物分配到地下部分的有機碳有15%—25%會通過細根周轉輸送到土壤碳庫中[17—18],是陸地生態系統碳循環的重要環節之一。作為植物根系中活性最強、周轉最快的組分,直徑≤2 mm的細根對全球氣候變化的反應更為敏感,并顯著影響生態系統地下部分的碳平衡和養分循環[19],因此成為全球氣候變化研究的熱點之一。研究表明,升溫對細根形態和生長的影響可能與物種[20]、微地形[21]、營養物質有效性[22]、土壤含水率[23]等有關,不同植物細根生產和死亡節律對升溫響應的不一致性也會造成不同的細根功能性狀變化[23]。Wang等[24]綜述了177篇相關文獻發現,升溫總體上增加了細根生物量和生產量,但對其長度、形態和周轉影響不顯著,但這一研究缺乏來自濱海濕地的數據。微根管技術是目前研究細根生長動態的主流方法之一。與傳統的土鉆法[25]、內生長法[26—27]和挖掘法等方法相比,該方法通過在土壤中安裝透明微根管,使用小型攝像機或掃描儀對細根進行長期、定點和非破壞性的連續監測[28],能直接捕捉不同土壤層次中細根的生長、死亡和周轉等動態變化[29]。但目前這一方法在濕地生態系統中的應用還比較少見[30]。
崇明島位于長江入海口,其沿河口海岸線分布的大片蘆葦濕地為超大城市上海提供了多種重要的生態系統服務。項目組前期在崇明東灘的模擬升溫實驗發現,氣溫升高對蘆葦濕地的土壤碳排放無顯著影響,但卻顯著增加了其植被地上生物量[31—32];升溫顯著增加了表層土壤總有機碳含量,但對微生物生物量和相關酶活影響較小[33]。鑒于蘆葦細根的活性和周轉速率遠高于根狀莖,且其主要分布在0—40 cm土層[34],為了解升溫條件下細根的潛在變化及其對土壤碳平衡的可能影響,本研究利用開頂式生長箱法對蘆葦濕地的原狀植物—土壤連續體進行模擬升溫。2019年5—10月,采用微根管法和根鉆法,對0—40 cm土層細根(本研究中指直徑≤2 mm蘆葦須根)的數量、長度、直徑、表面積和生物量等性狀特征開展了為期一個生長季的連續觀測,并計算細根的凈生長速率和周轉速率,從而探討氣溫升高對蘆葦濕地細根形態、生長特征及土壤碳平衡的潛在影響,以期了解細根介導的地下碳循環對全球變暖的響應機理,并為開展濱海蘆葦濕地碳匯功能的保護和修復提供科學依據。
1 研究區域與研究方法
1.1 研究區域概況
研究區域位于上海市崇明島東灘 (31°38′N,121°58′E),該地區分布有長江口規模最大、發育最完善的濱海鹽沼濕地之一。該區域屬北亞熱帶海洋性季風氣候,年平均氣溫15.3℃,年平均降水量為1004 mm。土壤類型為濱海鹽堿土,以河口沉積物為母質。土壤含鹽量在0.1‰—5.0‰之間,pH在8.2—9.2之間,有機質和養分含量較低[33]。崇明東灘也是長江口圍墾規模最大的地區之一。樣地所處地塊在1998年被人工建設的堤壩圈圍之前,是發育良好濱海鹽沼濕地的高潮灘,以蘆葦為原生優勢種。該地區被堤壩圈圍后,不再受潮汐周期性影響,但由于地勢較為低平,在豐水期(如7—9月份)及降水豐富時地表仍不時出現淹水狀態,年均水位約為地表以下30 cm。常見優勢植物包括蘆葦(P.australis)、白茅(I.cylindrica)、拂子茅(Calamagrostisepigeios)和荻(Triarrhenasacchariflora)等。
1.2 研究方法
1.2.1樣方建設
開頂式生長箱(open top chamber,OTC)最早建設于2008年。選取地勢均勻、植被均一的地塊設置樣地,OTC由8 mm厚的透明氟化玻璃和不銹鋼框架制成,箱體高3.5 m,底部和頂部呈八邊形,底部面積為12.5 m2,頂部開口略收緊[31],除必要結構性部件外,箱體均采用透光材料,并且底部留有通風口。將4個建設有OTC的樣方作為升溫樣方(ET),同時緊鄰每個OTC,等距離、隨機性設置等面積、無OTC的對照樣方(CK),具體樣方設置如圖1所示。2019年的溫度監測結果表明,4—11月份空氣升溫幅度為1.41℃,土壤升溫幅度為0.75℃,其中,7、8月份氣溫升幅最高。
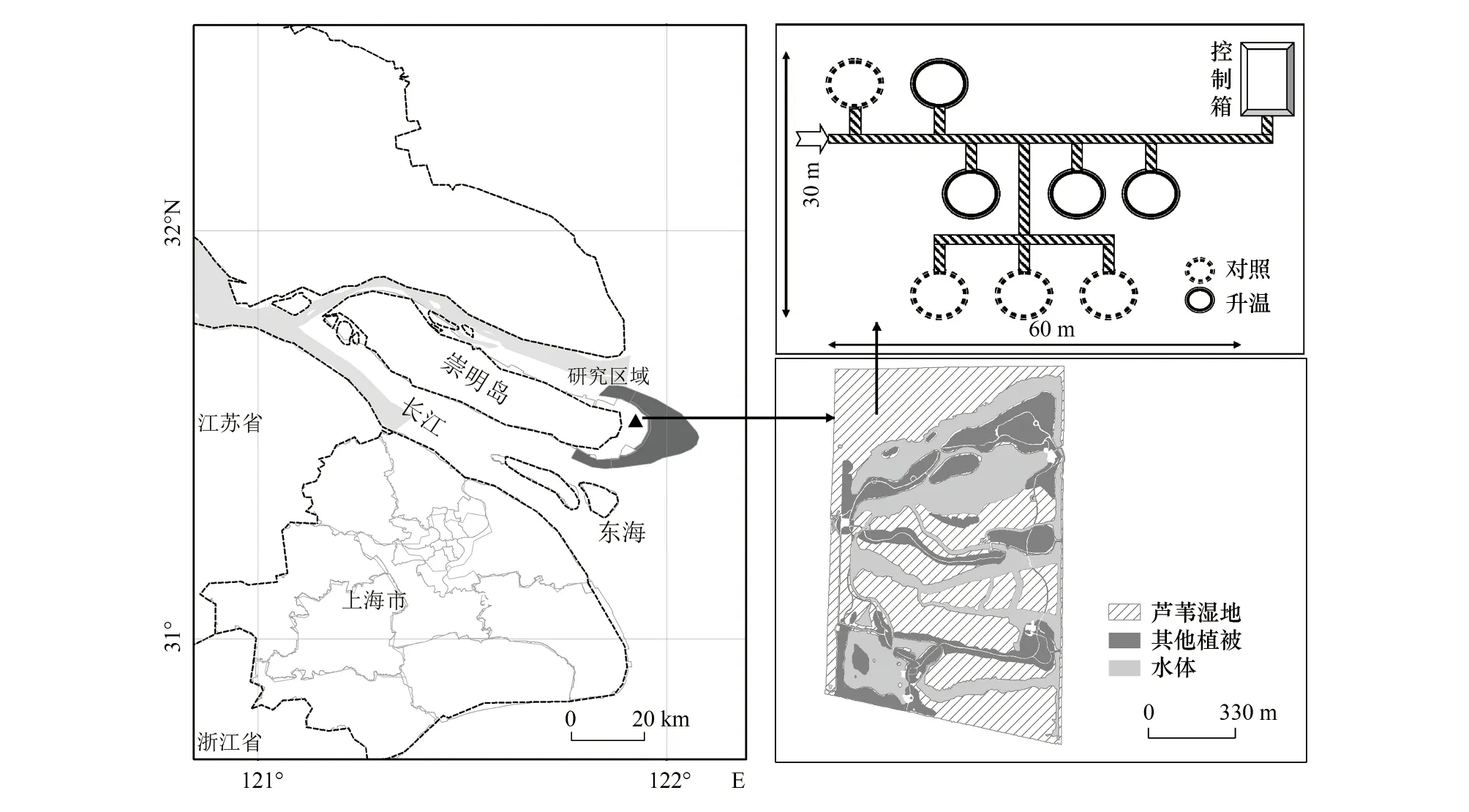
圖1 研究區域及試驗樣地設置Fig.1 The location of the study area and the patterns of plots configurationCK:對照樣方Control check;ET:升溫樣方Elevated temperature plots
目前,該樣地的濕地植被群落已經退化,蘆葦不再是單一的優勢種。為嚴格模擬氣溫升高對長江口原生蘆葦濕地植被的影響,2018年12月,于崇明東灘鳥類自然保護區內選取以蘆葦為單優勢種,植株高度、密度均一的濕地斑塊(約20 m×20 m大小),間隔2—3 m、相互無干擾地挖掘植物—土壤連續體(長×寬×深:0.6 m×0.6 m×0.6 m),并移栽至升溫設施所在樣地。在每個樣方內布設成對角分布的2個小樣方,共16個小樣方。在每個小樣方內挖設長寬皆略大于原狀土塊,深度為0.6 m的坑體,并用厚度為5 mm的不銹鋼板將四周圍隔。將挖掘的原狀蘆葦植物—土壤連續體隨機置放在16個小樣方中,然后用移栽現場取回的土壤將空隙填實。ET和CK中土壤的初始理化性質如表1所示。

表1 試驗開始前ET和CK中移栽樣方各層的土壤理化性質Table 1 Soil physical and chemical properties of the transplanted plots in ET and CK in different soil layer before the warming treatment
在每個樣方中,選取其中一側小樣方,首先用特制鋼鉆,以與地面成45°的夾角,鉆一個與微根管直徑相近的管洞,然后將有機玻璃質的透明微根管斜插入土壤,共埋設8 根(其中ET和CK樣方各4根);另一側則用于使用土鉆進行根系定期取樣。微根管外徑 7 cm,內徑 6.4 cm,長100 cm,埋入土中65 cm,垂直深度超過40 cm。在安裝微根管時將密封段插入土壤中,并將管壁緊貼土體。微根管露出地表部分用黑色塑料布裹住,頂端開口處用黑色蓋子密封,以避免水分和灰塵進入管中。
1.2.2細根圖像的采集與處理
在完成植物移栽和微根管埋設后,用5個月的平衡期使根系生長和土壤恢復穩定。2019年5—10月,使用CI—600根系生長監測系統(CID BioScience Inc.)采集微根窗內的根系圖像,每根微根管采集3張圖像,每張圖像面積為20.8 cm×21.7 cm。將圖像按照微根管45°的傾角,劃分到0—10 cm,10—20 cm以及20—40 cm土層。每月采集一次圖像,整個生長季內共獲取6次。采用Winrhizotron MF根系分析軟件(Regent Instrument Inc.)進行圖像數據的處理和分析,獲取和計算微根窗內直徑≤2 mm細根的總根長(Total root length TRL)(cm)、總表面積(Total surface area,TSA)(cm2)、平均細根直徑(Average root diameter,ARD)(mm)、根長密度(Root length density,RLD)(cm/cm3)以及凈生長速率(Net growth rate of root length density,RLDNGR)(cm cm-3d-1)。其中,RLDNGR通過計算一定觀測時間段內RLD的變化來計算,公式如下:
RLDNGR=(RLDn+1-RLDn)/T
(1)
式中,RLDn+1與RLDn分別表示第n+1次與第n次觀測到的細根根長密度,T為相鄰兩次觀測的間隔天數。
RLD的計算公式如下[35]:
RLD=RL/A×DOF
(2)
A=C根管×D/sin45°
(3)
式中,RL為圖片觀測到的細根根長;A為微根窗觀測區域的面積,C根管為根管圖片的邊長,即20.8 cm;D為劃分的土層深度,即10 cm或20 cm;DOF為田間觀測深度,即根管壁與土壤之間的距離,可計算獲得,但會破壞樣方;也可采用經驗值,本中統一取0.2 cm[36]。
1.2.3細根樣品的采集與處理
2019年6月期,在每個樣方中無根管的一側小樣方選點,用內徑5 cm土鉆分別鉆取 0—10 cm,10—20 cm和20—40 cm 土芯,帶回試驗室流水沖洗,手揀挑出全部根系并區分出直徑≤2 mm細根,同時通過顏色區分死活根。通過LA-S根系掃描儀(萬深檢測科技有限公司)獲取直徑≤2 mm的活細根圖像,并采用WinRHIZO Tron MF根系分析軟件測定根長。將掃描好的根系裝入牛皮紙信封內,65℃條件下烘干至恒重后稱取生物量。采用如下公式計算比根長(SRL)(m/g)、比表面積(SRA)(cm2/g):
SRL=RL/B
(4)
SRA=RSA/B
(5)
式中,RL為掃描圖片觀測到的細根根長,RSA為掃描圖片觀測到的細根表面積,B為用根鉆取出、洗凈、烘干并稱重的細根干物質量(g)。通過RLD和SRL計算單位體積細根生物量密度(RBDV)(g/m3),計算公式如下:
RBDV=RLD/SRL
(6)
單位面積的細根生物量RBDA(g/m2)通過單位體積細根生物量密度乘以取樣深度D(m)來換算:
RBDA=RBDV×D
(7)
1.2.4細根周轉速率計算
細根周轉速率(T(a-1)采用極差法計算[37—38],公式如下:
T=BNPP/Bavg
(8)
BNPP=RBDAmax-RBDAmin
(9)
式中,BNPP為年凈細根生產力(g m-2a-1),RBDAmax、RBDAmin分別為一個生長季中單位面積細根的最高生物量和最低生物量(g/m2),Bavg為月現存生物量平均值。
1.3 數據統計分析
采用SPSS 20.0對數據進行統計分析。在整個0—40 cm土層以及0—10、10—20、20—40 cm三個土層分別采用重復測量方差分析(RMANOVA),檢驗升溫和觀測時間對TRL、TSA、SRL、SRA、RBDA、ARD和RLDNGR等細根形態和生長指標的影響;對于升溫與觀測時間存在顯著交互效應的指標,進一步采用單因素方差分析(One-way Anova),分別在每個觀測時間點上,檢驗升溫對細根形態和生長指標的影響。所有統計結果中,均以P<0.05作為存在顯著性差異的標準。
2 結果與分析
2.1 升溫在整個生長季對細根的影響
重復資料方差分析表明(表2),在整個生長季內,升溫在0—40 cm土層顯著增加了細根的總根長、總表面積、總生物量和凈生長速率,增加幅度分別為125.8%、82.9%、100.5%和133.3%(表3),且觀測時間與升溫對這四個指標的影響均存在顯著交互作用。升溫對細根平均直徑、比根長、比表面積無顯著影響。
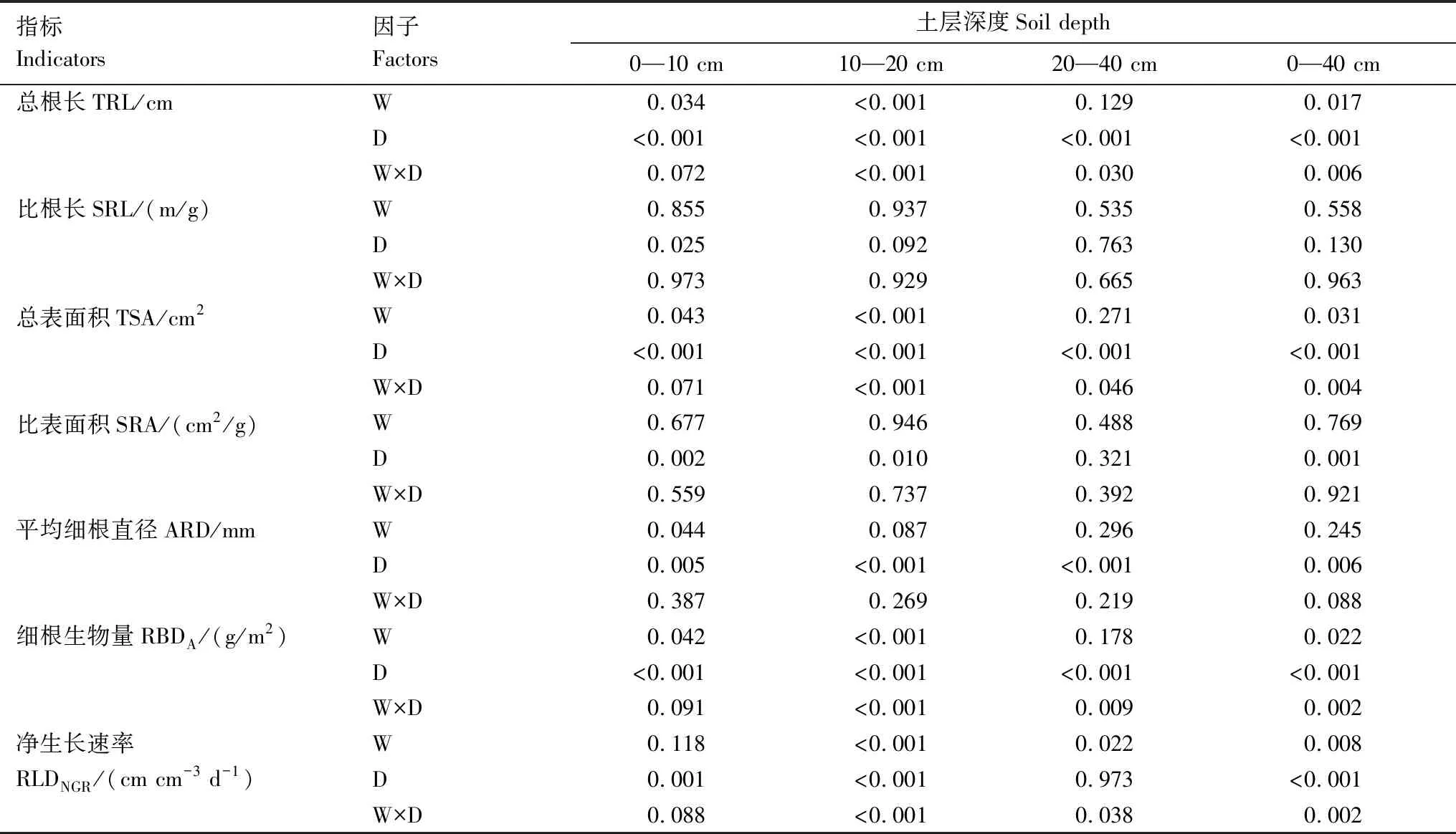
表2 整個生長季不同土層細根形態和生長指標的重復測量方差分析結果Table 2 Repetitive Measure ANOVA (RMANOVA)on the morphological and growth traits of fine roots of P.australis in different soil layer during the growing season

表3 整個生長季不同土層細根形態和生長指標的比較(平均值±標準誤)Table 3 Comparisons of the morphological and growth traits of fine roots of P.australis in different soil layer during the entire growing season (mean±SE)
分層來看,升溫處理顯著增加了0—10 cm和10—20 cm土層細根的總根長、總表面積、細根生物量和凈生長速率,且觀測時間與升溫處理的交互作用顯著。升溫還使0—10 cm土層細根的平均直徑顯著降低了13.2%。升溫處理對0—10 cm和10—20 cm土層細根的比根長以及比表面積均未產生顯著影響。與0—10 cm和10—20 cm土層相比,升溫對20—40 cm土層細根的影響較小,僅顯著提升了其凈生長速率這一項指標。
2.2 升溫在不同月份對細根的影響
如圖2所示,在整個0—40 cm土層,CK與ET中細根總根長均呈現逐月上升的趨勢,10月達到最大值,且前者月平均值始終高于后者。其中,5、8、9、10月,升溫分別將細根總根長顯著增加了67.1%、117.3%、149.5%和118.3%。在0—10 cm土層,除7月外,ET和CK中細根總根長在另5個月均差異顯著,其中8月增幅最大,為166.2%;在10—20 cm土層,升溫在除5月外的其他月份顯著增加了細根總根長,其中9月增幅最大,達219.4%;在20—40 cm土層,升溫僅在10月顯著增加了細根總根長,增幅為90.1%。

圖2 模擬升溫對不同土層、不同觀測時間細根總根長的影響Fig.2 Effects of experimental warming on total root length of fine roots of P.australis at different sampling date in different layer不同大寫字母表示相同月份不同處理差異顯著(P<0.05),不同小寫字母表示相同處理不同月份間差異顯著(P<0.05)
如圖3所示,在整個0—40 cm土層,與總根長類似,CK與ET的細根總表面積同樣呈現逐月上升趨勢,且ET始終高于CK。其中8、9、10月二者差異顯著,ET分別比CK分別高出93.1%、106.7%和96.4%。在0—10 cm土層,6、8、9月差異顯著,其中8月增幅最大,為148.3%。在10—20 cm土層,除5月外,升溫顯著提高了細根總表面積,其中9月增幅最大,達153.9%。在20—40 cm土層,升溫對細根總表面積無顯著影響。
施藥后7、15 d觀察各處理的小麥生長情況,1%甲基二磺隆·雙氟磺草胺可分散油懸浮劑各劑量對小麥安全性好,無明顯藥害癥狀;藥劑 30 g/L 甲基二磺隆可分散油懸浮劑、50 g/L雙氟磺草胺懸浮劑對小麥也無藥害癥狀。收獲時測產結果表明,各藥劑處理小區產量要低于人工除草處理,與空白對照相比,均有不同程度的增產作用,增產率為7.84%~11.46%(表4)。

圖3 模擬升溫對不同土層、不同觀測時間細根表面積的影響Fig.3 Effects of experimental warming on total root surface area of fine roots of P.australis at different sampling date in different layer
在整個0—40 cm土層(圖4),升溫顯著提高了8、9、10月的細根生物量,分別達到100.7%、149.4%、122.5%。在0—10 cm、10—20 cm土層,除7月外,其他月ET的細根生物量均顯著高于CK。其中,0—10 cm土層的細根生物量增幅8月最高,達143.3%;10—20 cm土層的細根生物量增幅9月最高,為274.4%;在20—40 cm土層,升溫僅在10月顯著增加了細根生物量。

圖4 模擬升溫對不同土層、不同觀測時間細根生物量的影響Fig.4 Effects of experimental warming on root biomass density per unit area of fine roots of P.australis at different sampling date in different layer
如圖5所示,在整個0—40 cm土層,CK與ET細根凈生長速率在整個生長季中均呈“M”型曲線:即先升高后降低、再升高再降低。ET中細根凈生長速率的季節波動幅度明顯高于CK。在生長季初級T1(5—6月)、末期T5(9—10月),CK與ET細根凈生長速率無顯著差異;但在T2(6—7月)、T3(7—8月)、T4(8—9月),ET的細根凈生長速率顯著高于CK。其中,T4階段升溫對細根凈生長速率的增幅最大(134.1%)。

圖5 模擬升溫對不同土層、不同觀測時間細根凈生長速率的影響Fig.5 Effects of experimental warming on net growth rate of root length density of fine roots of P.australis at different sampling date in different layer
同樣的“M”曲線趨勢出現在每個土層,但其波動幅度隨土層加深趨于平緩。在0—10 cm土層,ET細根的凈生長速率在T4顯著高于CK,在T5顯著低于CK;在10—20 cm土層,ET細根的凈生長速率在T3和T4顯著高于CK;在20—40 cm土層,ET細根的凈生長速率僅在T4顯著高于CK。
2.3 升溫對細根垂直分布的影響
排除體積影響后發現,升溫處理顯著改變了蘆葦細根在不同土層的分布(圖6)。在CK中,除20—40 cm土層細根比表面積顯著高于另外兩個土層外,其他指標在不同土層之間均無顯著差異。在ET中,20—40 cm土層細根比表面積與另兩個土層無顯著差異;0—10 cm、10—20 cm土層細根根長密度、表面積密度、生物量和凈生長速率均顯著高于20—40 cm土層。總體來看,升溫使更高比例的細根分布在0—20 cm土層中。

圖6 模擬升溫對0—40cm 土層細根垂直分布的影響Fig.6 Effects of experimental warming on vertical distribution of fine roots in the 0—40 cm layer不同小寫字母表示相同處理不同土層間差異顯著(P<0.05)
2.4 升溫對細根周轉速率的影響
如圖7所示,升溫顯著提高了10—20 cm以及20—40 cm土層的細根周轉速率,增幅分別為32.9%和94.6%。然而,升溫對0—10 cm土壤表層的細根周轉速率影響并不顯著。對比不同土層,在CK中,細根周轉速率隨土層加深呈下降趨勢;但在ET中,不同土層之間的細根周轉速率無顯著差異。

圖7 模擬升溫對不同土層細根周轉速率的影響Fig.7 Effects of experimental warming on turnover rate of fine roots of P.australis*表示相同土層不同處理間差異顯著(P<0.05),不同小寫字母表示相同處理不同土層間差異顯著(P<0.05)
3 討論
3.1 升溫對蘆葦濕地細根形態和生長的總體影響
本研究中,升溫在整個生長季中顯著提高了長江口蘆葦濕地0—40 cm土層細根的生物量、總根長與總表面積,同時也增加了其凈生長速率。這與Coldren等[20]通過原位升溫試驗發現的紅樹林濕地地下根系生物量升高的結果一致。Sullivan等[21]同樣發現升溫會導致北極沼澤植物的細根生物量增加。研究表明,如果土壤水分和養分有效性足夠,根生物量和生長速率會隨著溫度升高而增加[39]。項目組前期研究發現,升溫會使蘆葦濕地土壤微生物氮、磷及相關酶活顯著提升,增加微生物礦化作用[40—41],使植物可利用土壤養分增加,從而促進根系生長。同時,升溫還會增大蘆葦的光合作用[42]和總葉面積[11],光合產物總量增加會使分配給細根的量增加。另一方面,氣溫升高和葉面積指數同時增加[11]會顯著增強蘆葦的蒸騰作用,蘆葦可能選擇采用更長、更高表面積的細根來增加對土壤水分的吸收能力,從而維持其正常的生理代謝活動。
細根的比根長和比表面積反映了細根吸收土壤水分與養分的效率[43]。比值越大,相同的細根生物量會有更高的水分和養分吸收效率。平均細根直徑同樣可以表征植物細根吸收土壤養分、水分策略,也影響著細根的周轉和壽命(周轉的倒數)。本研究中,升溫未對蘆葦濕地細根的比根長、比表面積、平均細根直徑產生顯著影響。類似的,Bj?rk等[44]在挪威北部進行的模擬升溫試驗也發現苔原濕地干草甸群落植物的比根長、比表面積未發生顯著變化,但干石南灌叢群落細根的比根長與比表面積卻顯著升高。本研究中,0—40 cm的細根比根長、比表面積和直徑未發生顯著變化,其可能原因包括:(1)由于濕地水分和養分可用性高,蘆葦無需明顯改變其細根水分和養分吸收效率以應對升溫對其光合作用和蒸騰作用的影響。(2)對根系較細的物種來說,調節比根長的能力相對較弱[45]。上述結果表明,在長江口蘆葦濕地中,升溫可能會通過促進細根的伸長來增加其總表面積和總生物量,而不會顯著改變細根的形態特征,以及水分和養分吸收效率。
細根以周轉方式進入到土壤的碳含量約占土壤碳總輸入量的14%—50%[12],細根周轉深刻地影響著土壤碳循環過程。Fitter等[46]在對英國一個高地草原的土壤升溫試驗中發現,升溫會導致細根周轉速率的增加,這與我們的研究結果一致。導致這一現象的原因可能是由于升溫條件下細根的生理活性增加、壽命變短,同時微生物礦化作用增強使養分供應增加[47],進而促進細根新陳代謝增強、周轉速率加快;另一個可能的原因是升溫導致細根總量和密度增加,細根之間競爭加劇導致細根生產、死亡加速[48],周轉加快。
3.2 升溫對蘆葦濕地細根形態和生長時空格局的影響
植物根系的季節動態反映了其物候,生境條件變化如土壤水分、養分的改變通過影響植物細根的水養吸收而改變其季節動態[49],而光合作用強度通過影響光合產物向地下的運輸和分配影響細根的生長動態[50]。本研究中,蘆葦濕地細根的生物量、總根長、總表面積持續呈逐月升高趨勢,即生產大于死亡,現存量持續增加。升溫處理提高了濕地細根現存量的增加幅度,使升溫與對照的差值逐月擴大。細根生長的季節動態有單峰型曲線、雙峰型曲線[51—52]。本研究中,蘆葦細根在整個生長季中的凈生長速率呈雙峰型曲線,且升溫導致其季節變異性進一步增大。升溫在6—9月持續促進了整個0—40 cm土層的細根凈生長速率,其中在8—9月最為顯著。7—8月蘆葦細根凈生長速率相較于6—7月、8—9月相對降低,可能是由于上海2019年梅雨季持續到7月下旬,連續陰雨天氣導致蘆葦光合作用強度降低,影響了光合作用產物的生產和分配,從而影響植物細根的生長。
細根的垂直分布格局顯示了蘆葦的營養獲取策略,與物種、氣候因子以及土壤理化性質密切相關。一般認為,植物細根的分布隨土層加深而減少[53—55]。本研究中,對照中不同土層細根根長和生物量的差異不顯著。升溫使0—20 cm土層細根總根長、總表面積和生物量顯著增加,高于20—40 cm土層,這說明升溫條件下,細根更加趨向于集中在表層0—20 cm分布[44]。這種垂直梯度上細根分布的變化可能與升溫后不同土層中養分的有效性或養分向根的擴散速率的變化有關。Zhong等[33]發現升溫使蘆葦濕地0—10 cm土層總有機碳、總氮顯著提升,而在10—40 cm土層無顯著影響。另外,本研究中,升溫在不同土層對細根比根長、比表面積、平均細根直徑影響均不顯著,說明不同土層根系水分和養分吸收效率均未受顯著影響。
升溫對細根凈生長速率的影響在不同土層也存在一定差異。其中,與10—20 cm和20—40 cm土層相比,升溫對0—10 cm土層細根凈生長速率提升幅度最小,僅在根系生長最快的6—7月、8—9月呈現顯著促進作用。這可能是因為該地區土壤為濱海鹽堿土,升溫條件下蒸發散作用增強會導致表層土壤含鹽量顯著提高[33],土壤含鹽量增加則可能會抑制了細根的正常生長[56—57],限制了升溫在某些月份對細根凈生長速率的促進作用。
升溫對不同土層的細根周轉速率產生了不同的影響,升溫處理顯著提升了10—20 cm、20—40 cm土層的細根周轉速率,但對0—10 cm土層無顯著影響,導致升溫后不同土層的細根周轉速率之間差異不顯著。與凈生長速率一樣,0—10 cm土層細根周轉速率未產生顯著變化,同樣可能是由于表層土壤含鹽量增加[56—57],從而對細根的代謝和周轉產生了潛在抑制。
3.3 升溫條件下蘆葦細根變化對土壤碳平衡的潛在作用
項目組前期研究發現,升溫導致蘆葦濕地土壤表層有機碳增加,但對微生物生物量碳無顯著影響,我們推測這種現象與升溫引起的根源有機碳輸入的增加有關。本研究中,結合對根系周轉速率及其有機碳含量的測定結果,對根系周轉的年度有機碳輸入量進行估算。估算結果顯示,升溫條件下,蘆葦濕地通過細根周轉進入土壤的有機碳量在0—10 cm、10—20 cm、20—40 cm土層分別增加了55.08、89.27、39.67 g m-3a-1,初步證實了我們前期的推測。Bj?rk等[44]在研究苔原植物對升溫的響應時認為,由于升溫導致細根生物量的提高和形態的改變,造成更多根源碳輸入土壤。值得注意的是,在本研究中,盡管升溫未導致0—10 cm土層細根周轉速率發生顯著變化,但由于參與周轉的細根生物量顯著增加,該層的根源有機碳輸入也同樣增加。
根據Wang等[58]的預測,在RCP4.5或RCP8.5氣候變化情境下,盡管總面積可能會減少,但由于紅樹林濕地中溫度和降水的增加,以及鹽沼濕地中相對海平面的升高,濱海濕地碳的凈積累將持續增加。然而,該研究采用的沉積物斷代技術(sediment dating techniques)無法確定植物地下部分生產量增加導致的碳積累變化。本研究通過對模擬升溫條件下蘆葦濕地細根生長和形態指標的連續監測,則為了解氣候變化條件下濱海濕地植物根系的變化,及其對濱海濕地碳循環的影響提供了新的依據。本研究結果表明,對于水文條件良好、生態系統健康的濱海蘆葦濕地,氣候變暖可能會在一定程度上促進表層土壤的根源有機碳輸入,從而導致濕地土壤碳儲量的增加。另外,除根系周轉外,植物根系還能將分配到根部的有機、無機化合物釋放到周圍土壤形成根際沉積,其中根系分泌物是最重要組分[59]。后續研究將結合根系形態、生長和分泌物指標測定,進一步探討升溫條件下蘆葦濕地細根對土壤碳平衡的影響機制。
4 結論
在長江口蘆葦濕地中,通過一個生長季的連續監測和分析發現,模擬溫度升高顯著增加了0—40 cm細根(≤2 mm)的總量(總根長、總表面積、生物量),而對反映細根水分與養分吸收效率的指標(比根長、比表面積、直徑)無顯著影響。細根總量的增加主要源自0—20 cm土層細根指標的變化,20—40 cm土層的貢獻相對較小。升溫顯著提高了細根凈生長速率,增強了其“M”型曲線的生長節律,而這種促進作用隨土層加深而逐漸減弱。升溫顯著增加了10—40 cm土層的細根周轉速率,而在0—10 cm土層影響不顯著。升溫使細根周轉速率加快以及參與周轉的細根總量增加,導致各土層特別是0—20 cm土層根源有機碳輸入顯著增加,這可能會深刻影響蘆葦濕地的土壤碳平衡。因此,未來在開展濱海濕地土壤碳平衡的模擬和預測時,應當充分考慮升溫條件下植物細根相關碳過程的可能變化。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
