薛家島海水網箱養殖海域貝藻綜合養殖效果及其影響因素分析*
2022-06-15 02:35:40吳文廣張繼紅趙云霞張坤陽王亮亮
漁業科學進展 2022年3期
關鍵詞:實驗
吳文廣 張繼紅 劉 毅 趙云霞 張坤陽 王亮亮
薛家島海水網箱養殖海域貝藻綜合養殖效果及其影響因素分析*
吳文廣1,3張繼紅1,2①劉 毅1趙云霞1張坤陽1王亮亮1
(1. 中國水產科學研究院黃海水產研究所 農業農村部海洋漁業可持續發展重點實驗室 山東 青島 266071; 2. 青島海洋科學與技術試點國家實驗室海洋漁業科學與食物產出過程功能實驗室 山東 青島 266071; 3. 上海海洋大學水產與生命學院 上海 201306)
本研究于2018年9—11月在山東省青島市薛家島海域構建了包含魚類網箱、馬氏珠母貝()和異枝麒麟菜()的多營養層次綜合養殖(IMTA)模式,開展現場實驗的同時,在室內測定了馬氏珠母貝的攝食生理指標和異枝麒麟菜的營養鹽吸收速率,對比分析了貝藻綜合養殖對網箱養殖區的水質改善效果。結果顯示,實驗點馬氏珠母貝殼高及特定生長率(SGR)均顯著高于對照點,水層為0.50~0.62 m的馬氏珠母貝平均殼高顯著高于其他水層。實驗點異枝麒麟菜的凈增重顯著高于對照點,水深為0.60~0.90 m的生長速率最高;在24.45℃條件下,馬氏珠母貝對顆粒有機物(POM)的攝食率、吸收率和吸收效率均值分別為(4.13±0.77) mg/(h·ind.)、(1.04±0.24) mg/(h·ind.)和(25.00±2.51)%,均顯著高于20.74℃實驗組;在實驗溫度范圍內,異枝麒麟菜對N、P、Si營養鹽的吸收速率均隨著溫度升高呈先升高再降低的趨勢,最高值出現在30℃。實驗期間,馬氏珠母貝攝食和吸收的POM分別為778.08和144.47 g,異枝麒麟菜吸收的N、P、Si總量分別為8.55、1.11和9.18 g,均高于對照點。研究表明,溫度、養殖深度、POM及營養鹽濃度是影響馬氏珠母貝和異枝麒麟菜生長及生理代謝的主要因素。
海水網箱養殖;馬氏珠母貝;異枝麒麟菜;多營養層次綜合養殖
近年來,隨著淺海網箱養殖產生的環境污染及生態問題日益凸顯,多營養層次綜合養殖(IMTA)作為一種健康可持續發展的海水養殖理念,可實現營養物質在不同營養級生物間的傳遞和再循環,同時,可提高食物產出效率,達到控制養殖水域富營養化的生態高效養殖目的(Chopin, 2001)。唐啟升等(2013)研究表明,多營養層次綜合水產養殖是應對多重壓力脅迫下近海生態系統顯著變化的一條有效途徑。目前,該養殖模式已經在世界多個國家(中國、加拿大、挪威、以色列等)廣泛實踐,并取得了諸多的積極效果(Neori, 2004; Gui, 2018; Ashkenazi, 2019; 張繼紅等, 2013a)。以貝藻為主體的IMTA是水產養殖業綠色發展的代表性模式,通過引入濾食性貝類和大型藻類,利用貝類的濾食及大型藻類光合作用吸收營養鹽的功能,可以減輕網箱養殖產生的環境壓力,降低有機物和營養鹽負荷(張繼紅等, 2013b)。
20世紀70年代,我國已開展了海水貝藻混養的研究,并取得了較好的養殖效果,但多集中于扇貝、貽貝()、牡蠣)與海帶()等北方養殖種類(王德利, 2001; 高樺楠等, 2016)。馬氏珠母貝()又稱合浦珠母貝,廣泛分布在熱帶或亞熱帶海域,以過濾海水中微藻、浮游性動物的幼蟲和海水中的有機碎屑雜質等為食(鄧正華等, 2016)。麒麟菜()是一種世界性海藻,主要分布在熱帶海域以及亞熱帶海域,而瓊枝麒麟菜()和異枝麒麟菜(),又名長心卡帕藻(),是我國南方地區養殖的主要經濟種類(陳攀等, 2018)。針對我國北方傳統貝藻養殖所面臨的問題,即夏季水溫過高導致櫛孔扇貝()、蝦夷扇貝()等死亡率上升、產量下降及藻類養殖種類空缺等,馬氏珠母貝和麒麟菜是開展貝藻綜合養殖的理想經濟種類。通過嘗試引入馬氏珠母貝、異枝麒麟菜等相對更耐高溫的熱帶種類到北方海域開展魚貝藻IMTA模式,有效地利用夏季北方海域相對較高的水溫特點及網箱養殖區較高的營養鹽濃度(王德利, 2001),探討其在北方海域進行規模化養殖的可能性,可作為北方海域夏季海水養殖種類的有益補充。目前,關于淺海網箱養殖、馬氏珠母貝與異枝麒麟菜的混養效果研究報道較少。本研究以山東青島薛家島網箱養殖海域為研究區域,構建了包含魚類網箱、馬氏珠母貝和異枝麒麟菜的IMTA模式,對比并分析了貝藻綜合養殖對網箱養殖區的水質改善效果,以期為淺海魚貝藻IMTA模式的建立提供數據支撐。
1 材料與方法
1.1 實驗站位設置
實驗所需馬氏珠母貝和異枝麒麟菜均采自海南省三亞市養殖區,空運至青島實驗室暫養7 d后開始實驗。2018年9—11月在山東青島薛家島海域設置網箱養殖區(實驗點, 35°55′16.6″N, 120°13′04.1″E)和非養殖區(對照點, 35°55′27.6″N, 120°13′15.3″E),開展海上貝藻綜合養殖對比實驗(圖1)。網箱養殖區鱸魚()養殖密度為10.8尾/m3。
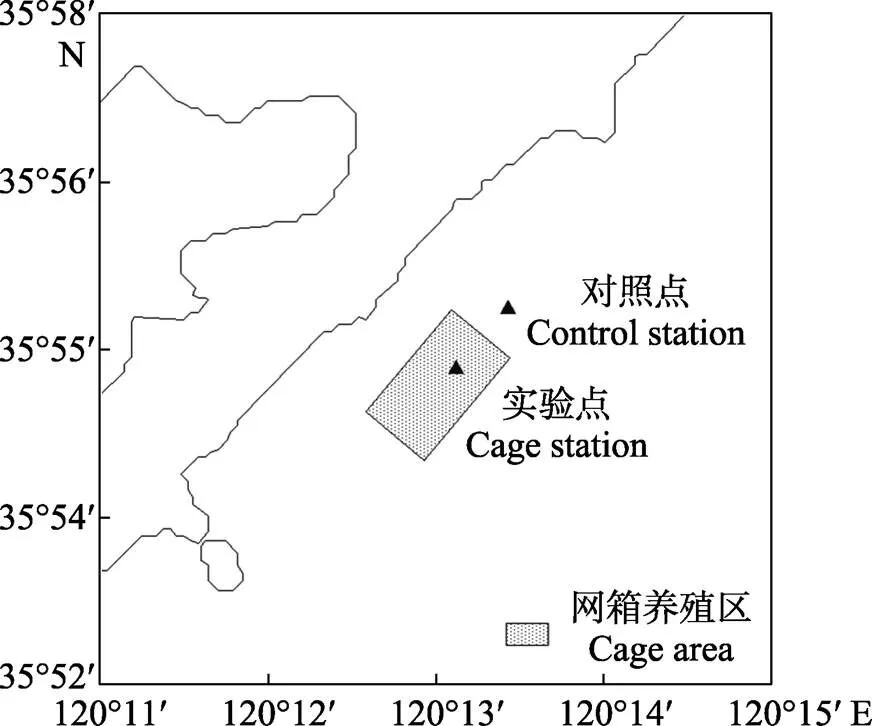
圖1 現場綜合養殖實驗站位
1.2 布置方案與樣品采集
1.2.1 布置方案 參照麒麟菜栽培技術規程(海南省地方標準)(DB46/T192-2010),挑選色澤鮮亮的異枝麒麟菜進行綁苗,主繩長為2.5 m,苗距為30 cm,每條主繩懸掛7個不同深度的異枝麒麟菜并編號,測量每層初始苗的濕重,分別在對照點和實驗點懸掛異枝麒麟菜各4繩并編號。參照馬氏珠母貝養成技術規程(海南省地方標準)(DB46/T2862-2014),挑選健康有活力的馬氏珠母貝置于養殖吊籠內,每層放置3個,共7層,每籠21個馬氏珠母貝,對照點和實驗點各懸掛5籠,測量馬氏珠母貝的初始濕重、殼高。在網箱養殖區,將異枝麒麟菜第1層苗繩懸掛在海水表層,馬氏珠母貝養殖籠懸掛在水下0.26 m處,苗繩和養殖籠下端均懸掛墜石使它們在海水中垂直展開,即馬氏珠母貝的吊養深度分別為0.26、0.32、0.38、0.44、0.50、0.56和0.62 m,異枝麒麟菜的吊養深度分別為 0、0.3、0.9、1.2、1.5、1.8、2.1和2.4 m。非養殖區(對照點)也同樣操作。
1.2.2 樣品采集和測定 養殖實驗于2018年9月21日—11月2日開展,并于10月15日和11月2日分別在實驗點和對照點采集樣品。溫度、鹽度、pH等環境參數均采用多參數水質儀(YSI-EXO2型)測定。總懸浮顆粒物(TPM)、顆粒有機物(POM)、營養鹽等環境參數的采集及測定均采用《海洋監測規范》(GBl7378.4–2007)的方法;葉綠素采用Turner- Designs Trilogy熒光儀(美國)進行測定(吳文廣等, 2018)。使用電子天平(梅特勒-ME-2002, 美國)現場稱量每個實驗組、每繩、每層異枝麒麟菜的濕重并記錄對應編號,統計馬氏珠母貝養殖籠內每層的死亡個數,并記錄對應編號,使用電子天平稱量每層馬氏珠母貝的生物學參數(殼高、濕重),稱量后均放回原處。實驗結束后,全部取回至實驗室進行最終的測量分析。
1.3 室內生理實驗
1.3.1 馬氏珠母貝攝食生理實驗 參照張繼紅 等(2013a, b)的方法,采用室內靜水法測定馬氏珠母貝的濾水率(Clearance rate, CR)、攝食率(Feeding rate, FR)、吸收率(Absorption rate)及吸收效率(Absorption efficiency)等生理指標,每個實驗組設置5個平行和3個空白對照,每個平行組放置1個馬氏珠母貝,實驗持續2 h后,收集糞便和假糞。實驗結束后,將貝類分別置于過濾海水中,排空2 h,再次收集糞便,測定實驗用貝的殼高、濕重等生物學指標,實驗用水為網箱區表層自然海水。
1.3.2 異枝麒麟菜營養鹽吸收速率實驗 選取生長正常的異枝麒麟菜,充氣暫養7 d后開始實驗。使用吸水紙吸干藻體表面的水分后,分別測量并記錄異枝麒麟菜的濕重,平均濕重為(9.82±0.51) g,實驗海水使用網箱區表層自然海水脫脂棉過濾,設置3個實驗組和1個空白對照組,實驗容器為體積12 L聚乙烯材質整理箱,每個實驗容器放入實驗海水10 L,微充氣培養,溫度梯度設置為17℃、20℃、25℃、30℃、35℃共5個梯度,每個溫度梯度結束后,按照每天升溫2℃,升至實驗溫度暫養3 d后,繼續進行下一個溫度梯度實驗。實驗期間,培養箱設置12 h光照和12 h黑暗,每個溫度梯度實驗持續24 h,按1.2.2所述方法測定營養鹽濃度。
1.4 計算方法
馬氏珠母貝和異枝麒麟菜的特定生長率(specific growth rate, SGR, %/d)計算公式:
SGR=100×(ln2–ln1)/
式中,2為結束時濕重(g)或殼高(mm),1為初始濕重(g)或殼高(mm),實驗持續時間(d)。
馬氏珠母貝攝食生理指標的相關計算公式參考周毅等(2002)和張繼紅等(2013a、b)的方法。
異枝麒麟菜對各項營養鹽吸收速率參照方哲 等(2008)的方法,計算公式:
=(0–N)×/(×)
式中,為營養鹽吸收速率(μmol/g·h),0為實驗結束時空白對照組的各項營養鹽濃度(μmol/L),N為實驗過程中各個取樣時間點的各項營養鹽濃度(μmol/L),為實驗所用海水的體積(L),為實驗時間(h),為異枝麒麟菜的濕重(g)。
馬氏珠母貝對POM的攝食及吸收總量計算公式:
pom=×W××T×10–3
式中,pom為馬氏珠母貝對POM的攝食總量或吸收總量(g),為馬氏珠母貝最終剩余總個數(ind.),W為馬氏珠母貝養殖最終單位個體軟組織干重(g/ind.),為馬氏珠母貝對POM的攝食率或吸收率[mg/(h·g DW)],T為養殖時間(h)。
異枝麒麟菜N、P、Si去除總量計算公式:
=G×××T×10–6
式中,為異枝麒麟菜對N、P或Si的去除總量(g),G為異枝麒麟菜凈增重(濕重計)(g),為N、P或Si元素的摩爾質量(g/moL),為20℃~25℃時,異枝麒麟菜對N、P或Si營養鹽吸收速率的平均值[μmol/(g·h)],T為養殖時間(h)
1.5 數據處理
實驗數據采用Microsoft Excel 2010、Surfer8.0和Matlab統計分析軟件進行分析及繪圖,采用SPSS17.0軟件進行獨立樣本檢驗分析組間差異(<0.05為差異顯著,<0.01為差異極顯著),貝藻的SGR與主要環境因子間的關系采用冗余分析(redundancy analysis, RDA),分析過程使用Canoco for Windows 4.5軟件按照國際通用標準完成(吳文廣等, 2018)。
2 結果
2.1 環境數據分析
近海網箱養殖實驗海區環境參數詳見表1。單因子方差分析(one-way ANOVA)表明,實驗期間,實驗點TPM、POM、氨氮(NH4+-N)及溶解無機氮(DIN)濃度均顯著高于對照點(<0.05),而透明度顯著低于對照點;除第1次取樣外,實驗點硝酸鹽氮(NO3–-N)濃度、葉綠素濃度均顯著高于對照點(<0.05);除初始取樣外,實驗點PO43–-P濃度均顯著高于對照點(<0.05);SiO32–-Si濃度在初始取樣時顯著低于對照點,而第1次和第2次取樣時則相反(<0.05)。
表1 近海網箱養殖實驗海區環境參數
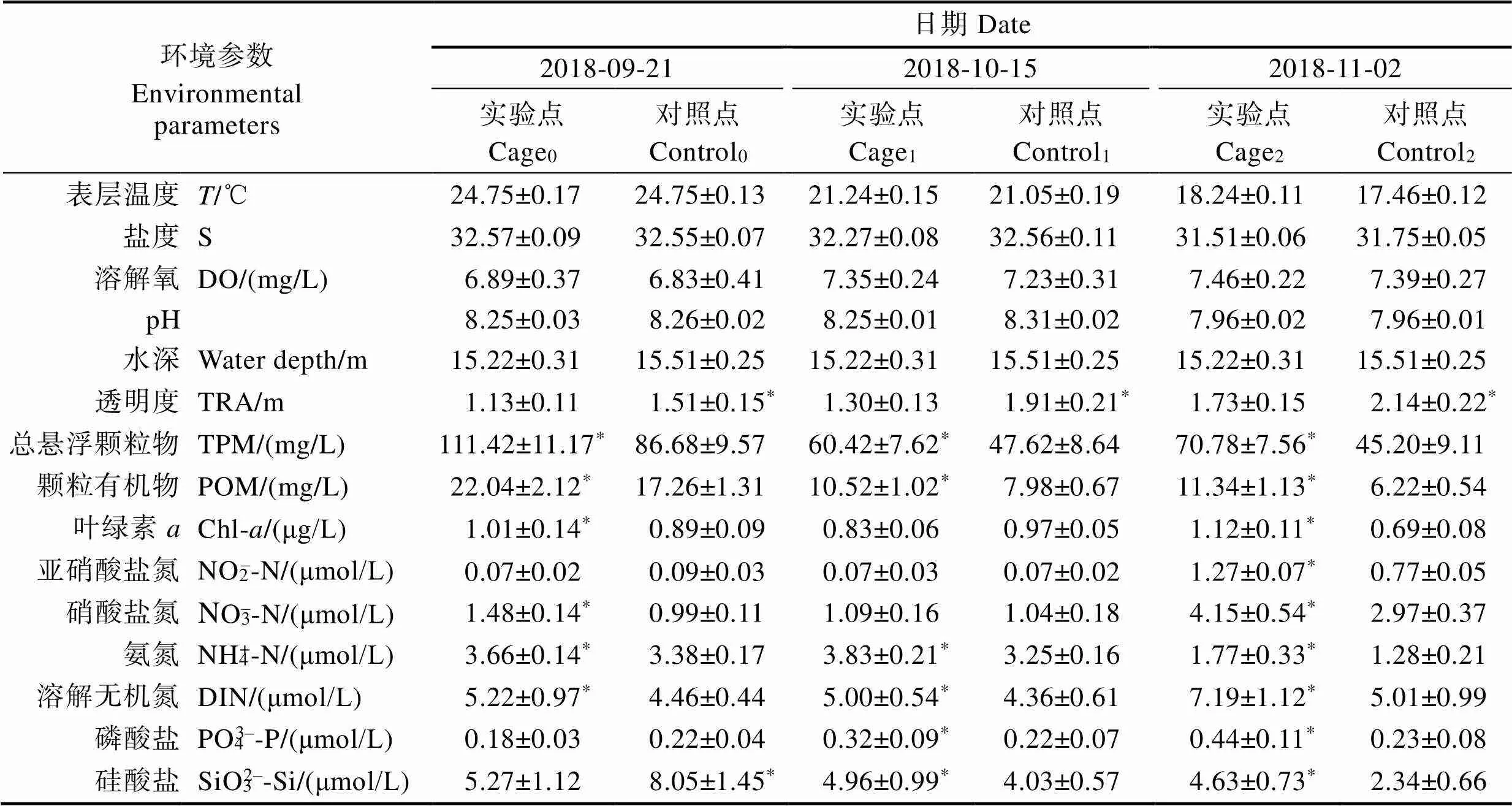
Tab.1 The environmental parameters of experimental area in offshore fishcage farming
注:同一取樣日期不同數據列中上標有“*”表示差異顯著(<0.05),Cage0、Cage1、Cage2分別代表實驗點初始、第1次、第2次測量;Control0、Control1、Control2分別代表對照點初始、第1次、第2次測量;下同
Note: “*”represented significant differences in the same sampling date, the same below, Cage0, Cage1, Cage2refers to the initial, first and second shell height in fish cage area, respectively; Conctrol0、Conctrol1、Conctrol2refers to the initial, first and second shell height in control area, the same as below
2.2 貝藻綜合養殖效果分析
2.2.1 馬氏珠母貝生長狀況 不同時間、不同海水深度的馬氏珠母貝殼高和濕重的生長狀況見圖2和圖3。結果表明,第3次取樣時,實驗點和對照點馬氏珠母貝殼高和濕重均顯著高于初始值(<0.05),實驗點馬氏珠母貝殼高在0.38 m和0.50~0.62 m水深顯著高于其他水層(<0.05),對照點馬氏珠母貝殼高在0.32和0.50 m處均顯著低于其他水層(<0.05),實驗點馬氏珠母貝的平均殼高和平均濕重分別為(43.71±3.64) mm和(8.42± 1.04) g,對照點馬氏珠母貝的平均殼高和平均濕重分別為(38.38±4.50) mm和(7.83±1.80) g。方差分析表明,第3次取樣時,實驗點馬氏珠母貝殼高顯著高于對照點(<0.05)。
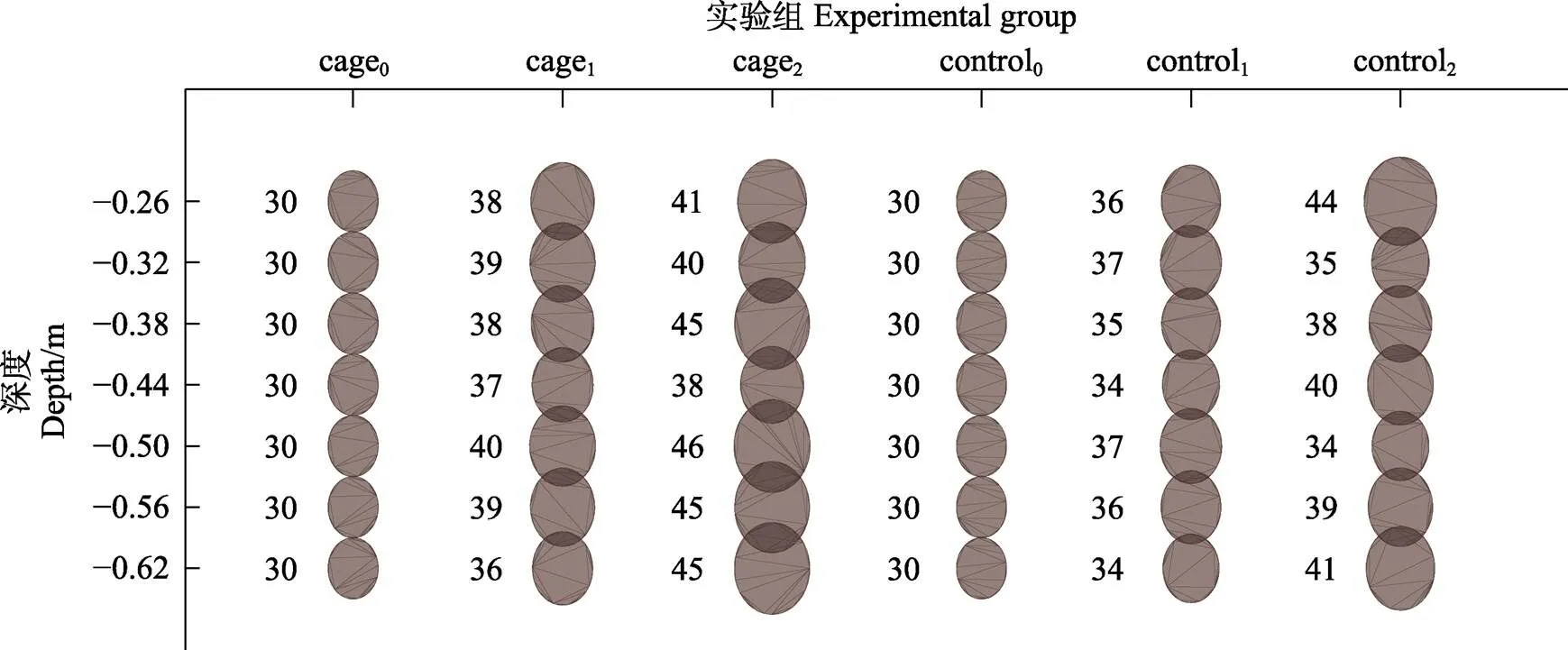
圖2 實驗點和對照點馬氏珠母貝殼高生長狀況(圓形的大小表示貝類的殼高)/mm
cage0、cage1、cage2分別代表實驗點初始、第1次、第2次測量;conctrol0、conctrol1、conctrol2分別代表對照點初始、第1次、第2次測量;下同
cage0, cage1, cage2refers to the initial, first and second shell height in fish cage area, respectively; conctrol0、conctrol1、conctrol2refers to the initial, first and second shell height in control area, the same as below

圖3 實驗點和對照點馬氏珠母貝濕重生長狀況(圓形的大小表示貝類的濕重)/g
馬氏珠母貝在不同深度的實驗點和對照點的SGR結果見表2。從表2可以看出,實驗點馬氏珠母貝殼高SGR在0.50 m時為最大值(0.96),在0.44 m為最小值(0.53);而濕重SGR最大值出現在0.26 m,為1.66,最小值出現在深度為0.44 m (0.83);對照點殼高SGR在0.26 m時為最大值(0.86),在0.50 m處為最小值(0.29);濕重SGR最大值出現在深度為0.62 m (1.58),最小值出現在深度為0.56 m (0.96)。實驗點的馬氏珠母貝殼高SGR平均值顯著高于對照點(<0.05)。
2.2.2 異枝麒麟菜生長狀況 異枝麒麟菜在實驗點和對照點不同養殖深度的生長狀況見圖4。從圖4可以看出,異枝麒麟菜在水深為0.60~0.90 m的生長速率最快,隨著水深的增加,異枝麒麟菜藻體濕重損失較大。經過43 d的海上養殖實驗,實驗點的異枝麒麟菜平均每繩總凈增重為(253.48±38.68) g,0.6~ 0.9 m水層平均每繩凈增重為(200.04±33.02) g,而對照點平均每繩總凈增重為(211.09±43.42) g,0.60~0.90 m水層平均每繩凈增重為(106.87±25.96) g,且該水層實驗點異枝麒麟菜的凈增重顯著高于對照點(<0.05)。異枝麒麟菜實驗點和對照點濕重的SGR結果見表3。從表3可以看出,水深為0.60~0.90 m時,異枝麒麟菜的SGR顯著高于其他水層,且實驗點的異枝麒麟菜SGR在水深為0.60和1.50 m時顯著高于對照點(<0.05)。
表2 不同深度實驗點和對照點馬氏珠母貝的SGR (%/d)
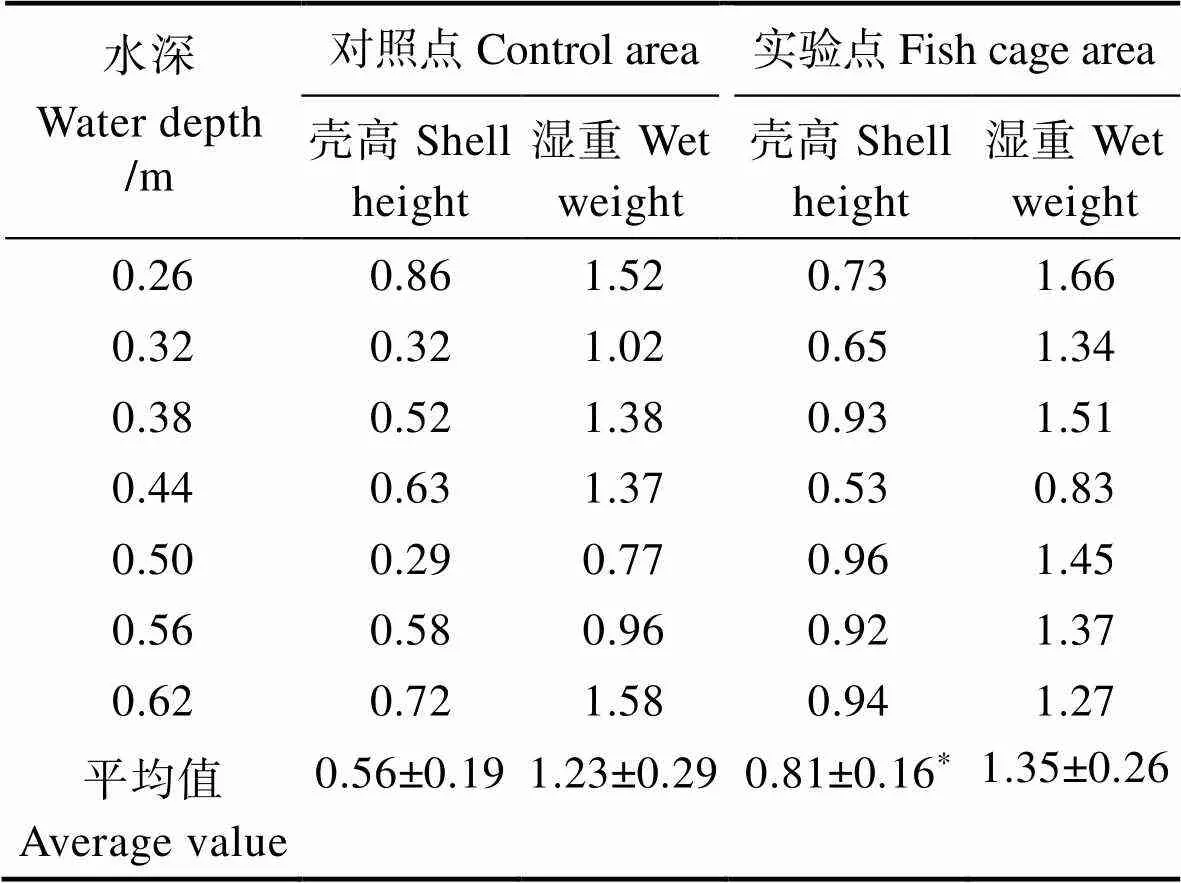
Tab.2 The SGR of shell height and wet weight of P. martensi in fishcage area and control area in different depth
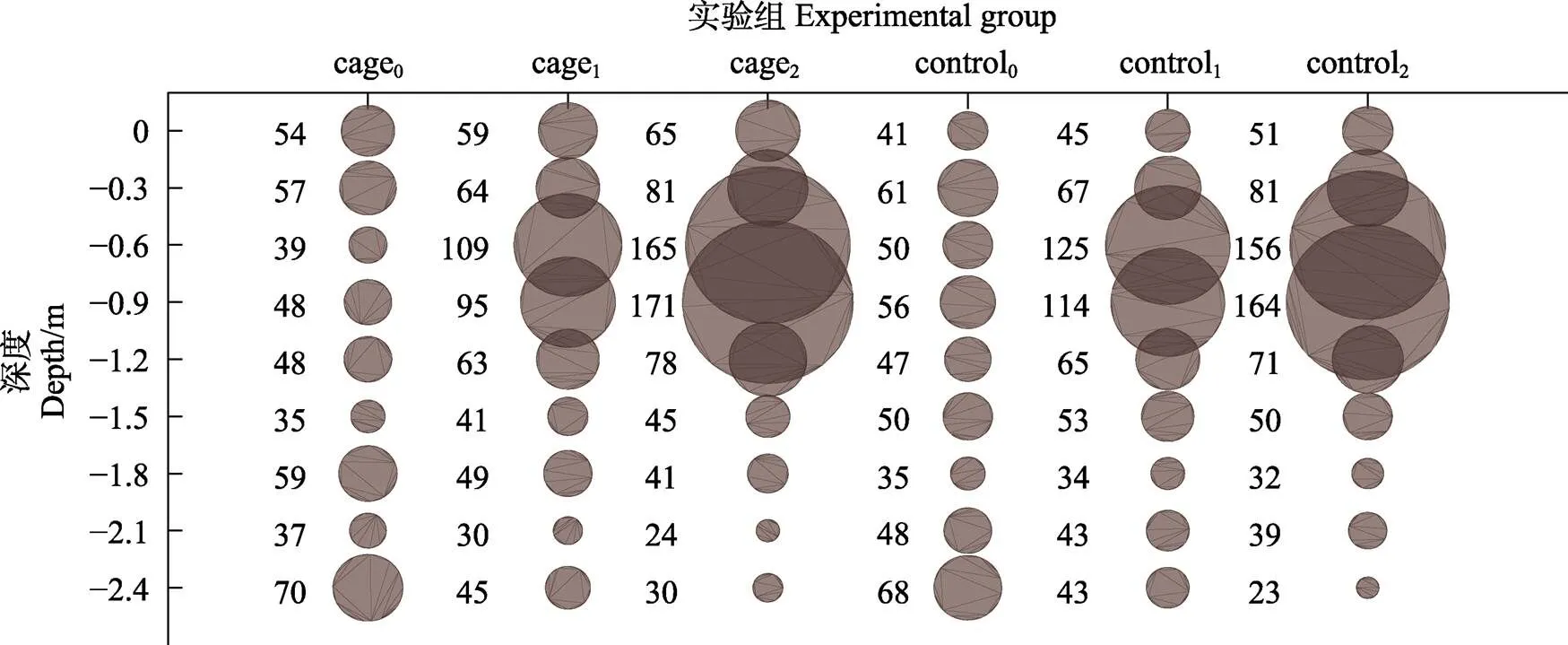
圖4 實驗點和對照點異枝麒麟菜生長狀況(圓形的大小指示藻類的濕重)/g
表3 不同深度下實驗點和對照點異枝麒麟菜濕重的SGR (%/d)
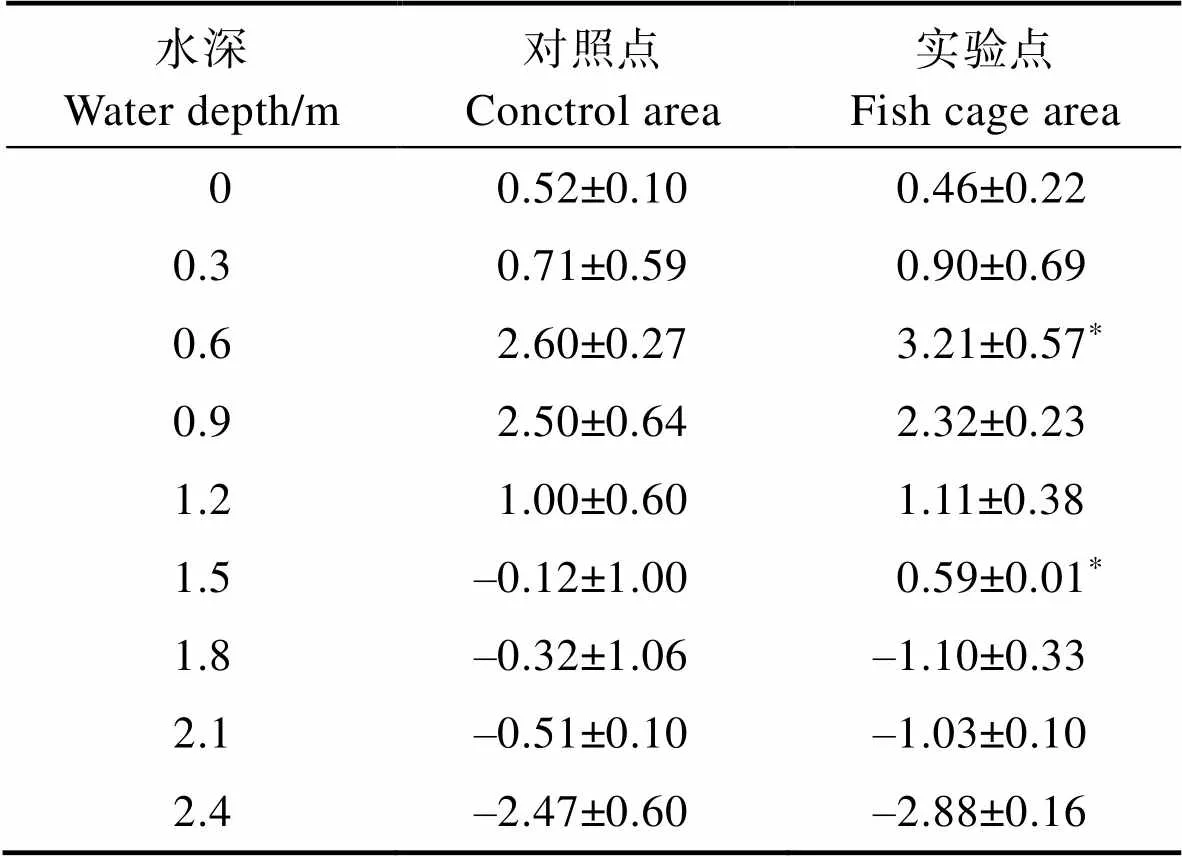
Tab.3 The SGR of E. striatum in fish cage area and control area in different depth
2.3 馬氏珠母貝的攝食生理
馬氏珠母貝攝食生理實驗組水體的環境參數及懸浮顆粒物的特性見表4。從表4可以看出,10月15日實驗組水體的TPM及顆粒無機物(PIM)顯著高于9月21日實驗組,而9月21日實驗組水體溫度及有機物比率()顯著高于10月15日實驗組(<0.05)。馬氏珠母貝的生物學參數見表5。從表5可以看出,2次實驗所用馬氏珠母貝生物學指標相比均無顯著差異(>0.05)。不同溫度條件下,馬氏珠母貝對實驗海區自然海水的攝食生理指標見表6。從表6可以看出,馬氏珠母貝在24.45℃條件下,實驗海區海水POM的攝食率、POM的吸收率和吸收效率平均值分別為(4.13±0.77) mg/(h·ind.)、(1.04±0.24) mg/(h·ind.)和(25.00±2.51)%,均顯著高于20.74℃ (<0.05),表明溫度對馬氏珠母貝的攝食生理產生顯著影響。估算了不同實驗組馬氏珠母貝對POM的去除總量,實驗點和對照點馬氏珠母貝對海水中POM攝食總量見表7。從表7可以看出,經過43 d的海上養殖實驗,實驗點馬氏珠母貝的POM攝食總量和對POM的吸收總量分別為778.08和144.47 g,均高于對照點。
2.4 異枝麒麟菜的營養鹽吸收速率
不同溫度的異枝麒麟菜(濕重計)對N、P、Si營養鹽吸收率見表8。從表8可以看出,異枝麒麟菜的N、P、Si營養鹽吸收速率均隨著溫度升高呈先升高再降低趨勢,最高值出現在30℃。在35℃溫度條件下,對N、P、Si的吸收速率均出現負值,溫度對異枝麒麟菜的營養鹽吸收產生顯著影響。不同實驗組異枝麒麟菜的N、P、Si去除效果總量見表9。從表9可以看出,經過43 d的海上養殖實驗,實驗點異枝麒麟菜的N、P、Si去除總量分別為8.55、1.11和9.18 g,均高于對照點。
表4 實驗組水體的環境參數及懸浮顆粒物的特性
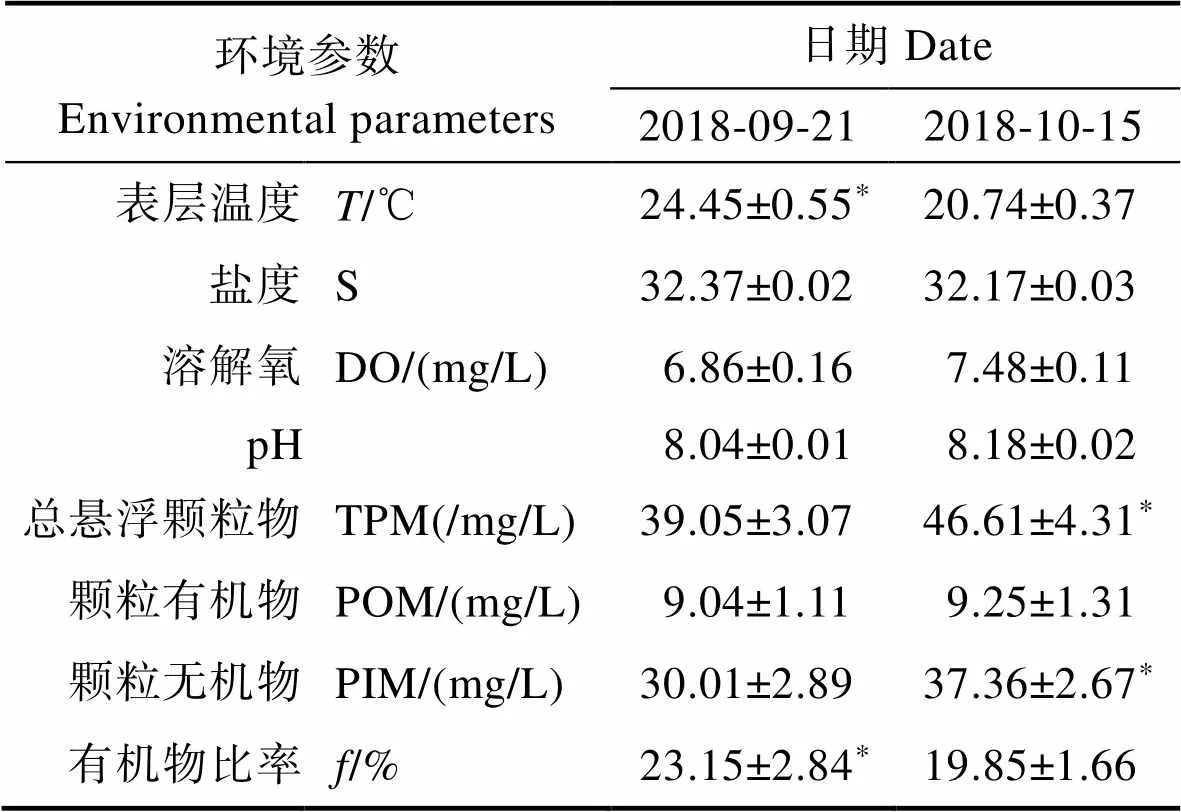
Tab.4 The environmental parameters and characteristics of suspended particles in experimental groups
注:*表示不同實驗日期同一行數據差異顯著(<0.05)
Note: “*”represented significant differences in the different experiment sampling date (<0.05)
表5 實驗組馬氏珠母貝的生物學參數
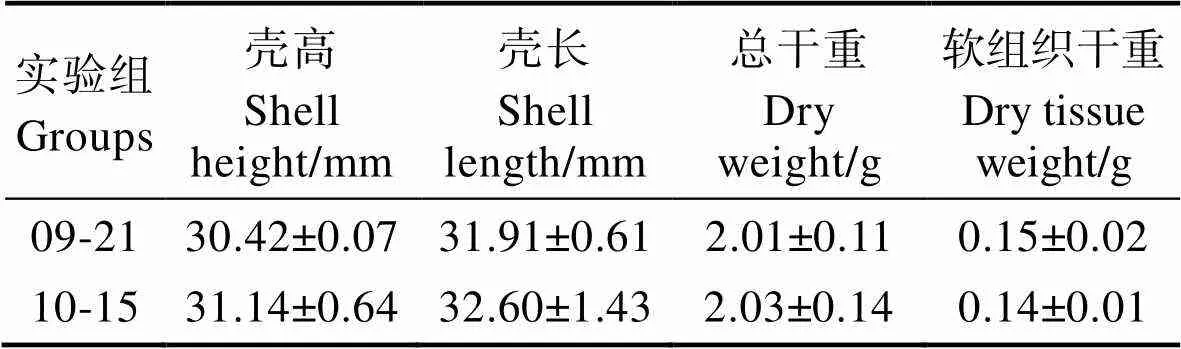
Tab.5 Shell length/height and weight of P. martensi used in experiment
表6 不同溫度下馬氏珠母貝對自然海水的攝食生理指標
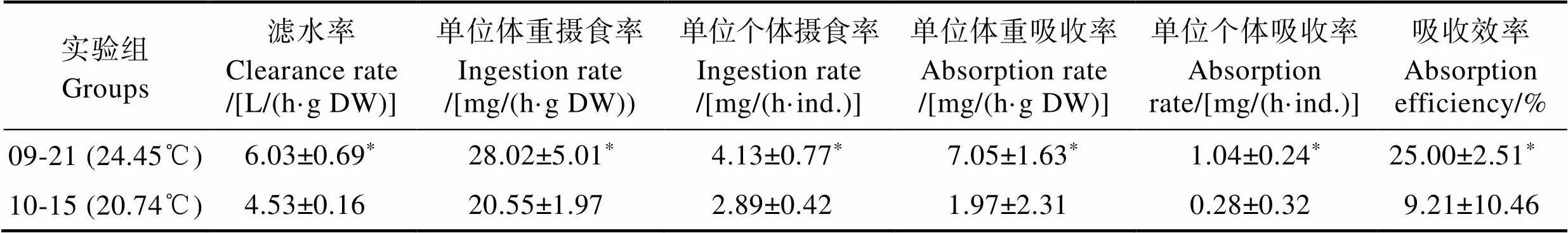
Tab.6 Feeding physiological index of P. martensi in different temperatures
表7 不同實驗組的馬氏珠母貝的濾除效果

Tab.7 Filtering effect of P. martensi from different experimental groups
表8 不同溫度下異枝麒麟菜對氮、磷、硅營養鹽的吸收率
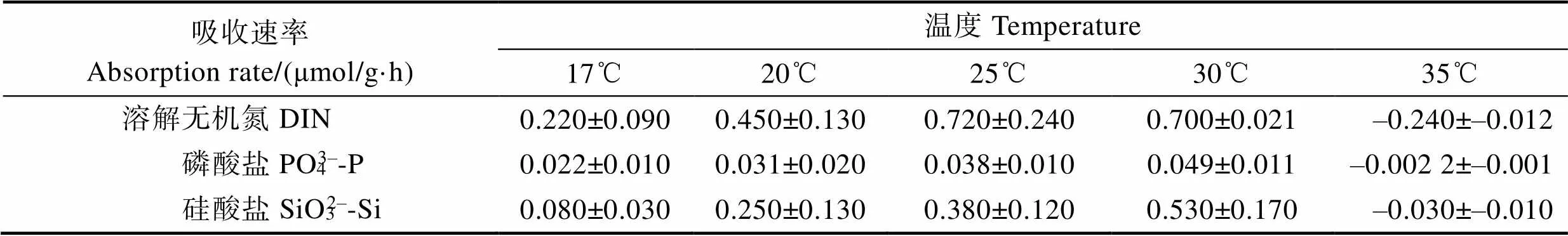
Tab.8 The nutrient absorption rate of E. striatum in different temperature
表9 不同實驗組異枝麒麟菜的N、P、Si去除效果對比
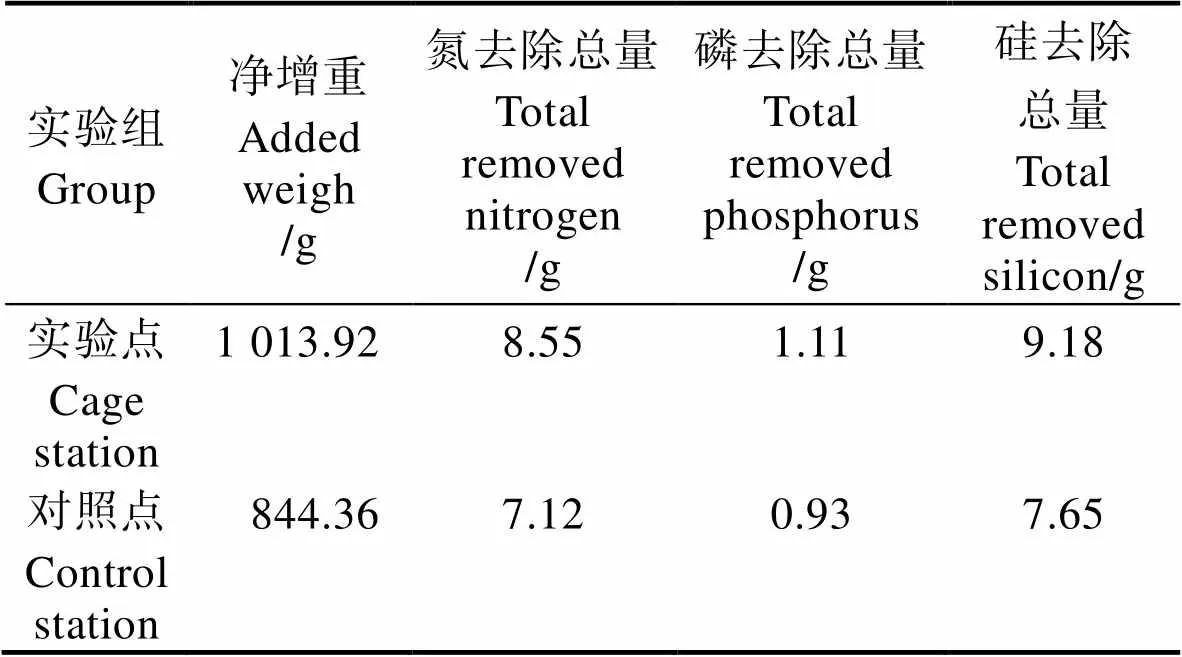
Tab.9 The removal amount of nutrient form E. striatum in different experimental groups
2.5 貝藻生長影響因素分析
馬氏珠母貝與異枝麒麟菜的SGR與環境因子的RDA排序見圖5。從圖5可以看出,環境因子用實線箭頭表示,貝藻SGR用虛線箭頭表示。環境因子與SGR之間夾角的余弦值代表其相關性,即夾角越小,其相關性越強。RDA分析顯示,溫度、TPM、POM對馬氏珠母貝的殼高和濕重的SGR有顯著影響,呈正相關(<0.05);而溫度、SiO32–-Si、pH、NH4+-N和PO43–-P與異枝麒麟菜的濕重SGR呈顯著正相關(<0.01)。
3 討論
3.1 魚貝藻混養系統中的貝藻養殖效果及影響因素
近年來,海水網箱養殖產生的環境及生態系統問題引起人們的關注,開展魚、貝、藻的IMTA是緩解這一環境壓力的有效途徑,目前,已成為國際上主推的養殖理念(張繼紅等, 2018)。然而,養殖種類的搭配、養殖密度、養殖區的合理布局等是開展IMTA的理論基礎(楊紅生等, 2000)。本研究表明,實驗點馬氏珠母貝殼高及SGR均顯著高于對照點,且實驗點馬氏珠母貝殼高在水深為0.5~0.62 m時顯著高于其他水層;異枝麒麟菜在水深為0.60~0.90 m時生長速率最快,且異枝麒麟菜的濕重凈增重顯著高于對照點。與貝、藻單養和混養模式等研究相比(Qian, 1996; Wu, 2003),本研究通過引入魚類網箱養殖單元的魚貝藻的混養效果顯著。魚類網箱養殖是投餌型養殖,是本研究養殖模式運轉的中心,其養殖過程中產生的魚類殘餌、糞便等微小顆粒物質可有效地被馬氏珠母貝攝食并吸收,同時,鱸魚和馬氏珠母貝代謝釋放的N、P、Si等無機營養鹽被異枝麒麟菜吸收并儲存在體內,并釋放O2到該養殖系統中,通過這種養殖方式去除多余的POM和營養鹽,進一步實現了營養物質高效循環利用(Chopin, 2001; Hayashi,, 2008; 張繼紅等, 2013a,b),同時,魚貝藻養殖模式的養殖效果顯著優于貝藻混養模式。

圖5 貝藻SGR與環境因子RDA排序
EU:異枝麒麟菜濕重SGR;SH:馬氏珠母貝殼高SGR;SW:馬氏珠母貝濕重SGR
EU: Wet weight SGR of; SH: Shell height SGR of; SW: Wet weight SGR of,
與貝、藻單養相比,影響魚貝藻養殖效果的因素則更加復雜。本研究表明,溫度、養殖水層、POM及營養鹽濃度是影響馬氏珠母貝和異枝麒麟菜生長及生理代謝的主要因素。溫度是影響海水養殖生物生長的主要因素之一(賴卓欣等, 2019)。近年來,我國北方海域夏季出現水溫過高導致扇貝等養殖種類死亡率、藻類養殖種類空缺等問題(高振錕等, 2017)。本研究引入馬氏珠母貝、異枝麒麟菜等相對更耐高溫的種類到北方海域進行接力養殖,通過有效利用夏季北方海域相對較高的水溫特點及網箱養殖區較高的營養鹽濃度。結果表明,馬氏珠母貝、異枝麒麟菜和鱸魚的混養模式可以適應北方夏季溫度變化,養殖效果顯著。養殖水層也是影響貝藻生長的重要因素,馬氏珠母貝幼貝單養的吊養深度通常在1.0~1.5或2.0~3.0 m,麒麟菜的吊養深度在20~40 cm范圍,其吊養深度主要取決于養殖海域的環境特性(符韶等, 2001; 王愛民等, 2010)。本研究中,實驗點馬氏珠母貝幼貝、異枝麒麟菜吊養深度分別在0.5~0.62 m和0.60~0.90 m的SGR狀況要優于其他水層,可能是該養殖系統中的POM分布特性、營養鹽濃度、透明度及水動力條件等因素對貝藻的適宜生長水層產生了影響(Kasim, 2017)。本研究RDA結果也表明,溫度、養殖深度、POM及營養鹽濃度是影響馬氏珠母貝和異枝麒麟菜SGR主要因素,因此,在構建魚貝藻綜合養殖模式時,需全面了解不同水層的POM及營養鹽濃度等因素的分布特性,進行合理的規劃,從而使IMTA養殖系統具有更高的食物產出能力和實現營養物質的高效利用。
3.2 魚、貝、藻混養系統中的貝藻養殖環境效應
海水魚類網箱養殖是一種高密度、集約化的養殖方式,給養殖者帶來巨大經濟、社會效益的同時,網箱周圍殘餌糞便等有機顆粒物及N、P、Si等無機營養鹽負荷不斷增大,對環境也造成了一定的負面影響,威脅了海水網箱業的可持續發展(張繼紅等, 2013a, b)。本研究結果表明,實驗點馬氏珠母貝的POM攝食總量和對POM的吸收總量均高于對照點,且實驗點異枝麒麟菜的N、P、Si去除總量也高于對照點。馬氏珠母貝可作為生物濾器有效去除網箱養殖產生的殘餌、糞便等顆粒有機物,濾食性貝類主要通過濾食的方式直接影響水體中的營養物質,通過濾食水體中的顆粒物并將其重新包裝成糞便和假糞,從而降低了水體中營養物質的濃度,同時,提高了養殖海域透明度,但濾食性貝類及網箱魚類在生長代謝過程中,會利用呼吸作用釋放CO2和NH4+-N,在IMTA系統中會降低水中的DO水平及升高DIN濃度,對貝類和魚類的生長產生負面影響。而本研究通過引入異枝麒麟菜這一藻類養殖單元,利用藻類的生物學特性,吸收并儲存網箱養殖產生的N、P、Si營養鹽,減輕了網箱養殖對環境的壓力,提高了養殖品種的多樣性和經濟效益對網箱養殖水域的生態環境優化作用顯著。
在IMTA系統中,將POM、營養鹽作為魚貝、魚藻和貝藻之間互利關系的紐帶已成為共識(Hayashi2008; 張繼紅等, 2013a,b;高樺楠等, 2016),魚貝藻綜合養殖系統中,投餌性養殖單元(主要為魚類)產生的殘餌、糞便和營養鹽等物質可轉化為非投餌性(濾食性貝類、藻類等)養殖單元的營養物質來源(唐啟升等, 2013)。本研究結果表明,與貝藻混養相比,網箱區馬氏珠母貝對養殖海區POM的攝食量和吸收量均提高了30.69%,異枝麒麟菜對海區DIN營養鹽的去除量提高了12.64%,同時,貝藻將系統內的多余營養物質轉化為自身的生物質,提高了養殖品種的多樣性和經濟效益,本研究為建立淺海魚貝藻IMTA技術提供了數據支持和理論支撐。
ASHKENAZI D Y, ISRAEL A, ABELSON A. A novel two-stage seaweed integrated multi-trophic aquaculture. Reviews in Aquaculture, 2019, 11(1): 246–262
CHEN P, CHEN Y, MA J,. Bottom sowing culture technology ofin Xisha Reef area. Journal of Hainan Tropical Ocean University, 2018, 25(5): 13–17 [陳攀, 陳燕, 馬軍, 等. 西沙礁盤區瓊枝麒麟菜底播養殖技術研究. 海南熱帶海洋學院學報, 2018, 25(5): 13–17]
CHOPIN T, BUSCHMANN A H, HALLING C,. Integrating seaweeds into marine aquaculture systems: A key towards sustainability. Journal of Phycology, 2001, 37(6): 975–986
DENG Z H, JIANG S, ZHANG B,. Ingestion and digestion of pearl oyster () on microalgae of different types and concentrations. South China Fisheries Science, 2016, 12(3): 112–118 [鄧正華, 姜松, 張博, 等. 合浦珠母貝對不同種類及濃度的單胞藻攝食與消化效果研究. 南方水產科學, 2016, 12(3): 112–118]
FANG Z, HUANG H Q, BAO S X,. The preliminary study ofon absorption of nitrogen and phosphorus in sea water. Journal of Aquaculture, 2008, 29(6): 1–4 [方哲, 黃惠琴, 鮑時翔, 等. 異技麒麟菜對海水中氮、磷吸收的初步研究. 水產養殖, 2008, 29(6): 1–4]
FU S, DENG C M, LIANG F L. Mariculture ofand cage improvement for pearl cultivation. Marine Sciences, 2001, 25(3): 23–24 [符韶, 鄧陳茂, 梁飛龍. 馬氏珠母貝養殖與育珠籠具的改進. 海洋科學, 2001, 25(3): 23–24]
GAO H N, LI L, ZHENG X,. Three co-culture patterns ofand macroalgae in circulating aquaculture system. Journal of Tropical Biology, 2016, 7(2): 147–152 [高樺楠, 李樂, 鄭興, 等. 循環水系統內馬氏珠母貝與3種大型藻類的混養狀況及水質分析. 熱帶生物學報, 2016, 7(2): 147–152]
GAO Z K , ZHANG J H , LI M,. Effects of Temperature fluctuation on physiological and immune parameters of scallop (). Progress in Fishery Sciences, 2017, 38(3): 148–154 [高振錕, 張繼紅, 李敏, 等. 溫度波動對不同規格蝦夷扇貝()生理和免疫指標的影響. 漁業科學進展, 2017, 38(3): 148–154]
GUI J F, TANG Q S, LI Z J,Aquaculture in China success stories and modern trends. Wiley Blackwell, 2018
HAYASHI L, YOKOYA N S, SéRGIO O,Nutrients removed by(Rhodophyta, Solieriaceae) in integrated cultivation with fishes in re-circulating water. Aquaculture, 2008, 277(3/4): 185–191
KASIM M, MUSTAFA A. Comparison growth of,(Rhodophyta, Solieriaceae) cultivation in floating cage and longline in Indonesia. Aquaculture Reports, 2017, 6: 49–55
LAI Z X, LIU Y, WANG Q H,Cloning of FBP gene inand its response to temperature stress. Progress in Fishery Sciences, 2019, 40(2): 106–114 [賴卓欣, 劉雅, 王慶恒, 等. 馬氏珠母貝() FBP基因的克隆及其對溫度脅迫的響應. 漁業科學進展, 2019, 40(2): 106–114]
NEORI A, CHOPIN T, TROELL M,. Integrated aquaculture: rationale, evolution and state of the art emphasizing seaweed biofiltration in modern mariculture. Aquaculture, 2004, 231(1/2/3/4): 361–391
QIAN P Y, WU C Y , WU M ,. Integrated cultivation of the red algaand the pearl oyster. Aquaculture, 1996, 147(1/2): 21–35
TANG Q S, FANG J G, ZHANG J H,. Impacts of multiple stressors on coastal ocean ecosystems and Integrated Multi-trophic Aquaculture. Progress In Fishery Sciences, 2013, 34(1): 1–11 [唐啟升, 方建光, 張繼紅, 等. 多重壓力脅迫下近海生態系統與多營養層次綜合養殖.海洋水產研究, 2013, 34(1): 1–1-1]
WANG A M, SHI Y H, WANG Y,. Biology and new farming techniques ofBeijing: China Agricultural Science and Technology Press, 2010 [王愛民, 石耀華, 王嫣等.馬氏珠母貝生物學與養殖新技術. 北京: 中國農業科學技術出版社, 2010]
WANG D L. Application and research of the co-culture technique of shellfish and seaweed in China. Journal of Oceanography of Huanghai and Bohai Seas, 2001, 19(1): 78–81 [王德利.貝藻混養技術在我國海水養殖中的應用與研究.海洋科學進展, 2001, 19(1): 78–81]
WU M, MAK S K K, ZHANG X J,The effect of co-cultivation on the pearl yield of(Dumker). Aquaculture, 2003, 221(1/2/3/4): 347–356
WU C M, GAO H F, DING M L,. The growth promoting effect of metabolites ofon, Oceanologia et Limnologia Since, 1997, 28(5): 453–457 [吳汪黔生, 高洪峰, 丁美麗, 等. 合浦珠母貝代謝產物對異枝麒麟菜生長的促進作用. 海洋與湖沼, 1997, 28(5): 453–457]
WU W G, ZHANG J H, LIU Y,Size-fractionated phytoplankton and its influencing factors in autumn near Zhangzidao Island and its adjacent waters. Acta Ecologica Sinica, 2018, 38(4):1418–1426 [吳文廣, 張繼紅, 劉毅, 等. 獐子島及鄰近海域秋季浮游植物的粒級結構及其影響因素. 生態學報, 2018, 38(4): 1418–1426]
WU W G, ZHANG J H, WANG W,.Variation of Chl-concentration and its control factors pre-and-post-the- harvest of kelp in Sanggou Bay. Journal of Fisheries of China, 2015, 39(8): 1178–1186 [吳文廣, 張繼紅, 王巍, 等. 桑溝灣水域葉綠素在海帶收獲前后的變化及其影響因素. 水產學報, 2015, 39(8): 1178–1186]
YANG H S, ZHOU Y, WANG J,A modelling estimation of carrying capacities for Chlamys farreri, Laminaria japonica and Apostichopus japonicus in Sishiliwan Bay, Yantai, China. Journal of Fishery Sciences of China, 2000, 7(4): 27–31 [楊紅生, 周毅, 王健, 等. 煙臺四十里灣櫛孔扇貝、海帶和刺參負荷力的模擬測定. 中國水產科學, 2000, 7(4): 27–31]
ZHANG J H, WU T, XU D,. Feeding on feces, diets and cage farming sediments of Paralichthys olivaceus by Japanese scallop. Progress In Fishery Sciences, 2013a, 34(2): 96–102 [張繼紅, 吳桃, 徐東, 等. 蝦夷扇貝對牙鲆的糞便、殘餌及網箱養殖區沉積物的攝食. 漁業科學進展, 2013a, 34(2): 96–102]
ZHANG J H, WU T, GAO Y P,Feeding behavior of 5 species filter-feeding bivalves onfeed, fecal and sediment particulates in cage farming area. Journal of Fisheries of China, 2013b, 37(5): 90–97 [張繼紅, 吳桃, 高亞平, 等. 5種濾食性貝類對牙鲆的糞便、殘餌及網箱養殖區沉降物的攝食行為. 水產學報, 2013b, 37(5): 90–97]
ZHANG J H, REN L H, WEI Y W,Ecological environment and sustainable utilization of biological resources in Sanggou Bay. Beijing: Agricultural Press of China, 2018 [張繼紅, 任黎華, 魏龑偉, 等. 桑溝灣生態環境與生物資源可持續利用. 北京: 中國農業出版社, 2018]
ZHOU Y, YANG H S. Biodeposition method used in bivalval physio-ecological studies. Chinese Journal of Ecology, 2002, 21(6): 74–76 [周毅, 楊紅生. 應用于貝類生理生態學研究的生物沉積法. 生態學雜志, 2002, 21(6): 74–76]
Analysis of the Effectiveness of the Polyculture of Shellfish and Algae and Influencing Factors in the Xuejiadao Sea Cage Aquaculture Area
WU Wenguang1,3, ZHANG Jihong1,2①, LIU Yi1, ZHAO Yunxia1, ZHANG Kunyang1, WANG Liangliang1
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Qingdao, Shandong 266071, China;2. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao, Shandong 266071, China; 3. College of Fisheries and Life Sciences, Shanghai Ocean University, Shanghai 201306, China)
In recent years, environmental pollution and ecological problems caused by cage aquaculture have become increasingly prominent in shallow seas. The cultivation technique of integrated multi-trophic aquaculture (IMTA) can be applied to minimize the impact of nutrient enrichment from organic fisheries. The IMTA mode was implemented in September to November 2018 in this study, which included fish cage aquaculture, the shellfish, and the alga. The feeding physiology indicators ofand the nutrient uptake rate ofwere measured at different temperatures in the laboratory concurrent with conducting field experiments, and the improvement of the water quality in the cage aquaculture area by IMTA of shellfish and algae were compared and analyzed. The results showed that: (1) the specific growth rate (SGR) ofin the marine fish cage area was significantly higher than that in control area, and the average shell height ofin the 0.50~0.62 m water layer in the cage area was significantly higher than the other water layers. (2)had the fastest SGR in the 0.60~0.90 m water layer, and the net wet weight gain ofwas significantly higher than that in the control area. (3) At 24.45℃, the average feeding rate, absorption rate, and absorption efficiency ofon particulate organic matter were, respectively, (4.13±0.77) mg/h·ind., (1.04±0.24) mg/h·ind., and (25.00±2.51)%, which were significantly higher than those at 20.74℃. Within the experimental temperature range, the absorption rates of nitrogen, phosphorus, and silicon first increased then decreased with increasing temperature, and the maximum values appeared at 30℃. (4) It is estimated that the total amount of filtered particulate organic matter and the total amount of absorbed particulate organic matter byin the marine fish cage area during the culture experiment was 778.08 g and 144.47 g, respectively, and the total amount of nitrogen, phosphorus, and silicon removed byin the marine fish cage area was 8.55 g, 1.11 g, and 9.18 g, respectively; all the above indicators were higher than those in the control area. Temperature, depth of cultivation, particulate organic matter, and nutrient concentrations were the primary factors affecting the growth and physiological metabolism ofand. This study provides data and theoretical support for the establishment of IMTA technology.
Sea cage mariculture;;; Integrated multi-trophic aquaculture (IMTA)
S966
A
2095-9869(2022)03-0146-10
10.19663/j.issn2095-9869.20210201001
http://www.yykxjz.cn/
吳文廣, 張繼紅, 劉毅, 趙云霞, 張坤陽, 王亮亮. 薛家島海水網箱養殖海域貝藻綜合養殖效果及其影響因素分析. 漁業科學進展, 2022, 43(3): 146–155
WU W G, ZHANG J H, LIU Y, ZHAO Y X, ZHANG K Y, WANG L L. Analysis of the effectiveness of the polyculture of shellfish and algae and influencing factors in the Xuejiadao sea cage aquaculture area. Progress in Fishery Sciences, 2022, 43(3): 146–155
ZHANG Jihong, E-mail: zhangjh@ysfri.ac.cn
* 農業農村部農業國際合作交流項目(“一帶一路”熱帶國家水產養殖科技創新合作) (20180101)、山東省重大科技創新工程專項——“健康海洋”技術創新工程(2018SDKJ0501-3)和中國科學院戰略性先導科技專項(A類)“美麗中國生態文明建設科技工程”(XDA23050402)共同資助 [This work was supported by Aquaculture Science and Technology Cooperation Projects of Tropical Countries Along the “Belt and Road” of the Ministry of Agriculture (20180101), the Marine Science and Technology Fund of Shandong Province for Pilot National Laboratory for Marine Science and Technology (Qingdao) (2018SDKJ0501-3), and the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA23050402)].吳文廣,E-mail: wuwg@ysfri.ac.cn
張繼紅,研究員,E-mail: zhangjh@ysfri.ac.cn
2021-02-01,
2021-03-16
(編輯 陳 嚴)
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
