桂林巖溶石山密花樹群落主要物種的種間關(guān)聯(lián)及群落穩(wěn)定性
2022-05-26 08:32:30涂洪潤農(nóng)娟麗趙佳佳楊婉琳朱琪琪謝彥軍劉潤紅
生態(tài)學(xué)報 2022年9期
關(guān)鍵詞:關(guān)聯(lián)物種
涂洪潤,農(nóng)娟麗,朱 軍,趙佳佳,楊婉琳,朱琪琪,謝彥軍,劉潤紅
1 柳州鐵道職業(yè)技術(shù)學(xué)院建筑技術(shù)學(xué)院,柳州 545616 2 廣西師范大學(xué)生命科學(xué)學(xué)院,桂林 541006 3 貴州省氣候中心,貴陽 550002 4 河池學(xué)院化學(xué)與生物工程學(xué)院,宜州 546300 5 廣西大學(xué)林學(xué)院,南寧 530004
植物群落是由共存物種共同組成的有機結(jié)合體,群落內(nèi)各物種之間并不是孤立存在的,而是彼此聯(lián)系、相互依存、相互促進、相互制約、協(xié)同進化并與生境相互作用,使得群落處于相對穩(wěn)定的狀態(tài)[1—3]。種間關(guān)聯(lián)是群落內(nèi)各物種在生境中相互作用和相互影響所形成的有機聯(lián)系,其是各物種自身演化到群落穩(wěn)定與否的表現(xiàn)形式[2—5]。而群落穩(wěn)定性作為群落結(jié)構(gòu)與功能的綜合性特征,是外部條件發(fā)生變化或存在擾動時群落維持自身穩(wěn)定性的能力[6—7]。因此,綜合研究群落內(nèi)主要物種的種間關(guān)聯(lián)及群落穩(wěn)定性,有助于全面地認(rèn)識群落的類型、結(jié)構(gòu)、演替趨勢以及生物多樣性維持機制[8],進而為植被恢復(fù)與重建、森林經(jīng)營與管理以及生物多樣性保護等提供理論基礎(chǔ)[9—10]。
近年來,國內(nèi)外學(xué)者對群落內(nèi)主要物種的種間關(guān)聯(lián)及群落穩(wěn)定性進行了許多卓有成效的研究,結(jié)果表明種間關(guān)聯(lián)性與群落穩(wěn)定性密切相關(guān)[11—13]。一般而言,群落內(nèi)各物種間正關(guān)聯(lián)程度越高,群落越穩(wěn)定;負(fù)關(guān)聯(lián)程度越高,物種間相對獨立,群落越不穩(wěn)定[14—15]。隨著群落由穩(wěn)定性較差的先鋒群落正向演替為穩(wěn)定性較高的頂極群落,群落內(nèi)各物種間的正關(guān)聯(lián)關(guān)系逐漸增強,以求得物種間的穩(wěn)定共存[16—20],在植被的恢復(fù)過程中也存在類似的規(guī)律[21—23]。相反,在遭受生物入侵和退化的生態(tài)系統(tǒng)中,群落內(nèi)各物種間的正關(guān)聯(lián)關(guān)系減弱,負(fù)關(guān)聯(lián)關(guān)系增強[2,24—27]。此外,處于相對穩(wěn)定環(huán)境條件下的群落內(nèi)各物種間的正關(guān)聯(lián)關(guān)系更強,反之,處于多變環(huán)境條件或經(jīng)常遭受較強干擾的群落,各物種間的正關(guān)聯(lián)關(guān)系較弱,負(fù)關(guān)聯(lián)關(guān)系較強[28—29]。然而,上述研究結(jié)論會因所研究的生態(tài)系統(tǒng)類型、群落類型、群落所處的演替階段、取樣面積、樣方數(shù)量、物種數(shù)量以及時間和空間尺度等諸多因素而異[30],未來仍需要從以上方面進行深入研究,以揭示其普適性規(guī)律。此外,在先前的絕大部分研究中,群落內(nèi)主要物種的種間關(guān)聯(lián)研究的研究對象主要側(cè)重于群落某一垂直結(jié)構(gòu)層次中的物種之間[31—33],整個群落的優(yōu)勢物種之間[34—36]以及群落內(nèi)某一珍稀瀕危物種或外來入侵物種與共存物種之間[24,37—39]。與之相比,關(guān)于群落內(nèi)不同垂直層次(喬木層、灌木層和草本層)內(nèi)優(yōu)勢物種之間的種間關(guān)聯(lián)研究相對較少,尤其是針對桂林巖溶石山次生演替早期階段植物群落內(nèi)主要物種的種間關(guān)聯(lián)及其群落穩(wěn)定性的研究尚鮮見報道。
作為桂林巖溶石山地區(qū)次生演替早期階段典型群落之一的密花樹(Myrsineseguinii)群落,由于其特殊的石灰?guī)r地質(zhì)地貌背景和復(fù)雜多樣的生境類型,形成了豐富的物種組成、特殊的區(qū)系成分和復(fù)雜的群落結(jié)構(gòu),為研究植物種間關(guān)聯(lián)及群落穩(wěn)定性提供了良好的研究對象。但是,前人關(guān)于密花樹的研究主要集中在種子萌發(fā)以及移栽技術(shù)等方面[40],而關(guān)于其群落內(nèi)主要物種的種間關(guān)聯(lián)特征及群落穩(wěn)定性尚未見報道。桂林巖溶石山密花樹群落主要物種的種間關(guān)聯(lián)特征和群落穩(wěn)定性如何?該群落主要物種的種間關(guān)聯(lián)特征與群落穩(wěn)定性有何關(guān)系?這些都有待揭示。對處于次生演替早期的桂林巖溶石山密花樹群落而言,其群落內(nèi)各物種生態(tài)位重疊嚴(yán)重,對資源競爭激烈,尤其在垂直結(jié)構(gòu)上對光照存在強烈競爭,因而此時群落內(nèi)物種共存尚不穩(wěn)定。基于此假設(shè):(1)桂林巖溶石山密花樹群落主要物種的種間關(guān)系松散,表現(xiàn)出較低的種間關(guān)聯(lián)程度,各物種多呈獨立分布格局;(2)該群落處于不穩(wěn)定狀態(tài);(3)種間關(guān)聯(lián)性與群落穩(wěn)定性密切關(guān)聯(lián),群落內(nèi)各物種間負(fù)關(guān)聯(lián)程度越高,物種間相對獨立,群落越不穩(wěn)定。為了檢驗以上科學(xué)假說,本研究以桂林巖溶石山密花樹群落為研究對象,采用方差比率法(VR)、基于2×2列聯(lián)表的χ2檢驗、Pearson相關(guān)檢驗、Spearman秩相關(guān)檢驗以及M.Godron穩(wěn)定性測定等數(shù)量分析方法,對密花樹群落內(nèi)主要物種的種間關(guān)聯(lián)特征及其群落穩(wěn)定性進行分析,旨在深入認(rèn)識該群落的結(jié)構(gòu)特征及其演替趨勢,進而為該地區(qū)的植被恢復(fù)與重建、森林經(jīng)營與管理以及生物多樣性保護等提供理論基礎(chǔ)。
1 材料與方法
1.1 研究區(qū)概況
研究地點位于桂林市靈川縣大圩鎮(zhèn)桐子園村,地理坐標(biāo)為110°22′E,25°13′N,海拔約為132—220 m。該地區(qū)地表形態(tài)復(fù)雜多樣,巖溶地貌分布廣泛且發(fā)育典型,地貌類型主要是由碳酸鹽巖溶蝕作用為主形成的峰叢洼地和峰林谷地,峰林峰叢密集,基巖裸露,地形復(fù)雜而破碎。研究區(qū)位于典型的中亞熱帶濕潤季風(fēng)氣候區(qū),氣候溫和,雨量充沛,無霜期長,光照充足,熱量豐富,夏長冬短,四季分明且雨熱基本同期。年平均氣溫為18—19℃,其中,最冷月(1月)和最熱月(7月)平均氣溫分別為7.9℃和28℃;年降水量為1814—1941 mm,降水量年內(nèi)分配不均,春夏濕潤多雨(多集中在4—7月,占全年的60%,秋冬季干燥少雨,年平均蒸發(fā)量約為1490—1905 mm,年平均相對濕度為73%—79%[41]。全年光照充足,年日照時數(shù)為1670 h,全年無霜期約309 d,風(fēng)向以偏北風(fēng)為主,平均風(fēng)速為2.2—2.7 m/s,平均氣壓為99.49 kPa。該區(qū)域的基巖主要為石灰?guī)r,土壤類型以黃棕色或黑色石灰土為主,具有巖石裸露率高,成土速度慢,土層淺薄且分布不連續(xù),厚度分布不均勻,鈣鎂含量高,保水保肥能力差等特征。密花樹群落層次結(jié)構(gòu)相對簡單,成層現(xiàn)象明顯,可明顯劃分為喬木層、灌木層和草本層等層次。其中,喬木層物種較少,主要物種有密花樹(Myrsineseguinii)、化香樹(Platycaryastrobilacea)、光蠟樹(Fraxinusgriffithii)、麻櫟(Quercusacutissima)、雞仔木(Sinoadinaracemosa)、薄葉潤楠(Machilusleptophylla)、陰香(Cinnamomumburmanii)等;灌木層物種組成較豐富,主要物種有密花樹(Myrsineseguinii)、檵木(Loropetalumchinense)、杜鵑(Rhododendronsimsii)、刺葉冬青(Ilexbioritsensis)、紅背山麻桿(Alchorneatrewioides)和柃木(Euryajaponica),以及化香樹和陰香等喬木幼樹;草本層物種較少,蓋度較低,主要物種有闊鱗鱗毛蕨(Dryopterischampionii)、褐果薹草(Carexbrunnea)、山麥冬(Liriopespicata)、刺齒半邊旗(Pterisdispar)和刺頭復(fù)葉耳蕨(Arachniodesaristata),以及化香樹和密花樹等喬、灌木幼苗。
1.2 群落學(xué)調(diào)查
于2020年7—8月,在全面踏查的基礎(chǔ)上,根據(jù)密花樹群落的類型、結(jié)構(gòu)以及演替的現(xiàn)狀與趨勢等特點,在桂林市靈川縣大圩鎮(zhèn)桐子園村選取受人為干擾較輕的密花樹群落,建立了30個大小為20 m×20 m的樣方,各樣方的基本情況如表1所示。其中,喬木層樣方大小為20 m×20 m,在此基礎(chǔ)上,在每個樣方的四角和中心各設(shè)置1個5 m×5 m的灌木層樣方,并在樣方內(nèi)隨機設(shè)置5個1 m×1 m的草本層樣方。對喬木層調(diào)查樣方中所有DBH ≥ 1 cm喬木植株進行調(diào)查,記錄種名、相對坐標(biāo)、胸徑、高度、枝下高、冠幅和生長狀況等指標(biāo);灌木層調(diào)查對象包括灌木物種以及未滿足喬木層測量標(biāo)準(zhǔn)的更新幼樹,記錄種名、相對坐標(biāo)、高度、基徑和冠幅等指標(biāo);草本層記錄種名、株數(shù)、平均高度和蓋度等指標(biāo)。同時記錄每個樣方的經(jīng)緯度、海拔、坡向、坡位、坡度、土壤類型、土壤厚度、巖石裸露率和干擾程度等生境特征信息。
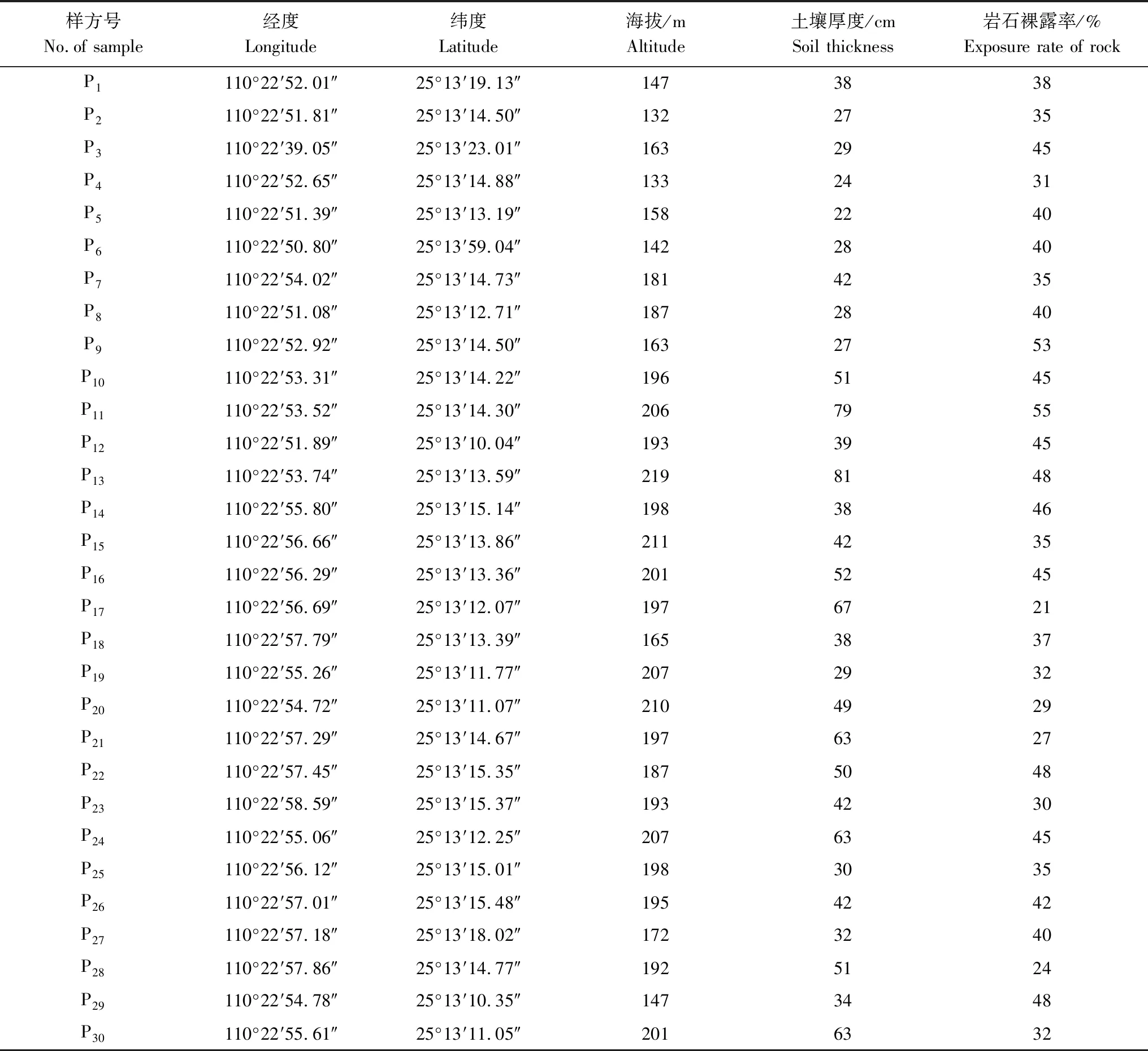
表1 各樣方基本情況Table 1 Basic information of habitat in different plots
1.3 統(tǒng)計與分析
1.3.1重要值計算
根據(jù)30個樣方的統(tǒng)計資料,共記錄有145種植物,其中,喬木層植物31種,灌木層植物69種,草本層植物45種。在數(shù)據(jù)分析過程中考慮到物種過多和計算需要,本研究選取對群落結(jié)構(gòu)和群落環(huán)境的形成具有較強影響的主要物種作為研究對象。為客觀反映各物種在群落中的地位和作用,用重要值(IV)代表各物種的優(yōu)勢度,分喬木層、灌木層和草本層計算重要值,計算公式如下[42]:
喬木層物種重要值(%)=(相對多度+相對頻度+相對顯著度)/3
灌木層和草本層物種重要值(%)=(相對多度+相對頻度)/2
相對多度(RA)=(某一物種的多度/層片中所有物種多度總和)×100%
相對頻度(RF)=(某一物種出現(xiàn)的頻度/層片中所有物種頻度總和)×100%
相對顯著度(RD)=(某一物種的胸高斷面積/層片中所有物種的胸高斷面積總和)×100%
由于各層次物種重要值計算公式的差異,本研究分層選取喬木層、灌木層和草本層中重要值較高的16、20和20種植物(表2)進行種間關(guān)聯(lián)分析。
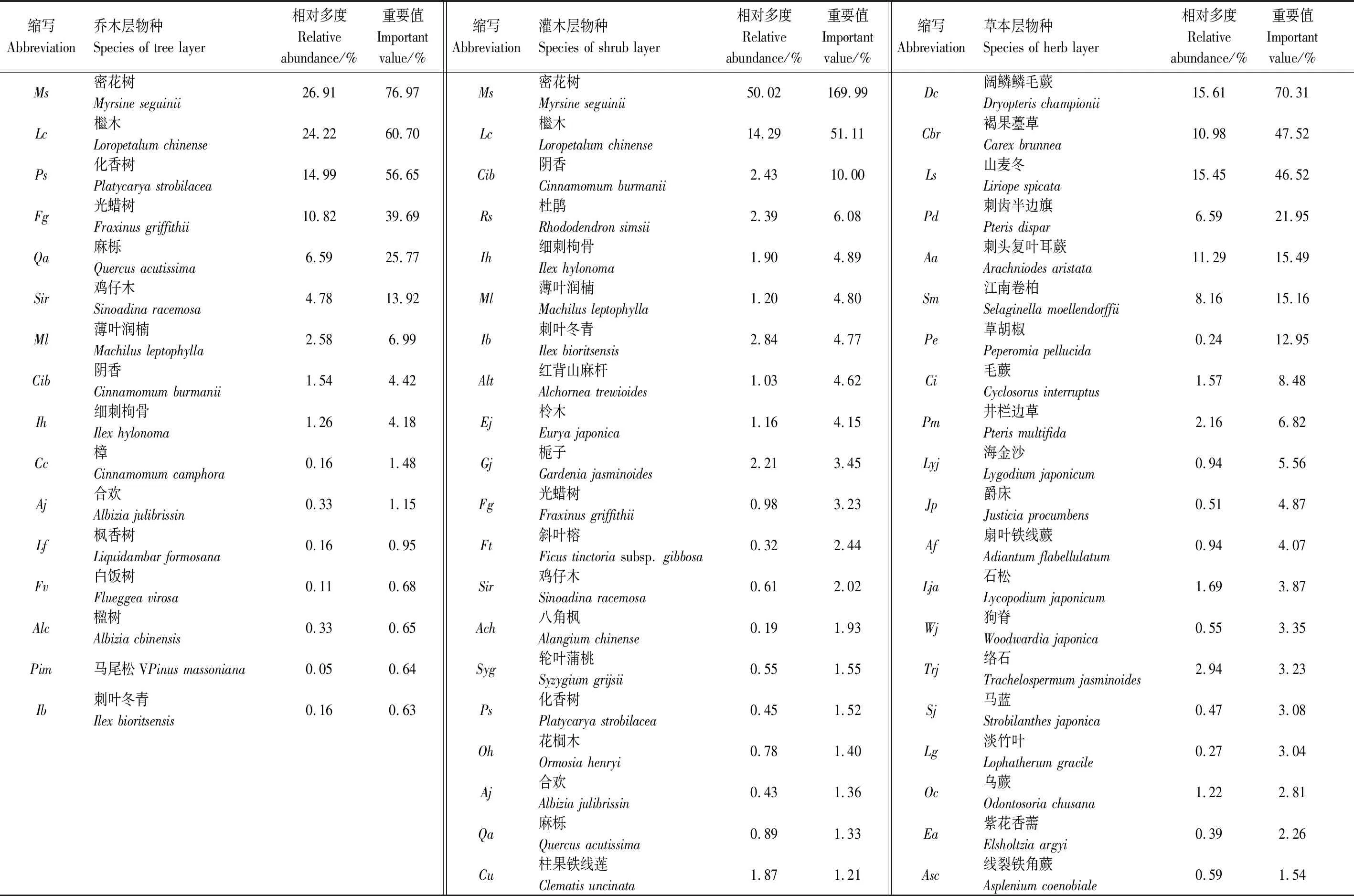
表2 喬木層、灌木層和草本層主要物種及其重要值Table 2 The main species and their important value in the tree layer,shrub layer and herb layer of Myrsine seguinii community in Karst hills of Guilin,southwest China
1.3.2總體關(guān)聯(lián)性檢驗
以物種在樣方內(nèi)是否出現(xiàn)的二元數(shù)據(jù)矩陣作為原始數(shù)據(jù),采用Schluter[43]提出的方差比率法(VR)測定該群落各層次主要物種間的總體關(guān)聯(lián)性,并通過統(tǒng)計量W檢驗總體關(guān)聯(lián)性是否顯著,計算公式如下:
Pi=ni/N
W=VR×N

1.3.3種間關(guān)聯(lián)性分析
采用基于2×2列聯(lián)表的χ2統(tǒng)計量對種間關(guān)聯(lián)性進行定性研究,以確定實測值與期望值之間偏差的顯著程度。在進行χ2檢驗前,首先需根據(jù)物種在30個樣方中是否存在,將原始數(shù)據(jù)轉(zhuǎn)換成0或1形式的二元數(shù)據(jù)矩陣,0表示物種在樣方中不存在,1表示物種在樣方中存在。然后根據(jù)上述二元數(shù)據(jù)矩陣,將主要種群所有種對的定性數(shù)據(jù)列入2×2聯(lián)列表,計算a、b、c和d的值。由于本研究的取樣方法為非連續(xù)性取樣,因此非連續(xù)性數(shù)據(jù)χ2統(tǒng)計量需采用Yates的連續(xù)校正公式進行計算[45],其計算公式如下:
式中,N為樣方總數(shù),a為2個物種同時出現(xiàn)的樣方數(shù),b和c為2個物種單獨出現(xiàn)的樣方數(shù),d為2個物種均未出現(xiàn)的樣方數(shù)。種間關(guān)聯(lián)性通常分為正關(guān)聯(lián)和負(fù)關(guān)聯(lián)2類,設(shè)V=((a+d)-(b+c))/(a+b+c+d),當(dāng)ad>bc時,則V>0,種間關(guān)聯(lián)性為正關(guān)聯(lián);當(dāng)ad
1.3.4種間相關(guān)性測定
χ2統(tǒng)計量僅能定性描述物種間的關(guān)聯(lián)性是否顯著,并不意味著那些χ2檢驗不顯著的種對間沒有關(guān)聯(lián)性,且其無法區(qū)分物種間的關(guān)聯(lián)性強弱,故在一定程度上模糊了種間關(guān)聯(lián)性之間的差異性[28]。與之相比,Pearson相關(guān)和Spearman秩相關(guān)系數(shù)檢驗則是基于數(shù)量數(shù)據(jù),能夠比較客觀地反映物種間的線性關(guān)系,且較其他相關(guān)系數(shù)檢驗更靈敏,可以有效地彌補χ2檢驗的不足[30]。因此,本研究利用各層次主要物種的重要值作為Pearson相關(guān)和Spearman秩相關(guān)檢驗的數(shù)量指標(biāo),對種對間的線性關(guān)系進行定量分析它們同時出現(xiàn)的可能性[45]。
Pearson相關(guān)系數(shù)計算公式如下:
Spearman秩相關(guān)系數(shù)計算公式如下:

1.3.5群落穩(wěn)定性分析
采用鄭元潤[46]改進后的M.Godron貢獻定律法對群落的穩(wěn)定性進行分析。首先,將群落中所選取的主要物種按照頻度由大到小的順序排列,換算成相對頻度后逐步累加,得到其累積相對頻度;再將樣地內(nèi)出現(xiàn)的物種總數(shù)取倒數(shù),按照上述植物排序逐個累加,累加結(jié)果乘以100%得到物種累積倒數(shù)百分比。然后,以群落內(nèi)物種累積倒數(shù)百分比為橫坐標(biāo)(x),物種累積相對頻度為縱坐標(biāo)(y),建立模糊散點平滑曲線模型。另外,做一條經(jīng)過(0,100)、(100,0)的直線,該直線與平滑曲線的交點即為所求的群落穩(wěn)定比值。其計算公式如下:
平滑曲線模擬方程:y=ax2+bx+c
直線方程:y=100-x
將直線方程代入平滑曲線模擬方程:ax2+(b+1)x+c-100=0
式中,x為群落內(nèi)物種累積倒數(shù)百分比,y為物種累積相對頻度,a、b和c為回歸曲線模型參數(shù)。該方程有2個解,本研究取0—100的解(另一個解遠(yuǎn)大于100,屬于無效值)。x/y值和20/80的接近程度體現(xiàn)了群落的穩(wěn)定性,越接近表示穩(wěn)定性越好,反之則穩(wěn)定性越差。此外,僅通過交點坐標(biāo)無法直接反映群落穩(wěn)定性的大小,故本研究采用群落交點坐標(biāo)與穩(wěn)定點坐標(biāo)之間的歐式距離來表示群落穩(wěn)定性的大小。
本研究所有統(tǒng)計分析與制圖均在Excel 2019和R 4.0.4 (http://cran.r-project.org/)中完成。其中,χ2檢驗、Pearson相關(guān)系數(shù)和Spearman秩相關(guān)系數(shù)檢驗分別由R語言中spaa程序包[47]的sp.assoc()、sp.pair()函數(shù)計算得出,Pearson相關(guān)和Spearman秩相關(guān)顯著性檢驗通過R語言中psych程序包的corr.test()函數(shù)完成。
2 結(jié)果與分析
2.1 總體關(guān)聯(lián)性
2.2 主要物種的種間關(guān)聯(lián)性分析
2.2.1喬木層主要物種的種間關(guān)聯(lián)性
喬木層主要物種的χ2檢驗結(jié)果表明(圖1和表4),在所有120個種對中,呈正關(guān)聯(lián)的種對有40對,占總種對數(shù)的33.33%,其中,呈極顯著、顯著和不顯著正關(guān)聯(lián)的種對分別有6、7和27對,分別占總種對數(shù)的5.00%、5.83%和22.50%;呈負(fù)關(guān)聯(lián)種對為72對,占總種對數(shù)的60.00%,其中,呈極顯著、顯著和不顯著負(fù)關(guān)聯(lián)的種對分別有11、12和49對,分別占總種對數(shù)的9.17%、10.00%和40.83%;正負(fù)關(guān)聯(lián)種對的比值為0.56,負(fù)關(guān)聯(lián)種對占優(yōu)勢。不顯著關(guān)聯(lián)和無關(guān)聯(lián)種對有84對,占總種對數(shù)的70.00%,種間關(guān)聯(lián)較松散,各物種間呈獨立分布格局。
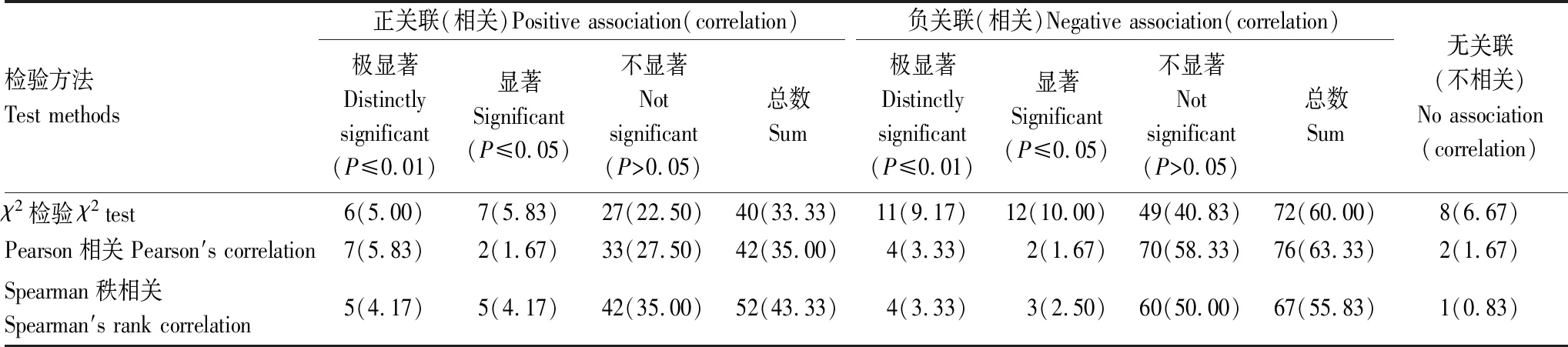
表4 桂林巖溶石山密花樹群落喬木層主要物種的χ2檢驗、Pearson相關(guān)檢驗和Spearman秩相關(guān)檢驗結(jié)果比較Table 4 The comparison of χ2 test,Pearson′ s correlation test and Spearman′ s rank correlation test among main species in tree layer of Myrsine seguinii community in Karst hills of Guilin,southwest China
2.2.2灌木層主要物種的種間關(guān)聯(lián)性
灌木層主要物種的χ2檢驗結(jié)果表明(圖2和表5),在所有190個種對中,呈正關(guān)聯(lián)的種對有57對,占總種對數(shù)的30.00%,其中,呈極顯著、顯著和不顯著正關(guān)聯(lián)的種對數(shù)分別有9、18和30對,分別占總種對數(shù)的4.74%、9.47%和15.79%;呈負(fù)關(guān)聯(lián)種對有124對,占總種對數(shù)的65.26%,其中,呈極顯著、顯著和不顯著負(fù)關(guān)聯(lián)的種對分別有20、32和72對,分別占總種對數(shù)的10.53%、16.84%和37.89%;正負(fù)關(guān)聯(lián)種對的比值為0.46,負(fù)關(guān)聯(lián)種對占優(yōu)勢。不顯著關(guān)聯(lián)和無關(guān)聯(lián)種對有111對,占總種對數(shù)的58.42%,種間關(guān)聯(lián)較松散,各物種呈獨立分布格局。

圖1 桂林巖溶石山密花樹群落喬木層主要物種的χ2檢驗半矩陣圖Fig.1 Semi-matrix diagram of interspecific association χ2 test among main species in tree layer of Myrsine seguinii community in Karst hills of Guilin,southwest ChinaMs:密花樹Myrsine seguinii;Lc:檵木Loropetalum chinense;Ps:化香樹Platycarya strobilacea;Fg:光蠟樹Fraxinus griffithii;Qa:麻櫟Quercus acutissima;Sir:雞仔木Sinoadina racemosa;Ml:薄葉潤楠Machilus leptophylla;Cib:陰香Cinnamomum burmanii;Ih:細(xì)刺枸骨Ilex hylonoma;Cc:樟Cinnamomum camphora;Aj:合歡Albizia julibrissin;Lf:楓香樹Liquidambar formosana;Fv:白飯樹Flueggea virosa;Alc:楹樹Albizia cbinensis;Pim:馬尾松Pinus massoniana;Ib:刺葉冬青,Ilex bioritsensis

圖2 桂林巖溶石山密花樹群落灌木層主要物種的χ2檢驗半矩陣圖Fig.2 Semi-matrix diagram of interspecific association χ2 test among main species in shrub layer of Myrsine seguinii community in Karst hills of Guilin,southwest ChinaMs:密花樹Myrsine seguinii;Lc:檵木Loropetalum chinense;Cib:陰香Cinnamomum burmanii;Rs:杜鵑Rhododendron simsii;Ih:細(xì)刺枸骨Ilex hylonoma;Ml:薄葉潤楠Machilus leptophylla;Ib:刺葉冬青Ilex bioritsensis;Alt:紅背山麻桿Alchornea trewioides;Ej:柃木Eurya japonica;Gj:梔子Gardenia jasminoides;Fg:光蠟樹Fraxinus griffithii;Ft:斜葉榕Ficus tinctoria subsp.gibbosa;Sir:雞仔木Sinoadina racemosa;Ach:八角楓Alangium chinense;Syg:輪葉蒲桃Syzygium grijsii;Ps:化香樹Platycarya strobilacea;Oh:花櫚木Ormosia henryi;Aj:合歡Albizia julibrissin;Qa:麻櫟Quercus acutissima;Cu:柱果鐵線蓮Clematis uncinata

表5 桂林巖溶石山密花樹群落灌木層主要物種的χ2檢驗、Pearson相關(guān)檢驗和Spearman秩相關(guān)檢驗結(jié)果比較Table 5 The comparison of χ2 test,Pearson′ s correlation test and Spearman′ s rank correlation test among main species in shrub layer of Myrsine seguinii community in Karst hills of Guilin,southwest China
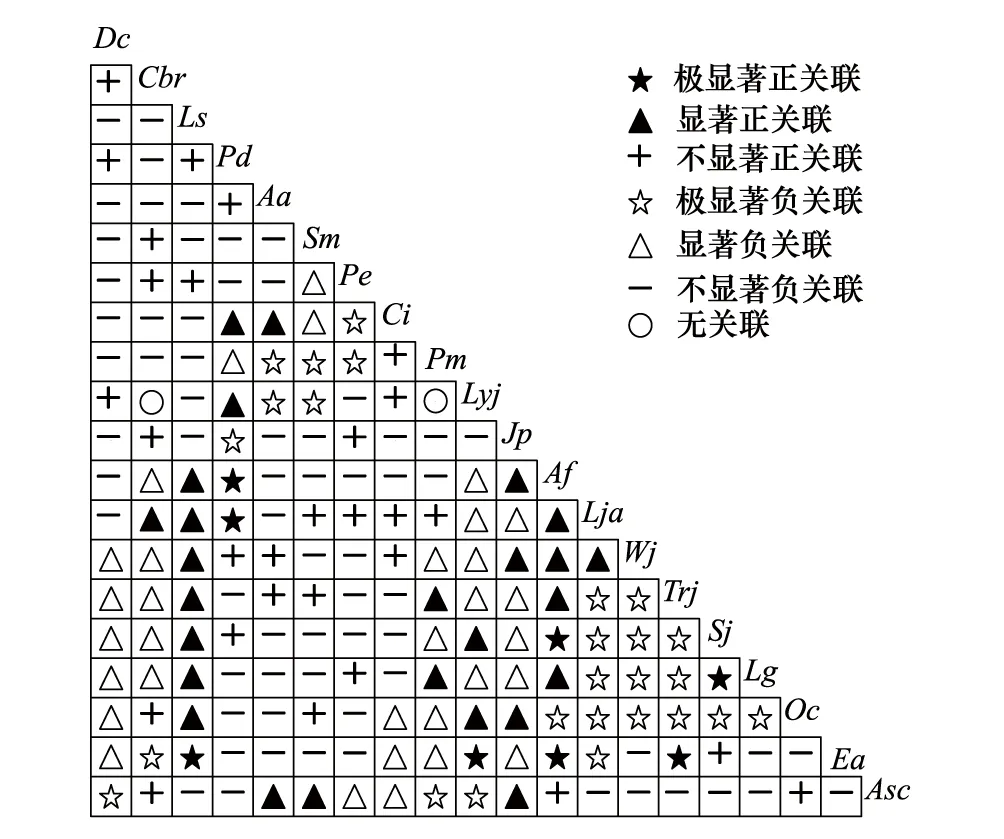
圖3 桂林巖溶石山密花樹群落草本層主要物種的χ2檢驗半矩陣圖Fig.3 Semi-matrix diagram of interspecific association χ2 test among main species in herb layer of Myrsine seguinii community in Karst hills of Guilin,southwest ChinaDc:闊鱗鱗毛蕨Dryopteris championii;Cbr:褐果薹草Carex brunnea;Ls:山麥冬Liriope spicata;Pd:刺齒半邊旗Pteris dispar;Aa:刺頭復(fù)葉耳蕨Arachniodes aristata;Sm:江南卷柏;Selaginella moellendorffii;Pe:草胡椒Peperomia pellucida;Ci:毛蕨Cyclosorus interruptus;Pm:井欄邊草Pteris multifida;Lyj:海金沙Lygodium japonicum;Jpj:爵床Justicia procumbens;Af:扇葉鐵線蕨Adiantum flabellulatum;Lja:石松Lycopodium japonicum;Wj:狗脊Woodwardia japonica;Trj:絡(luò)石Trachelospermum jasminoides;Sj:馬藍(lán)Strobilanthes japonica;Lg:淡竹葉Lophatherum gracile;Oc:烏蕨Odontosoria chusana;Ea:紫花香薷Elsholtzia argyi;Asc:線裂鐵角蕨Asplenium coenobiale
2.2.3草本層主要物種的種間關(guān)聯(lián)性
草本層主要物種的χ2檢驗結(jié)果表明(圖3和表6),在所有190個種對中,呈正關(guān)聯(lián)的種對有63對,占總對數(shù)的33.16%,其中,呈極顯著、顯著和不顯著正關(guān)聯(lián)的種對分別有8、26和29對,分別占總種對數(shù)的4.21%、13.68%和15.26%;呈負(fù)關(guān)聯(lián)種對有125對,占總種對數(shù)的65.79%,其中,呈極顯著、顯著和不顯著負(fù)關(guān)聯(lián)的種對分別有26、32和67對,分別占總種對數(shù)的13.68%、16.84%和35.26%;正負(fù)關(guān)聯(lián)種對的比值為0.50,負(fù)關(guān)聯(lián)種對占優(yōu)勢。不顯著關(guān)聯(lián)和無關(guān)聯(lián)種對有98對,占總種對數(shù)的51.58%,種間關(guān)聯(lián)較松散,各物種呈獨立分布格局。

表6 桂林巖溶石山密花樹群落草本層主要物種的χ2檢驗、Pearson相關(guān)檢驗和Spearman秩相關(guān)檢驗結(jié)果比較Table 6 The comparison of χ2 test,Pearson′ s correlation test and Spearman′ s rank correlation test among main species in herb layer of Myrsine seguinii community in Karst hills of Guilin,southwest China
2.3 主要物種的種間相關(guān)性分析
2.3.1喬木層主要物種的種間相關(guān)性
喬木層主要物種的Pearson相關(guān)檢驗結(jié)果表明(圖4和表4),在所有120個種對中,呈正相關(guān)的種對有42對,占總對數(shù)的35.00%,其中,呈極顯著、顯著和不顯著正相關(guān)的種對數(shù)分別有7、2和33對,分別占總種對數(shù)的5.83%、1.67%和27.50%;呈負(fù)相關(guān)的物種有76對,占總種對數(shù)的63.33%,其中,呈極顯著、顯著和不顯著負(fù)相關(guān)的種對分別有4、2和70對,分別占總種對數(shù)的3.33%、1.67%和58.33%;不顯著相關(guān)和不相關(guān)種對有105對,占總種對數(shù)的87.50%,絕大部分種對為不顯著相關(guān),相關(guān)性不強。
喬木層主要物種的Spearman秩相關(guān)檢驗結(jié)果表明(圖4和表4),在所有120個種對中,呈正相關(guān)的種對有52對,占總種對數(shù)的43.33%,其中,呈極顯著和顯著正相關(guān)的種對均有5對,呈不顯著相關(guān)的種對有42對,分別占總種對數(shù)的4.17%和35.00%;呈負(fù)相關(guān)的種對有67對,占總種對數(shù)的55.83%,其中,呈極顯著、顯著和不顯著負(fù)相關(guān)的種對分別有4、3和60對,分別占總種對數(shù)的3.33%、2.50%和50.00%;不顯著相關(guān)和不相關(guān)種對有103對,占總種對數(shù)的85.83%,絕大部分種對為不顯著相關(guān),相關(guān)性不強。

圖4 喬木層主要物種的Pearson相關(guān)系數(shù)和Spearman秩相關(guān)系數(shù)半矩陣圖Fig.4 Semi-matrix diagram of Pearson′ s correlation coefficients and Spearman′ s rank correlation coefficients among main species in tree layer of Myrsine seguinii community in Karst hills of Guilin,southwest China* P<0.05;** P<0.01;*** P<0.001
2.3.2灌木層主要物種的種間相關(guān)性
灌木層主要物種的Pearson相關(guān)檢驗結(jié)果表明(圖5和表5),在所有190個種對中,呈正相關(guān)的種對有59對,占總種對數(shù)的31.05%,其中,呈極顯著、顯著和不顯著正相關(guān)的種對數(shù)分別有7、7和45對,分別占總種對數(shù)的3.68%、3.68%和23.68%;呈負(fù)相關(guān)的種對有126對,占總種對數(shù)的66.32%,其中,呈極顯著、顯著和不顯著負(fù)相關(guān)的種對分別有3、8和115對,分別占總種對數(shù)的1.58%、4.21%和60.53%;不顯著相關(guān)和不相關(guān)種對有165對,占總種對數(shù)的86.84%,物種的獨立性較強。
灌木層主要物種的Spearsman秩相關(guān)檢驗結(jié)果表明(圖5),在所有190個種對中,呈正相關(guān)的種對有90對,占總種對數(shù)的46.84%,其中,呈極顯著、顯著和不顯著正相關(guān)的種對分別有7、5和77對,分別占總種對數(shù)的3.68%、2.63%和40.53%;呈負(fù)相關(guān)的種對有98對,占總種對數(shù)的51.58%,其中,呈極顯著、顯著和不顯著負(fù)相關(guān)的種對分別有3、9和85對,分別占總種對數(shù)的1.58%、4.74%和45.26%;不顯著相關(guān)和不相關(guān)種對有166對,占總種對數(shù)的87.37%,物種之間的獨立性較強。

圖5 灌木層主要物種的Pearson相關(guān)系數(shù)和Spearman秩相關(guān)系數(shù)半矩陣圖Fig.5 Semi-matrix diagram of Pearson′ s correlation coefficients and Spearman′ s rank correlation coefficients among main species in shrub layer of Myrsine seguinii community in Karst hills of Guilin,southwest China* P<0.05;** P<0.01;*** P<0.001
2.3.3草本層主要物種的種間相關(guān)性
草本層主要物種的Pearson相關(guān)檢驗結(jié)果表明(圖6和表6),在所有190個種對中,呈正相關(guān)的種對有61對,占總種對數(shù)的32.11%,其中,呈極顯著、顯著和不顯著正相關(guān)的種對數(shù)分別有4、4和53對,分別占總種對數(shù)的2.11%、2.11%和27.89%;呈負(fù)相關(guān)的種對有126對,占總種對數(shù)的66.32%,其中,呈顯著和不顯著負(fù)相關(guān)的種對分別有4對和122對,占總種對數(shù)的2.11%和64.21%;不顯著相關(guān)和不相關(guān)種對有178對,占總種對數(shù)的93.68%,絕大部分種對相互獨立。
草本層主要物種的Spearsman秩相關(guān)檢驗結(jié)果表明(圖6和表6),在所有190個種對中,呈正相關(guān)的種對有78對,占總種對數(shù)的41.05%,其中,呈極顯著、顯著和不顯著正相關(guān)的種對分別有3對、2對和73對,分別占總種對數(shù)的1.58%、1.05%和38.42%;呈負(fù)相關(guān)的種對有109對,占總種對數(shù)的57.37%,其中,呈極顯著、顯著和不顯著負(fù)相關(guān)的種對分別有2對、9對和98對,分別占總種對數(shù)的1.05%、4.74%和51.58%;不顯著相關(guān)和不相關(guān)種對有174對,占總種對數(shù)的91.58%,絕大部分種對相互獨立。

圖6 草本層主要物種的Pearson相關(guān)系數(shù)和Spearman秩相關(guān)系數(shù)半矩陣圖Fig.6 Semi-matrix diagram of Pearson′ s correlation coefficients and Spearman′ s rank correlation coefficients among main species in herb layer of Myrsine seguinii community in Karst hills of Guilin,southwest China* P<0.05;** P<0.01;*** P<0.001
2.4 群落穩(wěn)定性分析
群落穩(wěn)定性分析結(jié)果表明(圖7和表7),桂林巖溶石山密花樹群落喬木層、灌木層和草本層以及群落整體的物種累積倒數(shù)百分比與累積相對頻度比值分別為27.93/72.07、30.40/69.60、29.05/70.95和27.52/72.48,均遠(yuǎn)離20/80,表明桂林巖溶石山密花樹群落無論是群落整體,還是群落各垂直層次均處于不穩(wěn)定狀態(tài)。

圖7 桂林巖溶石山密花樹群落各層次群落穩(wěn)定性 Fig.7 The stability of different hierarchical levels of Myrsine seguinii community in Karst hills of Guilin,southwest China

表7 桂林巖溶石山密花樹群落各層次群落穩(wěn)定性分析結(jié)果Table 7 Results of community stability among main species of different hierarchical levels of Myrsine seguinii community in Karst hills of Guilin,southwest China
3 討論
3.1 桂林巖溶石山密花樹群落主要物種的種間關(guān)聯(lián)性
本研究利用χ2檢驗、Pearson相關(guān)系數(shù)檢驗和Spearsman秩相關(guān)系數(shù)檢驗3種方法對桂林巖溶石山密花樹群落主要物種的種間關(guān)聯(lián)性進行分析,發(fā)現(xiàn)該群落喬木層、灌木層和草本層主要物種間的總體關(guān)聯(lián)性均呈顯著負(fù)關(guān)聯(lián),絕大部分種對均呈不顯著關(guān)聯(lián),種間關(guān)聯(lián)較松散,相關(guān)性較弱,各物種呈獨立分布格局。上述研究結(jié)果基本上符合本研究提出的科學(xué)假說(1),也同李帥鋒等[48]、俞筱押等[49]、史浩伯等[50]和陳龍等[51]分別對滇西北云南紅豆杉群落、塔里木河下游植被、貴州茂蘭國家級自然保護區(qū)四藥門花群落和貴州北盤江石漠化區(qū)植物群落進行研究得到的結(jié)論一致,卻同涂洪潤等[52]和劉潤紅等[53]分別研究同處于桂林巖溶石山的桂林巖溶石山頂極群落—青岡群落主要喬木和灌木物種的種間關(guān)聯(lián)所得到的結(jié)論相反,表明桂林巖溶石山密花樹群落正處于動態(tài)演替過程中,群落內(nèi)各層次物種間分布較松散,尚未形成密切聯(lián)系,群落處于不穩(wěn)定狀態(tài)。這可能是由于桂林巖溶石山密花樹群落處于巖溶石山植被生態(tài)系統(tǒng)次生演替的早期階段,群落內(nèi)各垂直層次的物種多以喜陽植物為主[42,54],它們的生物學(xué)特性和生態(tài)習(xí)性一致,對生境的要求較為相似,生態(tài)位互補性不明顯,對環(huán)境資源尤其是光照存在競爭,因而喬木層、灌木層和草本層主要物種間的總體關(guān)聯(lián)性不顯著。與此同時,由于次生演替早期群落內(nèi)的物種組成復(fù)雜,不同葉習(xí)性物種(喜陽和耐陰物種、常綠和落葉物種)共存,某些物種對資源利用存在互補性差異,而另一些物種之間不存在生態(tài)位互補,其種間聯(lián)結(jié)性并不一定呈現(xiàn)出顯著正關(guān)聯(lián)或負(fù)關(guān)聯(lián)的特性,而呈現(xiàn)出許多弱關(guān)聯(lián)或無關(guān)聯(lián)種對共存的現(xiàn)象,故物種間分布較松散,種間關(guān)聯(lián)性較弱。然而,隨著植物群落演替的進行,群落內(nèi)物種組成會呈現(xiàn)出此消彼長的動態(tài)變化,各物種的種間關(guān)系也會隨之發(fā)生變化,因此,在群落演替中后期,群落內(nèi)不同垂直層次物種間將會呈現(xiàn)出何種關(guān)系尚待進一步研究。此外,種間關(guān)聯(lián)性揭示的僅僅是種間和種內(nèi)競爭的結(jié)果或現(xiàn)狀,并不能揭示其背后的生態(tài)學(xué)過程與機制,因此未來應(yīng)在對該群落進行長期定位監(jiān)測的基礎(chǔ)上,綜合運用植物生理學(xué)、分子生態(tài)學(xué)和遺傳學(xué)等學(xué)科的先進方法與技術(shù),深入揭示種間關(guān)聯(lián)性形成的生態(tài)學(xué)機制。
種間關(guān)聯(lián)性是不同物種在空間分布上的相互關(guān)聯(lián)性,它反映了物種之間的相互關(guān)系及其對環(huán)境的適應(yīng)性[3,52]。若種對之間表現(xiàn)為正關(guān)聯(lián),說明它們的資源利用方式、生物學(xué)特征和生態(tài)學(xué)習(xí)性相近,對生境具有相似的生態(tài)適應(yīng)性;而負(fù)關(guān)聯(lián)則表明它們的資源利用方式、生物學(xué)特征和生態(tài)學(xué)習(xí)性不同,對生境具有不同的生態(tài)適應(yīng)性或物種之間存在競爭,具有排斥性[55]。本研究中,喬木層的樟和楓香樹、樟和刺葉冬青、楓香樹和馬尾松、陰香和楓香樹等,灌木層的刺葉冬青和麻櫟、柃木和梔子、杜鵑和柃木等種對呈極顯著或顯著正相關(guān),可能是由于這些物種是巖溶石山地區(qū)的適生植物,對生境適應(yīng)性強,相似的生物學(xué)特性和生境適應(yīng)性使得它們的種間關(guān)系呈現(xiàn)為極顯著或顯著正相關(guān)。與之相反,喬木層的密花樹和檵木、化香樹和光蠟樹、密花樹和陰香,灌木層的檵木和杜鵑、檵木和刺葉冬青等種對的種間關(guān)系呈現(xiàn)為極顯著或顯著負(fù)相關(guān),這可能是由于這些物種的生物學(xué)特性差異較大,對生境具有不同的適應(yīng)性所造成的。
3.2 桂林巖溶石山密花樹群落的群落穩(wěn)定性
本研究利用M.Godron法對密花樹群落的喬木層、灌木層和草本層以及群落整體進行群落穩(wěn)定性分析發(fā)現(xiàn):該群落無論是各垂直層次還是群落整體均處于不穩(wěn)定狀態(tài)。上述結(jié)果基本上符合本研究的科學(xué)假說(2),也同周賽霞等[5]、黃祥童等[8]、張滋芳等[14]和簡小枚等[56]分別研究對開蕨與其伴生物種、狹果秤錘樹群落、重度退化的喀斯特天坑草地群落和矮牡丹生存群落得到的結(jié)論一致,說明該群落尚未發(fā)育到成熟階段,物種組成與群落結(jié)構(gòu)仍處于動態(tài)發(fā)展過程中,群落的結(jié)構(gòu)和功能尚不穩(wěn)定,仍需要漫長的時間才能演替成為頂級群落[8,18]。群落穩(wěn)定性受種內(nèi)和種間競爭、環(huán)境壓力以及人為活動干擾等生物因素和非生物因素的共同影響[57]。本研究發(fā)現(xiàn)密花樹群落穩(wěn)定性較差,這一方面可能是由于該群落處于次生演替的早期階段,群落內(nèi)物種由于對光照等資源的爭奪,各物種的種內(nèi)和種間競爭較強,例如化香樹和光蠟樹等處于群落上層的樹種對土壤養(yǎng)分和水分等資源利用的相似性導(dǎo)致競爭激烈,垂直層次內(nèi)物種種對之間關(guān)聯(lián)性較弱,使得該群落整體上呈顯著負(fù)關(guān)聯(lián),群落穩(wěn)定性較差。再者,這可能與桂林巖溶石山惡劣的生境條件有關(guān),該地區(qū)地形復(fù)雜而破碎,巖石裸露率高,土層淺薄且分布不均,土壤蓄水能力差,水分和養(yǎng)分匱乏,植物為共享稀缺的資源,多傾向于聚集在局部適宜的生境斑塊中,由于它們對土壤水分和養(yǎng)分等資源利用方式的相似性加劇了種內(nèi)和種間競爭,導(dǎo)致群落穩(wěn)定性較差。同時,我們在群落調(diào)查過程中發(fā)現(xiàn)密花樹群落受到一定程度的人為活動干擾,如樵采和祭掃等,改變了物種之間的相互關(guān)系,從而對群落穩(wěn)定性產(chǎn)生不利影響,故需加強對該群落的保護與管理,誘導(dǎo)其進行順向演替更新,進而提高植物群落的穩(wěn)定性。然而,本研究僅僅揭示密花樹群落穩(wěn)定性較差,尚不清楚其背后的生態(tài)學(xué)機制以及各機制的相對貢獻,未來應(yīng)深入研究各生態(tài)因子對該群落穩(wěn)定性的影響及其潛在機制,從而為該地區(qū)的植被恢復(fù)與重建、森林經(jīng)營與管理以及生物多樣性保護等提供理論基礎(chǔ)。
3.3 種間關(guān)聯(lián)性與群落穩(wěn)定性之間的關(guān)系
種間關(guān)聯(lián)性與群落穩(wěn)定性密切關(guān)聯(lián),物種間的總體關(guān)聯(lián)性反映了群落的演替階段及其穩(wěn)定性,而群落穩(wěn)定性則是種間關(guān)聯(lián)性的表現(xiàn)形式[53]。通常認(rèn)為處于演替早期的群落,群落內(nèi)各物種間的關(guān)聯(lián)程度較低,正負(fù)關(guān)聯(lián)比值偏低,群落穩(wěn)定性較差;隨著群落正向演替的進行,群落內(nèi)物種組成和群落結(jié)構(gòu)將逐漸趨于復(fù)雜化,各物種的種內(nèi)和種間競爭強度逐漸減弱,種間關(guān)聯(lián)性也將趨于以正關(guān)聯(lián)為主,正負(fù)關(guān)聯(lián)比值增大,無關(guān)聯(lián)的比例也逐漸增加,使得群落內(nèi)各種群能夠穩(wěn)定共存,群落穩(wěn)定性也會逐漸提高[14,48—49,57]。本研究利用種間關(guān)聯(lián)性與M.Godron法對桂林巖溶石山密花樹群落的種間關(guān)聯(lián)及其群落穩(wěn)定性進行研究,發(fā)現(xiàn)該群落種間關(guān)聯(lián)性與群落穩(wěn)定性密切關(guān)聯(lián),種間關(guān)聯(lián)松散,各物種呈相對獨立分布格局,群落處于不穩(wěn)定的演替早期階段。上述結(jié)果基本上符合本研究的科學(xué)假說(3),也同周賽霞等[5]、張滋芳等[14]和史浩伯等[50]分別對狹果秤錘樹群落、塔里木河下游植被和矮牡丹生存群落進行研究得到的結(jié)論一致。然而,種間關(guān)聯(lián)性和群落穩(wěn)定性會隨演替階段的不同而變化,隨著植被演替的進行,種間關(guān)聯(lián)性與群落穩(wěn)定性之間的關(guān)系會如何變化?這就需要對密花樹群落進行連續(xù)動態(tài)監(jiān)測,因此,未來可以基于長期動態(tài)監(jiān)測樣地或采取空間序列代替時間序列的方法對桂林市巖溶石山密花樹群落的種間關(guān)聯(lián)性與群落穩(wěn)定性進行研究,以揭示該群落在演替過程中種間關(guān)系及其群落穩(wěn)定性的變化規(guī)律及其潛在的生態(tài)學(xué)機制。
此外,本研究發(fā)現(xiàn)種間關(guān)聯(lián)性與M.Godron法測定結(jié)果一致,均表明該群落處于不穩(wěn)定的演替早期階段,說明這2種方法都適用于植物群落穩(wěn)定性的研究,二者得出的結(jié)論能夠相互印證。其中,種間關(guān)聯(lián)測定結(jié)果可以清楚地反映物種之間的關(guān)系,然而卻只能依據(jù)種間關(guān)聯(lián)方向的正負(fù)以及關(guān)聯(lián)強度的大小推測其群落穩(wěn)定性。與之相比,M.Godron法測定結(jié)果僅能反映群落是否穩(wěn)定,無法評估種間關(guān)系。由此可見,種間關(guān)聯(lián)性測定與M.Godron法測定結(jié)果各有優(yōu)缺點,二者結(jié)合可以互相補充和完善。因此,在進行種間關(guān)聯(lián)性測定后,可進一步利用M.Godron法測定其群落穩(wěn)定性,從而全面地認(rèn)識群落內(nèi)各物種的種間關(guān)系及其群落穩(wěn)定性。
3.4 桂林巖溶石山植被恢復(fù)與保護策略
研究桂林巖溶石山密花樹群落主要物種的種間關(guān)聯(lián)及群落穩(wěn)定性旨在全面地認(rèn)識該群落的類型、結(jié)構(gòu)、演替趨勢以及生物多樣性維持機制,從而為該地區(qū)的植被恢復(fù)與重建、森林經(jīng)營與管理以及生物多樣性保護等提供理論依據(jù)。從桂林巖溶石山密花樹群落主要物種的種間關(guān)聯(lián)及群落穩(wěn)定性來看,在桂林巖溶石山植被恢復(fù)與重建實踐過程中,應(yīng)將該生態(tài)系統(tǒng)所處退化階段的天然植被作為參考系,根據(jù)生境條件的差異和各物種間的種間關(guān)系,有選擇性地將該演替階段內(nèi)資源利用方式、生物學(xué)特征和生態(tài)學(xué)習(xí)性相近,對生境要求相似的優(yōu)勢鄉(xiāng)土物種(種間正關(guān)聯(lián)較強的鄉(xiāng)土物種)搭配在一起,防止產(chǎn)生惡性種間競爭。此外,在巖溶石山植被的封育過程中,可采取適當(dāng)?shù)膿嵊胧?調(diào)節(jié)群落內(nèi)各物種之間的種間關(guān)系,加強正關(guān)聯(lián)性較強種對的保護和建立,同時適度伐除或控制同優(yōu)勢種負(fù)關(guān)聯(lián)較強的物種,形成相對穩(wěn)定的種間關(guān)系,誘導(dǎo)植被進行演替更新,從而促進巖溶植被快速恢復(fù),提高群落的生物多樣性和穩(wěn)定性,改善巖溶植被生態(tài)系統(tǒng)多功能,增加其生態(tài)系統(tǒng)多服務(wù)。
4 結(jié)論
綜上所述,桂林巖溶石山密花樹群落內(nèi)各垂直層次主要物種間的總體關(guān)聯(lián)性均呈顯著負(fù)關(guān)聯(lián),絕大部分種對均呈不顯著關(guān)聯(lián),種間關(guān)聯(lián)較松散,種間相關(guān)性較弱,各物種間呈相對獨立分布格局,群落處于不穩(wěn)定的演替早期階段。因此,在巖溶地區(qū)植被恢復(fù)與重建過程中,應(yīng)將資源利用方式、生物學(xué)特征和生態(tài)學(xué)習(xí)性相近,對生境要求相似的優(yōu)勢鄉(xiāng)土物種搭配在一起。此外,在目前封山育林模式的基礎(chǔ)上,可采取適當(dāng)?shù)膿嵊胧?調(diào)節(jié)種間關(guān)系,誘導(dǎo)植被進行正向演替,提高群落穩(wěn)定性。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:30
當(dāng)代陜西(2021年17期)2021-11-06 03:21:36
當(dāng)代陜西(2019年15期)2019-09-02 01:52:00
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
學(xué)苑創(chuàng)造·A版(2018年11期)2018-02-01 06:29:20
讀者(2017年5期)2017-02-15 18:04:18
