鯉trim25多拷貝基因進(jìn)化和表達(dá)調(diào)控初探*
2022-06-15 02:29:20朱優(yōu)秀江炎亮馮建新張瀚元吳碧銀
漁業(yè)科學(xué)進(jìn)展 2022年3期
關(guān)鍵詞:物種
朱優(yōu)秀 江炎亮 張 芹 馮建新 張瀚元 吳碧銀 許 建①
鯉多拷貝基因進(jìn)化和表達(dá)調(diào)控初探*
朱優(yōu)秀1,2江炎亮2張 芹3馮建新3張瀚元2吳碧銀1,2許 建2①
(1. 上海海洋大學(xué) 水產(chǎn)科學(xué)國家級實(shí)驗(yàn)教學(xué)示范中心 上海 201306; 2. 中國水產(chǎn)科學(xué)研究院 農(nóng)業(yè)農(nóng)村部水生動物基因組學(xué)重點(diǎn)實(shí)驗(yàn)室 北京 100141; 3. 河南省水產(chǎn)科學(xué)研究院 河南 鄭州 450044)
天然免疫系統(tǒng)是硬骨魚類抵抗病毒感染的主要防御系統(tǒng),三重基序(tripartite motif, TRIM)蛋白家族作為天然免疫系統(tǒng)的重要組成部分,參與病毒感染的免疫網(wǎng)絡(luò)調(diào)控,其中,TRIM25已被證實(shí)在多種魚類的免疫反應(yīng)中發(fā)揮重要作用。本研究對鯉()基因的16個拷貝進(jìn)行了序列進(jìn)化分析、共線性分析和功能域結(jié)構(gòu)分析,并比較了各拷貝在組織中的表達(dá)和順式調(diào)控位點(diǎn)的差異。序列比對和系統(tǒng)進(jìn)化分析結(jié)果均顯示,位于鯉11和12號染色上、結(jié)構(gòu)完整的TRIM25的2個拷貝與金線鲃()和斑馬魚()的TRIM25蛋白結(jié)構(gòu)高度相似,與鯉科魚類以外的其他物種的結(jié)構(gòu)差異較大。基因共線性結(jié)果顯示,基因上下游基因在不同物種的進(jìn)化過程中相對保守。鯉TRIM25蛋白的結(jié)構(gòu)分析顯示,在鯉TRIM25的16個拷貝中,有6個拷貝具有完整功能結(jié)構(gòu)域,其中,各有5個拷貝在鯉的肝與腦組織中高表達(dá)。在構(gòu)建的表達(dá)數(shù)量性狀基因座(eQTL)調(diào)控網(wǎng)絡(luò)中,在肝和腦組織中分別篩選到5個和17個順式調(diào)控基因表達(dá)的單核苷酸多態(tài)性(SNP)位點(diǎn)。本研究對鯉基因多個拷貝的序列差異進(jìn)行了比較,并對鯉與其他物種TRIM25的序列、進(jìn)化關(guān)系和共線性相似度進(jìn)行了比較,揭示了鯉各拷貝間的結(jié)構(gòu)多樣性和在組織中的表達(dá)情況,篩選出了可能調(diào)控基因表達(dá)的SNP位點(diǎn),為今后研究鯉TRIM25相關(guān)的調(diào)控和抗病研究提供了理論依據(jù)。
鯉;TRIM25;蛋白結(jié)構(gòu);系統(tǒng)進(jìn)化樹;eQTL
天然免疫系統(tǒng)是宿主抵抗病原體感染的第一道防線,其對病原體的直接殺傷和清除作用是通過天然免疫分子和吞噬細(xì)胞實(shí)現(xiàn)的,其中,三重基序(tripartite motif, TRIM)家族蛋白作為具有抗病毒作用和天然免疫調(diào)節(jié)功能的代表性蛋白,成為近年來天然免疫領(lǐng)域的研究熱點(diǎn)。TRIM蛋白是一個結(jié)構(gòu)相對保守的蛋白家族,廣泛存在于無脊椎動物和脊椎動物的細(xì)胞中,其結(jié)構(gòu)包括1個RING鋅指結(jié)構(gòu)域、1~2個B-box結(jié)構(gòu)域、卷曲螺旋結(jié)構(gòu)域(Coiled-coil, CC)以及C端可變結(jié)構(gòu)域,因此,TRIM蛋白又被稱為RBCC蛋白(Micale, 2016)。TRIM蛋白家族作為一類重要的E3泛素連接酶,參與信號轉(zhuǎn)導(dǎo)、細(xì)胞凋亡、蛋白質(zhì)加工和運(yùn)輸以及機(jī)體對病毒的應(yīng)答等重要的生命過程。在人類醫(yī)學(xué)研究中發(fā)現(xiàn),它們的改變與多種疾病的發(fā)生密切相關(guān),如發(fā)育障礙、神經(jīng)退行性疾病、病毒感染和腫瘤(Tocchini, 2015)。
在硬骨魚類中,TRIM蛋白在天然免疫系統(tǒng)中同樣發(fā)揮重要的作用。目前,已在斑馬魚()基因組中鑒定出208個基因,在河豚()基因組中發(fā)現(xiàn)了66個基因(Boudinot, 2011),而人類基因組中包含了75個基因(Versteeg, 2013)。與人類相比硬骨魚類的基因家族進(jìn)化速度更快,具有更高的多樣性。在基因家族中,是非常重要的成員之一。近幾年,在魚類的天然免疫系統(tǒng)研究中發(fā)現(xiàn),TRIM25蛋白在多個重要養(yǎng)殖魚類,如羅非魚()、鯉、大黃魚()、石斑魚()等的抗病毒過程中發(fā)揮了重要作用(Manokaran, 2015)。周真真等(2019)通過實(shí)時熒光定量PCR發(fā)現(xiàn),TRIM25在大黃魚的心臟、腦、外周血、脾臟、皮膚、肌肉、頭腎、腸和肝臟組織中均有表達(dá),其中,在肝臟的表達(dá)水平最高;當(dāng)對大黃魚注射Poly (I:C)后,在外周血、頭腎、肝臟和脾臟中均檢測到基因表達(dá)量顯著上調(diào),且均表現(xiàn)為先上升后下降的趨勢,提示基因在大黃魚抗病毒免疫反應(yīng)中起著重要作用。Jin等(2019)研究發(fā)現(xiàn),斑馬魚TRIM25通過靶向調(diào)控RIG-I蛋白的2CARD和RD結(jié)構(gòu)域的K63泛素化,促進(jìn)斑馬魚對紅點(diǎn)石斑魚()神經(jīng)壞死病毒感染的先天免疫應(yīng)答。Yang等(2016)研究發(fā)現(xiàn),在石斑魚脾臟細(xì)胞中過表達(dá)TRIM25可抑制病毒復(fù)制,提高干擾素信號通路相關(guān)基因的表達(dá)水平。這些研究表明,TRIM25在硬骨魚對病毒感染的先天免疫應(yīng)答中發(fā)揮了關(guān)鍵作用。在Ross團(tuán)隊(duì)開展的鯉抗皰疹病毒QTL研究中,定位了44號染色體上的關(guān)聯(lián)SNP,而基因位于該SNP下游6.5 kb,提示TRIM25在鯉抗病毒免疫應(yīng)答中的作用(Palaiokostas, 2018)。鯉是研究基因組進(jìn)化和開展性狀解析的重要模式物種,由于鯉經(jīng)歷了第4輪全基因組復(fù)制,多倍化的基因組具有較高的環(huán)境適應(yīng)性,基因組中多套拷貝的基因?yàn)樾誀畹恼{(diào)控提供了豐富的變異來源和增加了調(diào)控的靈活性。以為例,鯉基因組中的拷貝數(shù)高達(dá)16個。如何精確定位在抗病和免疫應(yīng)答過程中具體發(fā)揮調(diào)控作用的基因拷貝,闡述基因調(diào)控的遺傳位點(diǎn)和作用機(jī)制,需要進(jìn)一步的精細(xì)分析。考慮到SNP對基因表達(dá)的調(diào)控作用,將SNP信息與基因表達(dá)結(jié)果整合分析成為性狀解析的重要手段。表達(dá)數(shù)量性狀基因座(expression quantitative trait loci, eQTL)分析將每個基因的表達(dá)量作為數(shù)量性狀,研究遺傳突變與基因表達(dá)之間的相關(guān)性,來尋找調(diào)控基因表達(dá)的位點(diǎn)及協(xié)同作用的基因,以此來構(gòu)建基因的調(diào)控網(wǎng)絡(luò)(彭啟迪, 2020)。eQTL分析方法廣泛應(yīng)用于人類醫(yī)學(xué)研究和動植物的性狀解析,通過分析與目的基因相關(guān)的基因組區(qū)域的表達(dá)水平(Guo, 2020; Kordas, 2019; Lowe, 2015; Nicolae, 2010; Walker, 2020),闡明由基因變異引起表型改變的分子機(jī)制。然而,在魚類的性狀解析中,eQTL仍處于起步階段,僅在大西洋鮭()(Aslam, 2020)、極地魚(Jacobs, 2020)等幾個物種中有少量報道。
本研究對鯉基因的多個拷貝與人()、小鼠()、爪蟾()、原雞()以及多種魚類進(jìn)行進(jìn)化分析,分析鯉與幾個主要模式物種的共線性。同時,為比較鯉基因各拷貝之間的差異,對鯉的16個拷貝進(jìn)行結(jié)構(gòu)分析及在多個組織中的表達(dá)研究。通過的eQTL網(wǎng)絡(luò)分析,提取可能調(diào)控基因表達(dá)的SNP位點(diǎn),旨在為今后研究鯉基因的相關(guān)調(diào)控和抗病研究提供參考。
1 材料與方法
1.1 TRIM25蛋白序列的提取和比對
鯉TRIM25蛋白16個拷貝的氨基酸序列取自本課題組鯉全基因組序列和注釋文件(Xu, 2019),人、小鼠、原雞、爪蟾、象鼻鯊()、腔棘魚()、紅鰭東方鲀()、大黃魚、石斑魚、斑馬魚、金線鲃()的TRIM25蛋白序列通過NCBI (https://www.ncbi.nlm.nih.gov/protein/)數(shù)據(jù)庫查詢獲得。利用NCBI數(shù)據(jù)庫中的BLAST (https:// blast.ncbi.nlm.nih.gov/Blast.cgi),對12個物種的TRIM25氨基酸序列進(jìn)行比對,計算各物種與鯉TRIM25蛋白的一致性。
1.2 進(jìn)化樹的繪制、共線性分析和蛋白結(jié)構(gòu)域預(yù)測
利用ClustalW對鯉、人、小鼠、雞、爪蟾、象鼻鯊、腔棘魚、紅鰭東方鲀、斑馬魚、大黃魚、金線鲃、石斑魚的TRIM25氨基酸序列進(jìn)行比對,利用RAxML軟件(V 7.2.8)構(gòu)建進(jìn)化樹,將Bootstrap設(shè)為1000,其他參數(shù)設(shè)為默認(rèn)值。各物種基因的共線性數(shù)據(jù),通過NCBI數(shù)據(jù)庫(https://www.ncbi.nlm. nih.gov/)查詢各物種上下游基因獲得。以鯉TRIM25的16個拷貝的蛋白序列為輸入文件,用在線軟件SMART(http://smart.emblheidelberg.de/)(高進(jìn)等, 2016),預(yù)測鯉TRIM25的16個拷貝的蛋白結(jié)構(gòu)域構(gòu)成。
1.3 trim25在組織中的表達(dá)和eQTL調(diào)控網(wǎng)絡(luò)分析
采集40尾黃河鯉和40尾鏡鯉的肝臟及腦組織,使用RNeasy試劑盒(Qiagen, 上海)提取RNA,并委托測序公司利用HiSeq2000測序平臺進(jìn)行150PE的轉(zhuǎn)錄組測序。使用Bowtie2 v2.3.4.2和Cufflinks 2.2.1軟件對轉(zhuǎn)錄組數(shù)據(jù)進(jìn)行基因表達(dá)(FPKM值)的計算,并提取每個樣本中的基因表達(dá)數(shù)據(jù),采用R軟件包ggplot2繪制箱式圖來展示的基因表達(dá)情況。從已有的鯉SNP變異數(shù)據(jù)集中提取對應(yīng)樣本的SNP分型數(shù)據(jù)和位置信息,結(jié)合基因表達(dá)數(shù)據(jù)和基因位置信息,利用MatrixEqtl軟件進(jìn)行順式eQTL的分析,篩選標(biāo)準(zhǔn)為<10–5。
2 結(jié)果
2.1 鯉TRIM25蛋白的結(jié)構(gòu)分析
蛋白結(jié)構(gòu)預(yù)測顯示,完整的鯉TRIM25蛋白包括4個結(jié)構(gòu)域:RING、B-box、Coiled-coil和PRY/SPRY結(jié)構(gòu)域。其中,HHLG38G0859、HHLG11g0652、HHLG12g0509、HHLG29g0701、HHLG3g0722和HHLG4g0919這6個拷貝有完整的4個結(jié)構(gòu)域;HHLG10g1061缺少PRY/SPRY結(jié)構(gòu)域,HHLG37g0859缺少Coiled-coil結(jié)構(gòu)域,HHLG13g0878、HHLG14g0181、HHLG6g0789和HHLG9g1073只包含RING和B-box結(jié)構(gòu)域。HHLG16g0672只包含RING和Coiled-coil結(jié)構(gòu)域,HHLG13g0902只包含B-box和Coiled-coil結(jié)構(gòu)域,HHLG46g0068和HHLG5g0226只包含RING結(jié)構(gòu)域。
2.2 trim25共線性分析
為了更好地理解基因在進(jìn)化上的保守性,對鯉、人、小鼠、雞、斑馬魚5個物種的基因及其上下游基因進(jìn)行了共線性分析(圖2)。由于鯉的染色體與斑馬魚存在1∶2的映射關(guān)系,優(yōu)先選擇對應(yīng)斑馬魚基因所在染色體的鯉11 12號染色體上的HHLG11g0652和HHLG12g05092這2個拷貝來分析。從共線性圖上可以看出,基因在進(jìn)化上高度保守。是與基因關(guān)聯(lián)性最強(qiáng)的基因,存在于人、小鼠、雞、斑馬魚及鯉5個物種。是與人、小鼠、雞基因關(guān)聯(lián)性較強(qiáng)的另一個基因;而、和這3個基因均存在于斑馬魚與鯉基因上下游。其中,位于鯉12號染色體上的基因上下游的基因分布與斑馬魚完全一致;位于鯉11和12號染色體上的基因的上下游基因構(gòu)成一致,只是上下游位置完全顛倒,這可能是鯉11號染色體的該區(qū)域在進(jìn)化過程中發(fā)生了倒位。
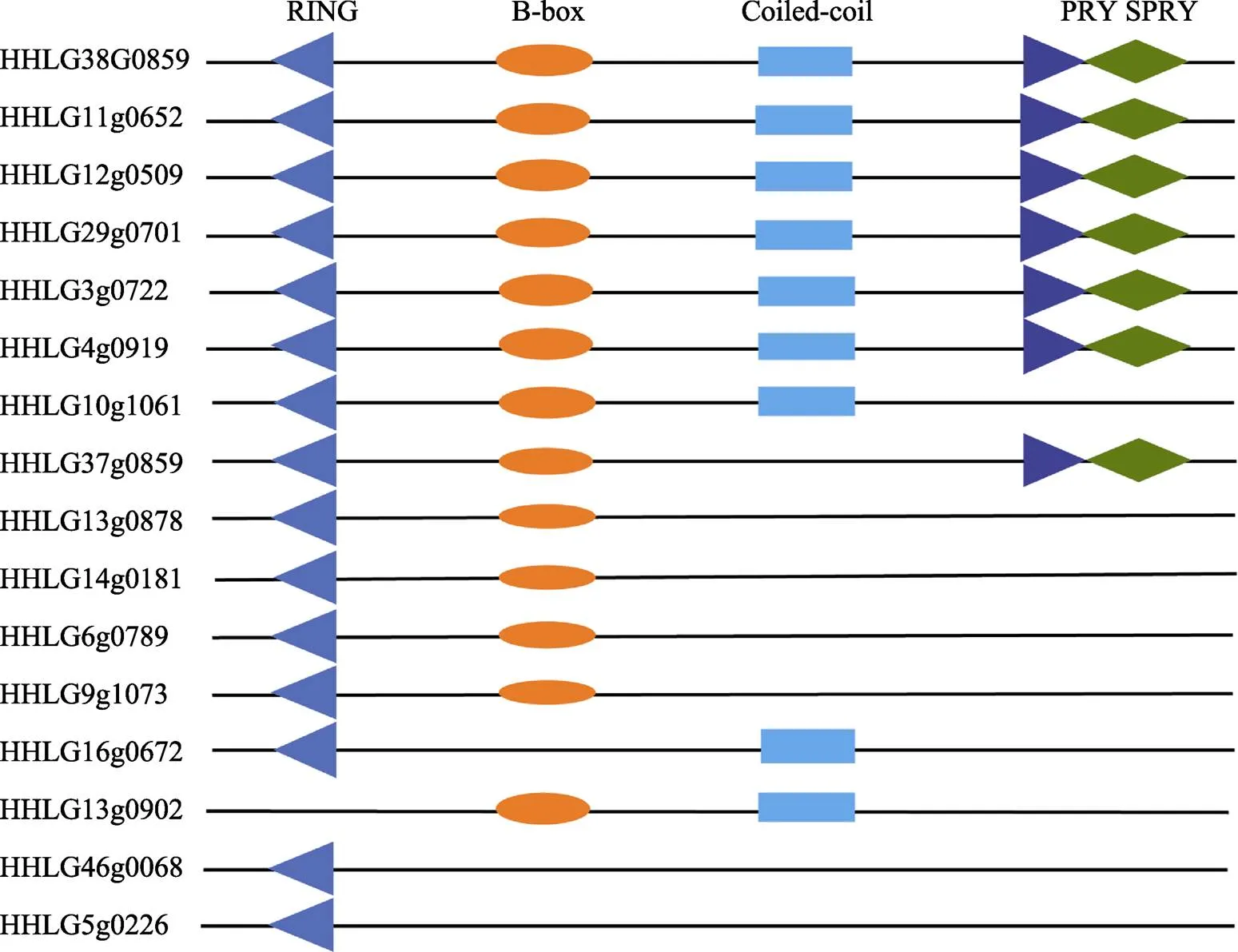
圖1 鯉TRIM25蛋白結(jié)構(gòu)域
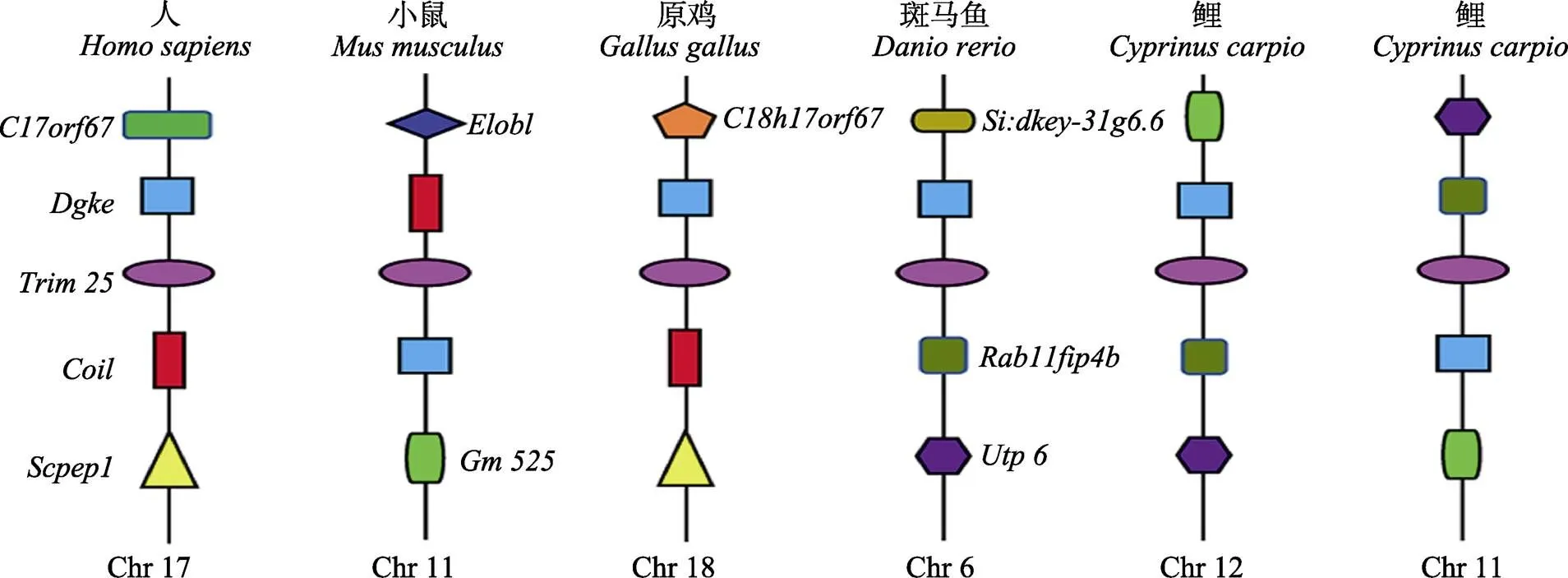
圖2 基因共線性分析
2.3 TRIM25蛋白序列比對
從NCBI數(shù)據(jù)庫中收集代表性物種(哺乳動物、兩棲動物、鳥類、軟骨魚類和硬骨魚類)的TRIM25蛋白氨基酸序列。由于鯉的染色體與斑馬魚存在1 : 2的映射關(guān)系,優(yōu)先選擇對應(yīng)斑馬魚TRIM25所在染色體的鯉的2個拷貝中結(jié)構(gòu)完整、氨基酸序列最長的HHLG12g0509蛋白與其他11個物種TRIM25蛋白進(jìn)行序列比對(圖3)。鯉16個TRIM25拷貝的位置信息見表1。12個物種中,TRIM25蛋白長度從528個氨基酸(爪蟾)到734個氨基酸(石斑魚),與鯉TRIM25蛋白一致性最高的是金線鲃,一致性最低的是爪蟾。
從序列比對的結(jié)果可以看出,哺乳動物中,人與小鼠TRIM25蛋白在RING、B-box、Coiled-coil和PRY/SPRY這4個結(jié)構(gòu)域均高度相似,二者的主要差異在第420~470氨基酸之間。哺乳動物與雞TRIM25蛋白在Coiled-coil和PRY/SPRY結(jié)構(gòu)域高度相似,差異存在于RING和B-box結(jié)構(gòu)域。腔棘魚與象鼻鯊在B-box和Coiled-coil結(jié)構(gòu)域高度相似,爪蟾TRIM25蛋白與其他物種相似性最低。海水魚類中,大黃魚與石斑魚在RING結(jié)構(gòu)域高度相近,紅鰭東方鲀與前二者在RING結(jié)構(gòu)域存在差異;三者的B-box和Coiled- coil結(jié)構(gòu)域高度相似,但三者的PRY/SPRY結(jié)構(gòu)域存在較大差異。鯉科魚類中,斑馬魚、金線鲃和鯉在RING、B-box和Coiled-coil結(jié)構(gòu)域高度相似;鯉與金線鲃的PRY/SPRY結(jié)構(gòu)域高度相似,但斑馬魚的PRY/SPRY結(jié)構(gòu)域與前二者存在差異。
2.4 TRIM25系統(tǒng)進(jìn)化分析
為了解析TRIM25在物種進(jìn)化過程中的差異,構(gòu)建了系統(tǒng)進(jìn)化樹(圖4)。可以看出,人與小鼠的TRIM25最為相近,其次是雞。腔棘魚作為一種原始的硬骨魚類,其TRIM25也與上面3個物種相近。海水魚類(大黃魚、石斑魚、紅鰭東方鲀)單獨(dú)聚為一支;鯉科魚類中,位于鯉11和12號染色體上結(jié)構(gòu)完整的HHLG11g0652、HHLG12g0509拷貝與金線鲃和斑馬魚聚為一支;鯉其余的TRIM25拷貝與象鼻鯊和爪蟾更為相近。鯉異源四倍化過程中產(chǎn)生了大量的多拷貝基因,基因的多個拷貝隨著進(jìn)化的壓力出現(xiàn)了序列和結(jié)構(gòu)的變化,最終導(dǎo)致亞功能化和去功能化。這些發(fā)生較大變異的基因拷貝,可能和其他物種聚到一起。
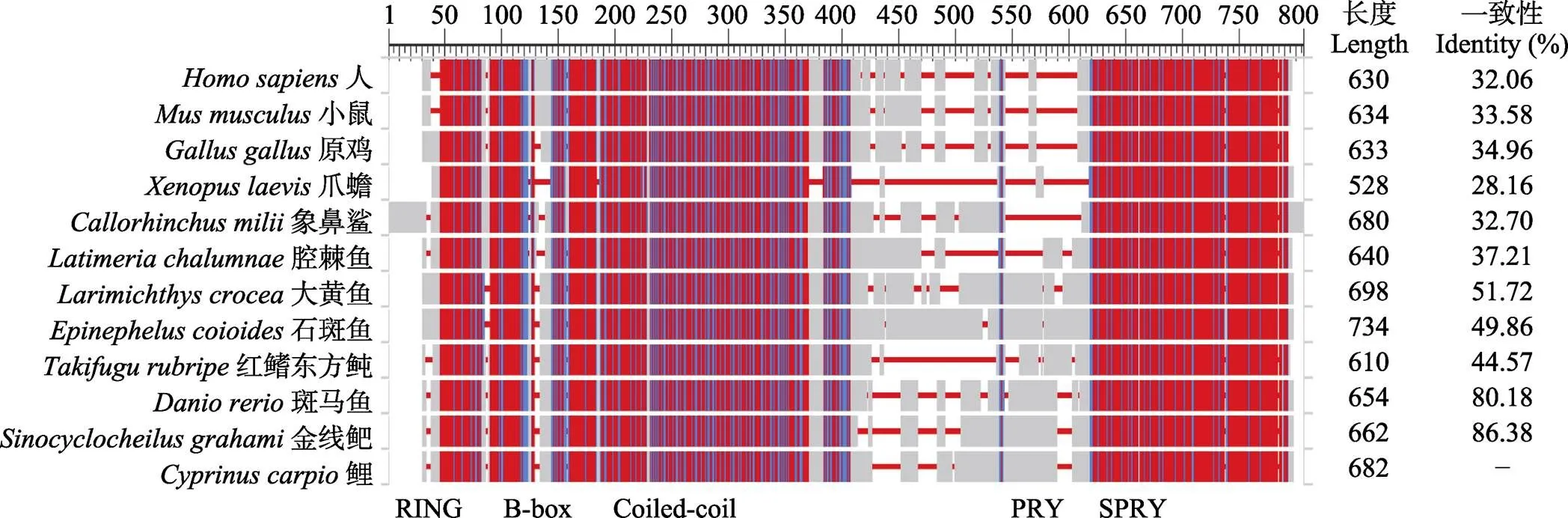
圖3 鯉TRIM25蛋白與其他物種的序列比對
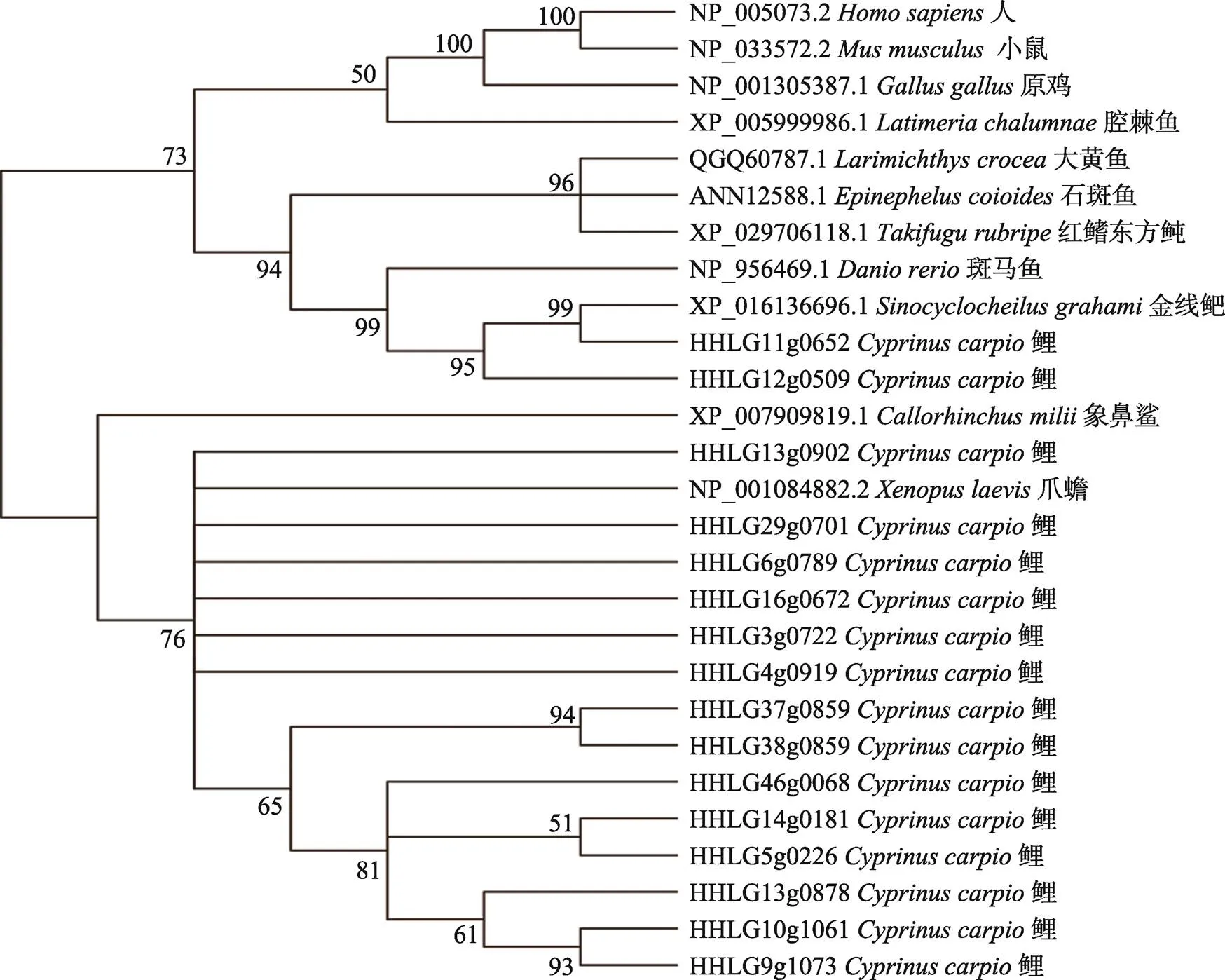
圖4 TRIM25蛋白的系統(tǒng)進(jìn)化樹
表1 鯉基因位置信息
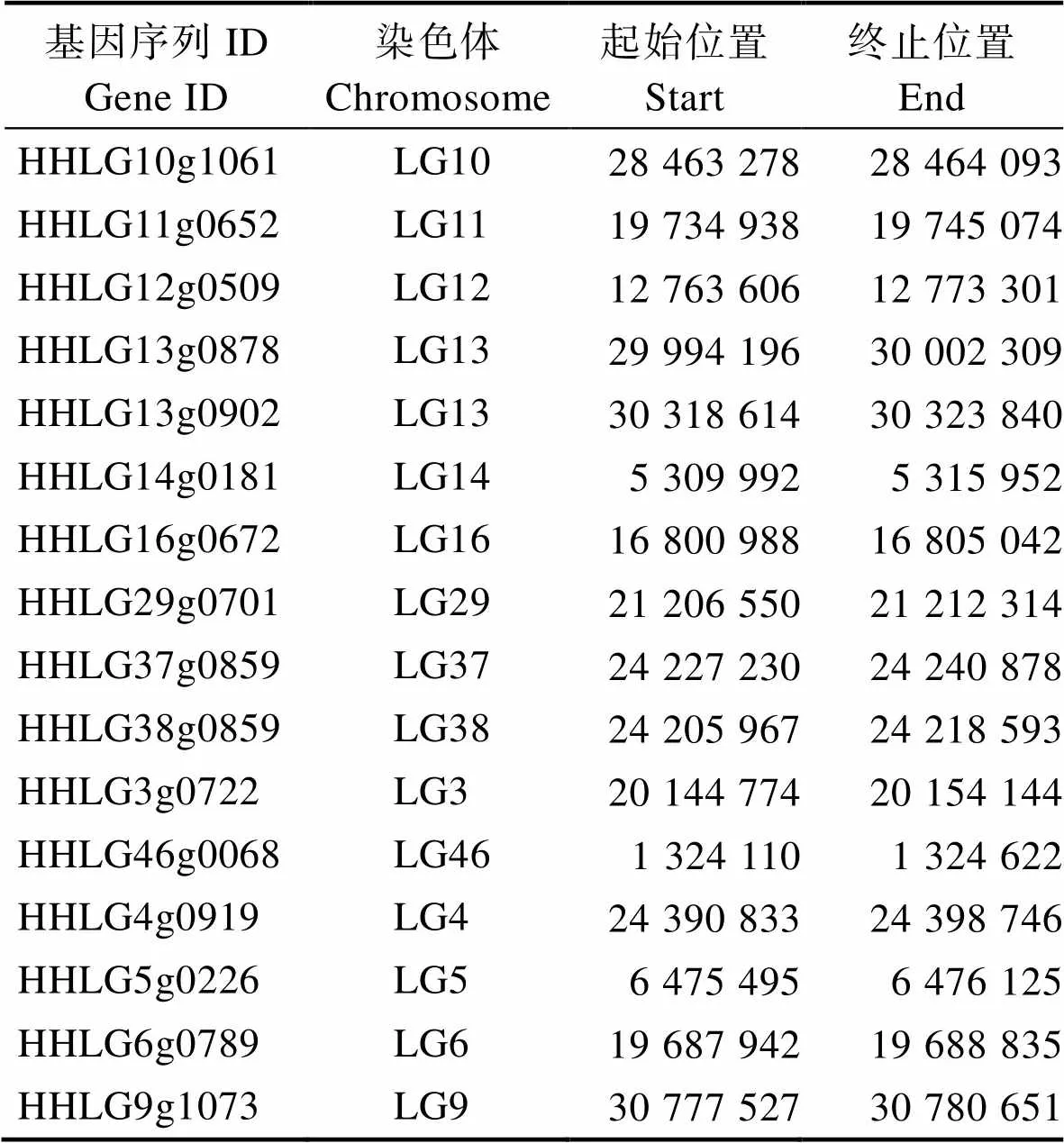
Tab.1 Location information of trim25 gene in C. carpio
2.5 trim25在黃河鯉及鏡鯉肝臟和腦組織中的表達(dá)
在黃河鯉和鏡鯉的肝臟中,表達(dá)量較高的5個拷貝是一致的,包括HHLG10g1061、HHLG11g0652、HHLG12g0509、HHLG13g0878和HHLG16g0672。其他的11個拷貝在黃河鯉和鏡鯉的肝臟組織中表達(dá)量極低或者幾乎不表達(dá)。在黃河鯉和鏡鯉的腦組織中,表達(dá)較高的5個拷貝相同,分別是HHLG10g1061、HHLG11g0652、HHLG12g0509、HHLG13g0878和HHLG3g0722,腦組織和肝臟組織的TRIM25表達(dá)情況非常相似(圖5和圖6)。
2.6 鯉trim25的調(diào)控網(wǎng)絡(luò)分析
從課題組已構(gòu)建的eQTL調(diào)控網(wǎng)絡(luò)中提取出肝臟和腦組織中與基因表達(dá)相關(guān)的順式SNP位點(diǎn)。肝臟中篩選到的陽性SNP位點(diǎn)有5個,位于鯉第4、6、12、13和37號染色體上,分別調(diào)控HHLG4g0919、HHLG6g0789、HHLG12g0509、HHLG13g0878和HHLG37g0859這5個基因。腦組織中篩選到17個陽性位點(diǎn),位于鯉第6、11和13號染色體上,其中,調(diào)控HHLG6g0789基因的有1個SNP位點(diǎn),調(diào)控HHLG11g0652基因的有9個SNP位點(diǎn),調(diào)控HHLG13g0878基因的有6個SNP位點(diǎn),調(diào)控HHLG13g0902的有1個SNP位點(diǎn)(表2和表3)。
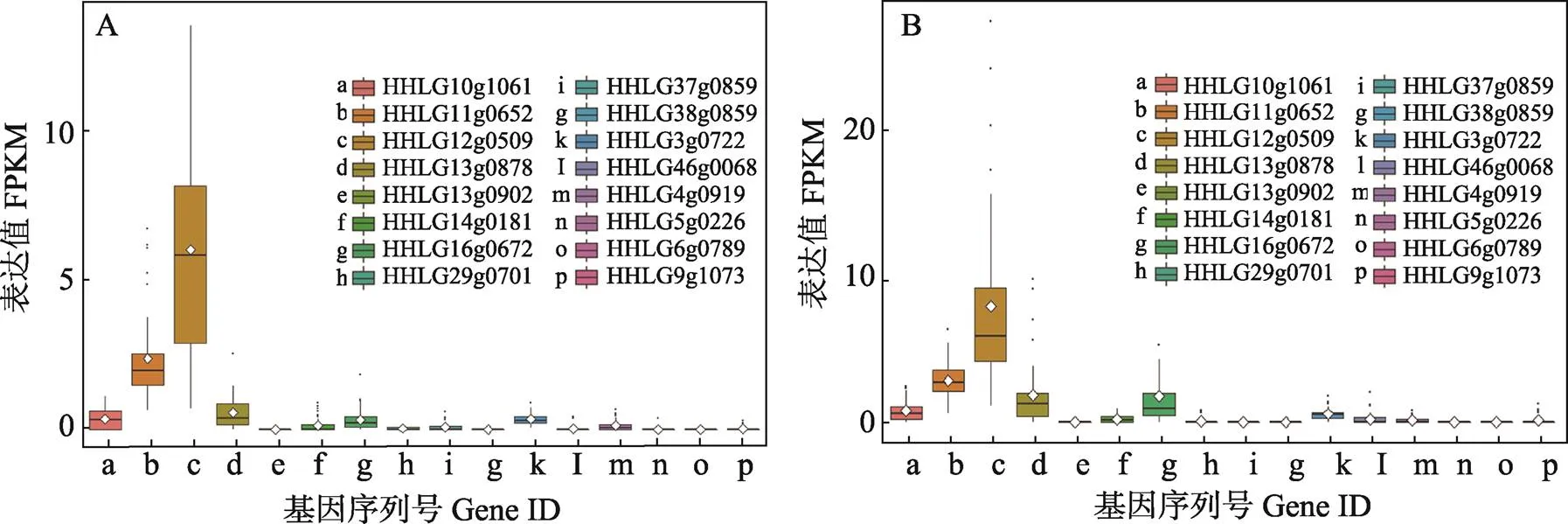
圖5 trim25基因在黃河鯉(A)和鏡鯉(B)肝臟組織中的表達(dá)
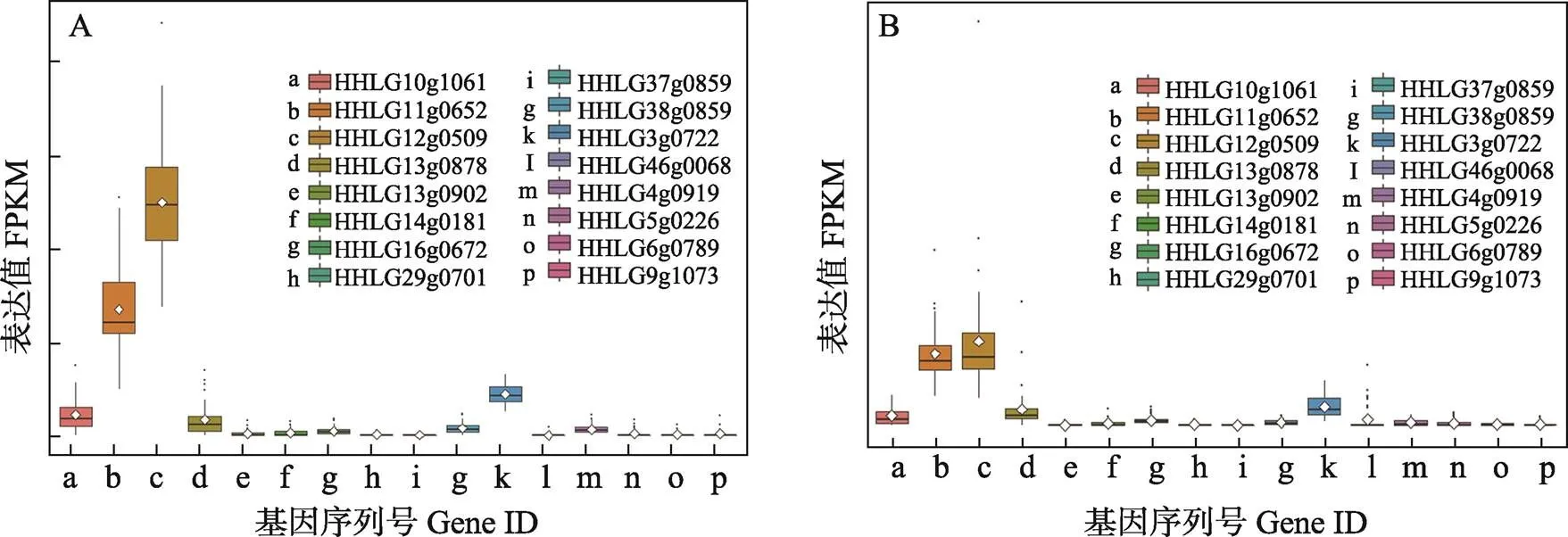
圖6 trim25基因在黃河鯉(A)和鏡鯉(B)腦組織中的表達(dá)
表2 鯉肝臟組織順式調(diào)控基因表達(dá)的SNP
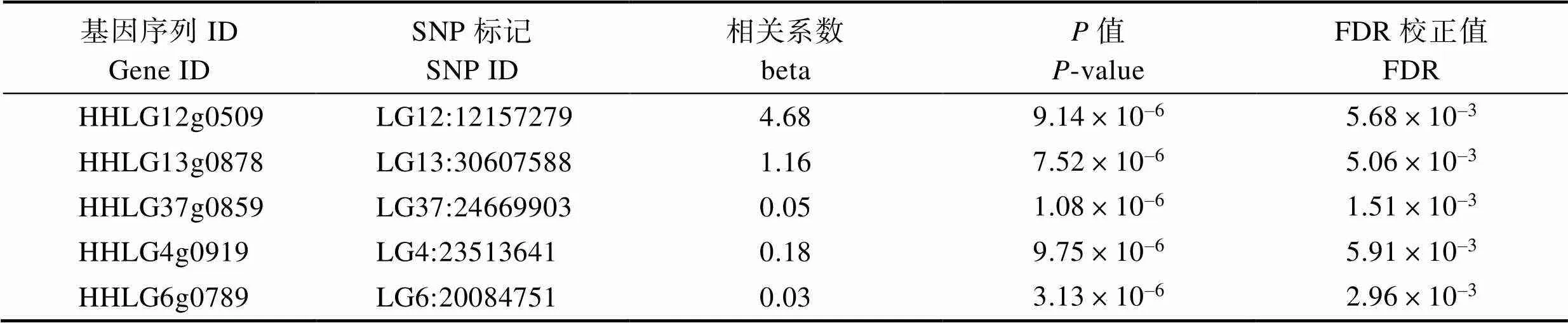
Tab.2 SNP loci in cis-regulation of trim25 genes in liver of C. carpio
3 討論
在進(jìn)化過程中,脊椎動物出現(xiàn)了基因數(shù)目的顯著增加,同時伴隨著SPRY結(jié)構(gòu)域的增加(Sardiello, 2008)。目前,已在人類基因組中鑒定出75個基因(Versteeg, 2013),在河豚基因組中發(fā)現(xiàn)了66個基因,在斑馬魚基因組中鑒定出208個基因(Boudinot, 2011),說明家族數(shù)量在不同物種中的差異很大,且與人類相比,硬骨魚類的基因家族進(jìn)化速度更快。硬骨魚類基因組中基因存在的多樣性和豐富變異,為基因家族的比較分析提供了理想的模型。鯉TRIM25蛋白屬于TRIM家族第二類成員,通過氨基酸多序列比對分析結(jié)果發(fā)現(xiàn),鯉與鯉科魚類金線鲃、斑馬魚的TRIM25蛋白結(jié)構(gòu)在3個保守結(jié)構(gòu)域高度相似。鯉12號染色體上的基因HHLG12g0509與斑馬魚上下游基因完全一致,基因下游的2個基因分別是和;上游距離最近的基因是,也是哺乳動物人、小鼠、雞、斑馬魚、鯉共有的基因;基因是人、小鼠、雞共有的基因。共線性的結(jié)果體現(xiàn)了硬骨魚類與高等動物之間的進(jìn)化差異,以及鯉在經(jīng)歷第4輪全基因組復(fù)制后,基因多樣性有所增加。相對于鯉科其他二倍體魚類,鯉經(jīng)歷了額外一輪全基因組復(fù)制,形成了四倍化的基因組,在基因組復(fù)制及之后的漫長進(jìn)化過程中,基因組的許多區(qū)域產(chǎn)生大量的結(jié)構(gòu)變異,如倒位、易位、插入、缺失等,類似鯉11號染色體的倒位現(xiàn)象大量存在。
表3 鯉腦組織順式調(diào)控基因表達(dá)的SNP
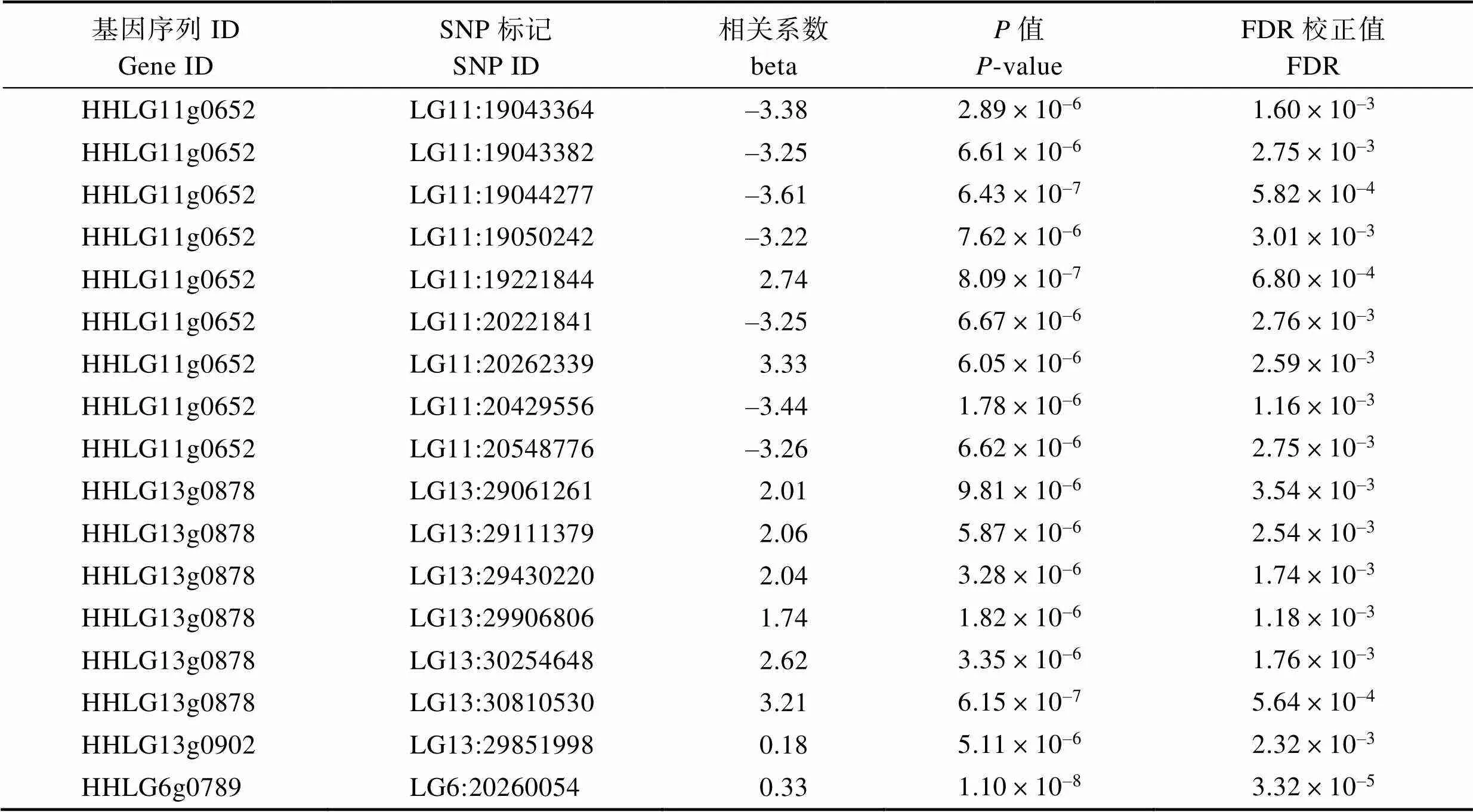
Tab.3 SNP loci in cis-regulation of trim25 genes in brain of Cyprinus carpio
本研究鑒定了鯉TRIM25蛋白有16個拷貝之多,這可能是進(jìn)化過程中,由于環(huán)境壓力導(dǎo)致的基因家族的擴(kuò)張。此外,研究表明,基因家族的擴(kuò)展與先天免疫及適應(yīng)性免疫的進(jìn)化驚人得平行,這表明TRIM蛋白可能在日益復(fù)雜的免疫功能中發(fā)揮核心調(diào)節(jié)因子的作用(Malfavon-Borja, 2013; Versteeg, 2014)。在鯉TRIM25的16個拷貝中,有6個拷貝具有完整的功能結(jié)構(gòu)域,而另外10個拷貝雖然具有TRIM25的部分結(jié)構(gòu)域,但并不完整,這可能由于進(jìn)化過程中發(fā)生了基因的亞功能化或去功能化。
為了精細(xì)定位魚類抗病毒性狀相關(guān)的基因和通路,高通量分子標(biāo)記的采集和大樣本的表型分析是非常必要的。對于鯉這樣經(jīng)歷了第4輪全基因組復(fù)制的異源四倍化物種,許多基因發(fā)生了擴(kuò)增和結(jié)構(gòu)變異,增加了性狀精準(zhǔn)定位的難度。在Ross團(tuán)隊(duì)發(fā)表的鯉抗皰疹病毒QTL研究中,初步定位到基因(Palaiokostas, 2018),但由于其研究采用了舊版本參考基因組,裝配和注釋結(jié)果不夠準(zhǔn)確,因此,并未精確解析該基因的多拷貝的差異。本研究基于高質(zhì)量的新版本基因組開展了鯉基于多個拷貝的篩選和分析,為了解析基因的調(diào)控機(jī)制,還開展了eQTL分析。eQTL是Jansen等(2001)提出的將全基因組中每個基因的表達(dá)量作為數(shù)量性狀,對其進(jìn)行QTL定位分析的方法。eQTL分析填補(bǔ)了基因型和表型之間的空白(Kim, 2012),與全基因組關(guān)聯(lián)研究(GWAS)和差異表達(dá)分析的整合研究已經(jīng)廣泛應(yīng)用于人類疾病研究和動植物育種研究(Joehanes, 2017; Nieuwenhuis, 2016; Pavlides, 2016),eQTL分析新統(tǒng)計方法和軟件的開發(fā)加速了該領(lǐng)域的進(jìn)展(Zhu, 2016)。本研究鑒定了肝臟組織中多個調(diào)控位點(diǎn),表明了肝臟與免疫功能的密切相關(guān)(Huang, 2013; Lee, 2010)。根據(jù)鯉的TRIM25在腦和肝臟組織的表達(dá)分析及eQTL分析結(jié)果,初步篩選了候選的調(diào)控位點(diǎn),對于目前處于起步階段的鯉抗病毒QTL和GWAS研究有重要的提示作用。
本研究初步定位了鯉基因的多個拷貝,比較了鯉與其他物種的序列差異、進(jìn)化關(guān)系和共線性相似度,揭示了鯉各拷貝之間的結(jié)構(gòu)多樣性和在組織中表達(dá)的分化情況,篩選了可能調(diào)控基因表達(dá)的順時調(diào)控位點(diǎn),為今后鯉的基因功能精細(xì)研究奠定了基礎(chǔ),也為魚類抗病研究和育種提供了重要參考。
ASLAM M L, ROBLEDO D, KRASNOV A,. Quantitative trait loci and genes associated with salmonid alphavirus load in Atlantic salmon: Implications for pancreas disease resistance and tolerance. Scientific Reports, 2020, 10(1): 10393
BOUDINOT P, VAN DER AA LM, JOUNEAU L,. Origin and evolution of TRIM proteins: New insights from the complete TRIM repertoire of zebrafish and pufferfish. PLoS One, 2011, 6(7): e22022
GAO J, MA J L, LIU Y,. Bioinformatics analysis of sox gene family in. Progress in Fishery Sciences, 2016, 37(2): 41–48 [高進(jìn), 馬佳璐, 劉洋, 等. 半滑舌鰨()基因家族生物信息學(xué)分析. 漁業(yè)科學(xué)進(jìn)展, 2016, 37(2): 41–48]
GUO J, CAO K, DENG C,. An integrated peach genome structural variation map uncovers genes associated with fruit traits. Genome Biology, 2020, 21(1): 258
HUANG L R, WOHLLEBER D, REISINGER F,. Intrahepatic myeloid-cell aggregates enable local proliferation of CD8(+) T cells and successful immunotherapy against chronic viral liver infection. Nature Immunology, 2013, 14(6): 574–583
JACOBS A, CARRUTHERS M, YURCHENKO A,. Parallelism in eco-morphology and gene expression despite variable evolutionary and genomic backgrounds in a Holarctic fish. PLoS Genetics, 2020, 16(4): e1008658
JANSEN R C, NAP J P. Genetical genomics: The added value from segregation. Trends in Genetics, 2001, 17(7): 388–391
JIN Y, JIA K, ZHANG W,. Zebrafish TRIM25 promotes innate immune response to RGNNV infection by targeting 2CARD and RD regions of RIG-I for K63-linked ubiquitination. Frontiers in Immunology, 2019, 10: 2805
JOEHANES R, ZHANG X, HUAN T,. Integrated genome-wide analysis of expression quantitative trait loci aids interpretation of genomic association studies. Genome Biology, 2017, 18(1): 16
KIM Y A, PRZYTYCKA T M. Bridging the gap between genotype and phenotype via network approaches. Frontiers in Genetics, 2012, 3: 227
KORDAS G, RUDRA P, HENDRICKS A,. Insight into genetic regulation of miRNA in mouse brain. BMC Genomics, 2019, 20(1): 849
LEE W Y, MORIARTY T J, WONG C H,. An intravascular immune response toinvolves Kupffer cells and iNKT cells. Nature Immunology, 2010, 11(4): 295–302
LOWE W L Jr, REDDY T E. Genomic approaches for understanding the genetics of complex disease. Genome Research, 2015, 25(10): 1432–1441
MALFAVON-BORJA R, SAWYER S L, WU L I,. An evolutionary screen highlights canonical and noncanonical candidate antiviral genes within the primate TRIM gene family. Genome Biology and Evolution, 2013, 5(11): 2141–2154
MANOKARAN G, FINOL E, WANG C,. Dengue subgenomic RNA binds TRIM25 to inhibit interferon expression for epidemiological fitness. Science, 2015, 350(6257): 217–221
MICALE L, CHAIGNAT E, FUSCO C,. The tripartite motif: Structure and function. 2016, 770: 11–25
NICOLAE DL, GAMAZON E, ZHANG W,. Trait-associated SNPs are more likely to be eQTLs: Annotation to enhance discovery from GWAS. PLoS Genetics, 2010, 6(4): e1000888
NIEUWENHUIS M A, SIEDLINSKI M, VAN DEN BERGE M,. Combining genomewide association study and lung eQTL analysis provides evidence for novel genes associated with asthma. Allergy, 2016, 71(12): 1712–1720
PALAIOKOSTAS C, ROBLEDO D, VESELY T,. Mapping and sequencing of a significant quantitative trait locus affecting resistance to koi herpesvirus in common carp. G3 (Bethesda), 2018, 8(11): 3507–3513
PAVLIDES J M, ZHU Z, GRATTEN J,. Predicting gene targets from integrative analyses of summary data from GWAS and eQTL studies for 28 human complex traits. Genome Medicine, 2016, 8(1): 84
PENG Q D. Method for constructing and analyzing biological network based on eQTL data. Master′s Thesis of Harbin Institute of Technology, 2020 [彭啟迪. 基于eQTL數(shù)據(jù)的生物網(wǎng)絡(luò)構(gòu)建與分析方法. 哈爾濱工業(yè)大學(xué)碩士研究生學(xué)位論文, 2020]
SARDIELLO M, CAIRO S, FONTANELLA B,. Genomic analysis of the TRIM family reveals two groups of genes with distinct evolutionary properties. BMC Evolutionary Biology, 2008, 8: 225
TOCCHINI C, CIOSK R. TRIM-NHL proteins in development and disease. Seminars in Cell and Developmental Biology, 2015, 47–48: 52–59
VERSTEEG G A, BENKE S, GARCíA-SASTRE A,. InTRIMsic immunity: Positive and negative regulation of immune signaling by tripartite motif proteins. Cytokine and Growth Factor Reviews, 2014, 25(5): 563–576
VERSTEEG G A, RAJSBAUM R, SáNCHEZ-APARICIO M T,. The E3-ligase TRIM family of proteins regulates signaling pathways triggered by innate immune pattern-recognition receptors. Immunity, 2013, 38(2): 384– 398
WALKER R L, RAMASWAMI G, HARTL C,. Genetic control of expression and splicing in developing human brain informs disease mechanisms. Cell, 2020, 181(2): 484
XU P, XU J, LIU G,. The allotetraploid origin and asymmetrical genome evolution of the common carp. Nature communications, 2019, 10(1): 4625
YANG Y, HUANG Y, YU Y,. RING domain is essential for the antiviral activity of TRIM25 from orange spotted grouper. Fish and Shellfish Immunology, 2016, 55: 304–314
ZHOU Z Z, JING F, WEI K,. Molecular cloning and expression analysis of TRIM25 gene in. Acta Hydrobiologica Sinica, 2019, 43(6): 1189–1196 [周真真, 景斐, 魏可, 等. 大黃魚TRIM25基因克隆和表達(dá)分析. 水生生物學(xué)報, 2019, 43(6): 1189–1196]
ZHU Z, ZHANG F, HU H,. Integration of summary data from GWAS and eQTL studies predicts complex trait gene targets. Nature Genetics, 2016, 48(5): 481–487
Gene Evolution and Expression Regulation of Multiple Copies ofin Common Carp,
ZHU Youxiu1,2, JIANG Yanliang2, ZHANG Qin3, FENG Jianxin3, ZHANG Hanyuan2, WU Biyin1,2, XU Jian2①
(1. National Demonstration Center for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China; 2. Key Laboratory of Aquatic Genomics, Ministry of Agriculture and Rural Affairs, Beijing Key Laboratory of Fishery Biotechnology, Chinese Academy of Fishery Sciences, Beijing 100141, China; 3. Henan Academy of Fishery Sciences, Zhengzhou, Henan 450044, China)
The innate immune system is the main defense against viral infections in teleost fish. As an important part of the innate immune system, the TRIM protein family participates in the regulation of the immune network during virus infection. Among such proteins, TRIM25 has been reported to play an important role in the immune response of many types of fish. In this study, 16 copies of thegene in common carp () were compared with those of other species by phylogenetic and syntenic analyses. Functional domain structures for 16 copies of the TRIM25 protein were predicted, and the expression and cis-regulatory network in tissues for each gene copy were compared. Sequence alignment and phylogenetic analysis showed that the TRIM25 protein structure ofwas highly similar to that ofand, but distinct from that of other species beyond Cyprinidae. The results of gene synteny indicated that the upstream and downstream genes ofwere relatively conserved in the evolution of different species. Structural analysis of TRIM25 showed that six of the sixteen copies inhad complete functional domains; five of these copies were highly expressed in liver and brain tissue. In the expression quantitative trait loci (eQTL) regulatory network for, 5 and 17 SNPs were determined to regulateexpression in liver and brain tissue, respectively. In this study, the sequence differences of multiple copies ofin common carp were compared, and the evolutionary relationship and synteny ofwere identified. The diversity of the structure and tissue expression ofgenes in the common carp were revealed, and the SNP sites that may regulategene expression were identified, providing a reference for future research onrelated regulation and disease resistance in the.
; TRIM25; Protein structure; Phylogenetic tree; eQTL
S917.4
A
2095-9869(2022)03-0024-09
10.19663/j.issn2095-9869.20210127001
http://www.yykxjz.cn/
朱優(yōu)秀, 江炎亮, 張芹, 馮建新, 張瀚元, 吳碧銀, 許建. 鯉多拷貝基因進(jìn)化和表達(dá)調(diào)控初探. 漁業(yè)科學(xué)進(jìn)展, 2022, 43(3): 24–32
ZHU Y X, JIANG Y L, ZHANG Q, FENG J X, ZHANG H Y, WU B Y, XU J. Gene evolution and expression regulation of multiple copies ofin common carp,. Progress in Fishery Sciences, 2022, 43(3): 24–32
XU Jian, E-mail: xuj@cafs.ac.cn
* 中國水產(chǎn)科學(xué)研究院基本科研業(yè)務(wù)費(fèi)(2020B003; 2020TD24)和國家重點(diǎn)研發(fā)計劃(2018YFD0900102)共同資助 [This work was supported by the Central Public-Interest Scientific Institution Basal Research Fund, CAFS (2020B003; 2020TD24), and National Key Research and Development Program (2018YFD0900102)]. 朱優(yōu)秀,E-mail: zhuyx0508@163.com
許 建,副研究員,E-mail: xuj@cafs.ac.cn
2021-01-27,
2021-03-01
(編輯 馮小花)
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學(xué)生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學(xué)苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(shù)(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
