核轉錄因子NFYB和NFYC促進紅系終末分化
2022-06-27 01:59:08余東林陰佳瀅劉雪會
基礎醫學與臨床 2022年6期
余東林,楊 希,陰佳瀅,劉雪會,呂 湘
(中國醫學科學院基礎醫學研究所 北京協和醫學院基礎學院 病理生理學系 醫學分子生物學國家重點實驗室,北京 100005)
紅細胞是血液中含量極其豐富的細胞類型,具有攜氧能力,對于維持生命至關重要。紅系終末分化是產生成熟紅細胞的前置步驟,依次經歷原幼紅(proerythroblast,ProE)、早幼紅(basophilic erythro-blast,BasoE)、中幼紅(polychromatophilic erythro-blast,PolyE)、晚幼紅(orthochromatic erythroblast,OrthoE) 細胞階段,經脫核產生網織紅細胞,進一步在血循環中形成成熟紅細胞[1]。紅系相關轉錄因子對于促進紅系終末分化成熟具有重要作用,如KLF1、FOXO3、E2F2、NFE2等,其表達異常或缺失導致紅系終末分化受阻或抑制脫核引發貧血[2-4]。因此篩選紅系終末分化的關鍵轉錄調控因子有助于尋找貧血的治療靶點。
利用單細胞轉錄組圖譜可精確刻畫細胞分化軌跡和轉錄調控網絡。近期發表的人與小鼠的紅系單細胞轉錄組圖譜可幫助揭示紅系終末分化的轉錄調控機制[5]。SCENIC(single-cell regulatory network inference and clustering)分析是目前較為公認的分析轉錄因子的方法[6],優勢在于在轉錄因子和靶點基因共表達分析的基礎上結合Motif富集分析,從而更加可靠地預測出轉錄因子主導的基因調控網絡。
本研究對人紅系終末分化的單細胞轉錄組進行SCENIC分析,并對預測所得的核轉錄因子NFYB和NFYC進行驗證,初步探究其在紅系終末分化中的表達模式及調控作用。
1 材料與方法
1.1 材料
細胞培養基IMDM、DMEM、胎牛血清和HEPES(Gibco公司);實驗所需質粒由暨南大學鞠振宇實驗室惠贈;T4 DNA連接酶及T4 DNA連接酶緩沖液(BioLabs公司);感受態細胞DH5α(北京全式金生物技術有限公司);C57BL/6 小鼠(北京維通利華實驗動物技術有限公司);EcoRⅠ/XhoⅠ內切酶(NEB 公司);反轉錄試劑盒(南京諾唯贊生物科技股份有限公司);膠回收試劑盒(Thermo Fisher Scientific公司);轉染試劑p4000(北京蘭博利德商貿有限公司); TER119磁珠(Miltenyi公司);TER119-PE抗體、CD71-APC抗體和FcR 封閉抗體(BD公司);Hoechst 33342和F-108(Sigma-Aldrich公司)。
1.2 方法
1.2.1 構建質粒:借助網站(http://splashrna.mskcc.org/)設計Nfyb和Nfyc的shRNA序列各3條,shLuc:5′-TTAATCAGAGACTTCAGGCGGT-3′,shNfyb-1:5′-TTTTGAACCATTTGTATCTTCA-3′,shNfyb-2:5′-TCTTCATGATCATTCATGCTGT-3′,shNfyb-3:5′-TCA CATAATGACTTCCTCCGAT-3′,shNfyc-1:5′-TTCA TAATCTTCTTAATACGAG-3′,shNfyc-2:5′-TAATCT TCTTAATACGAGCCAG-3′,shNfyc-3:5′-TTTCATA ATCTTCTTAATACGA-3′。合成相應序列后退火合成雙鏈DNA,與酶切后的SFFV-GFP-miR30 載體質粒連接,測序驗證插入序列的準確性。
1.2.2 包裝病毒:將HEK-293T細胞培養于含10%胎牛血清的DMEM培養基。加入轉染試劑P4000、病毒包裝質粒psPAX2、pMD2.G和相應的干擾質粒后48、72 h分別收集培養基,過濾后超高速離心得到病毒沉淀,用胎肝增殖培養基重懸保存于-80℃冰箱備用。
1.2.3 小鼠胎肝細胞體外紅系分化: 取小鼠E14.5的胎肝,使用1 mL注射器將肝臟吹成單細胞懸液,磁珠分選獲得TER119陰性細胞,增殖培養3 d,分化培養3 d[7]。
1.2.4 感染病毒:在體外培養的小鼠胎肝細胞增殖期第1天向細胞中加入F-108和HEPES,顛倒混勻后分孔,分別加入相應的慢病毒,24 h后換液。
1.2.5 熒光激活細胞分選技術分選與檢測:小鼠胎肝TER119陰性細胞體外誘導紅系分化3 d,取1×107個細胞加入0.5 μL FcR抗體封閉,4 ℃ 15 min,加入TER119-PE 1 μL和CD71-APC(稀釋5倍)1 μL,4 ℃ 30 min。使用索尼MA900流式細胞分選儀收集紅系終末分化各個時期的(R2~R5)細胞。對基因干擾和對照組細胞,在上述抗體孵育后繼續加入Hoechst進行核染色,使用MA900檢測紅系分化效果和脫核率,FlowJo軟件分析結果。
1.2.6 RT-qPCR檢測mRNA: 使用Trizol提取RNA,反轉錄得到cDNA后進行qPCR檢測。Nfyb:正向引物5′-GCCTCCCAGCTAGGGATTTC-3′,反向引物5′-TTCCTGTTTGAGGTATGGCATTT-3′。 Nfyc:正向引物5′- TGAAACCTCCAAAGCGCCA-3′,反向引物5′- GTGGTCTGACCCTGCTGAG-3′。 Actin:正向引物5′-CCTTCCTTCTTGGGTATGGAATC-3′,反向引物5′-AGCACTGTGTTGGCATAGAGGT-3′。 Hbb:正向引物5′-CCAGCCTCAGTGAGCTCCACT-3′,反向引物5′- GGCCCAGCACAATCACGAT-3′。
1.3 統計學分析
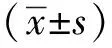
1.4 生物信息分析
1.4.1 Bulk轉錄組分析:GEO數據庫下載人類和小鼠的紅系終末分化各個時期的bulk轉錄組數據集(GSE53983),使用cutadapt和trimmomatic軟件去除接頭和低質量序列,通過hisat2軟件分別比對到人類基因組hg19和小鼠基因組mm10,得到sam文件,并使用samtools軟件壓縮為bam文件,使用featureCounts軟件得到基因表達矩陣,最后使用R軟件進行后續分析。
1.4.2 單細胞轉錄組分析:GEO數據庫下載數據集(GSE150774),使用Seurat包分別處理3個人類骨髓細胞的單細胞轉錄組表達矩陣,經過無監督細胞聚類,歸一化,降維分析等處理后合并。使用rSuperCT包預測細胞類型,只保留紅細胞,并使用DoubletFinder包去除doublet,最終保留9 277個細胞用于后續的分析。使用monocle3包進行擬時序分析,從而確定分化的時序關系。
1.4.3 SCENIC分析: 使用SCENIC包用于預測基因調控網絡。GENIE3包分析轉錄因子和靶基因的共表達情況;RcisTarget包進行Motif分析,將不具有共同Motif的靶基因刪除,留下的靶基因與轉錄因子作為一個調節單元,即regulon;AUCell包計算每個regulon在每個細胞的活性打分。
1.4.4 代碼可用性:代碼已經上傳于GitHub(https://github.com/yudonglin506311858/nfyb_project)。
2 結果
2.1 人骨髓紅系細胞單細胞轉錄組圖譜分析提示NFYB及NFYC因子參與紅系終末分化調控
利用人類骨髓紅系終末分化細胞的單細胞轉錄組數據集(GSE150774)[8],使用Seurat包和monocle3包進行分析,確定分化階段ProE/BasoE、 Early-PolyE、 Late-PolyE、 Early-OrthoE和 Late-OrthoE(圖1A~B)。使用SCENIC預測調控網絡單元(regulon)[6]。根據regulon在每個細胞亞群的特異性分數進行排序(圖1C~D),NFY轉錄因子復合物的兩個亞基NFYB和NFYC均被預測為紅系終末分化早期的調控因子且特異性較高,其regulon活性和RNA表達水平在紅系分化較早期的ProE/BasoE和 Early-PolyE中最高(圖1E~F)。
2.2 Nfyb和Nfyc在人和小鼠紅系終末分化不同時期的表達情況
進一步利用bulk轉錄組[9]分析Nfyb和Nfyc在紅系終末分化各個時期的表達情況。人骨髓細胞紅系終末分化早期高表達NFYB和NFYC(圖2A),進入OrthoE后顯著下降;而在小鼠骨髓紅系細胞的表達模式稍有不同(圖2B),Nfyb在PolyE時期有更高的表達,Nfyc則在OrthoE表達顯著降低。對小鼠胎肝細胞進行體外紅系誘導分化,可通過CD71與TER119信號強度將細胞分為R1-R5的5個漸次成熟的細胞群(圖2C),Nfyb和Nfyc在終末分化較早期的R3-R4表達上調,隨后表達下降(圖2D),與小鼠骨髓的轉錄組數據表現一致。
2.3 敲低Nfyb和Nfyc抑制血紅蛋白基因的表達
利用小鼠胎肝體外紅系分化體系進行Nfyb和Nfyc的功能研究。兩個基因各3條特異性shRNA序列均可在RNA水平顯著降低Nfyb或Nfyc的表達(圖3A~B)。進一步通過qPCR技術檢測紅系終末關鍵的血紅蛋白Hbb基因表達,靶向兩個基因的3條shRNA序列均可顯著下調Hbb的RNA表達水平,提示抑制Nfyb和Nfyc可影響紅系終末分化(圖3C~D)。
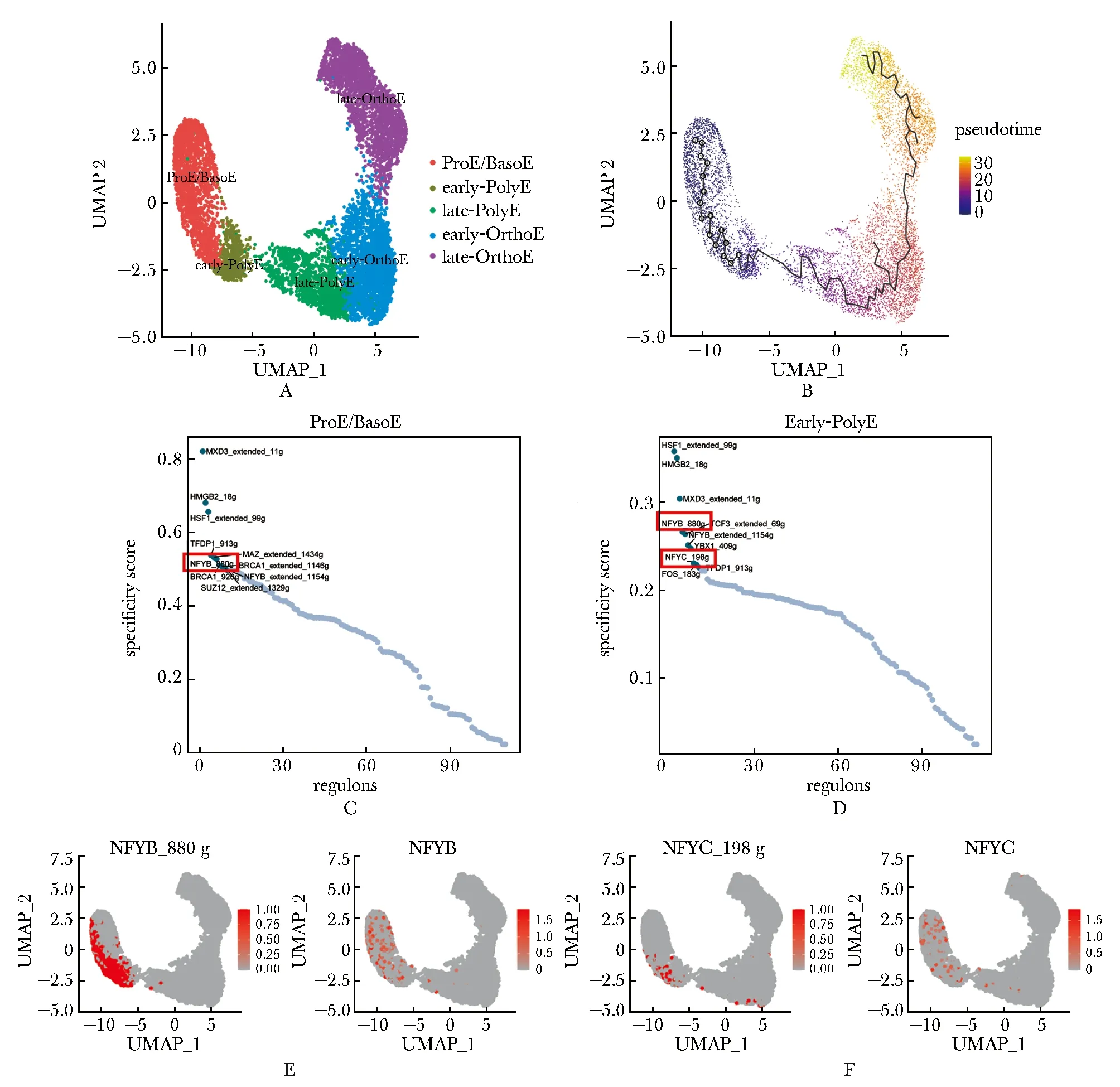
A.UMAP projection of human terminal erythropoiesis, cell types colored by different colors; B.pseudo-time analysis; C,D.regulon specificity score of different regulons in “ProE/BasoE” and “Early-PolyE” cells; E,F.activation (left) and expression (right) of NFYB (E) and NFYC (F) were indicated by red color
2.4 敲低Nfyb和Nfyc抑制紅系終末分化及脫核
敲低Nfyb和Nfyc后,流式細胞分析顯示紅系分化較早期R3和R4比例增加,晚期R5比例顯著下降(圖4A~B),且紅細胞脫核率顯著降低(圖4A,C),說明抑制Nfyb和Nfyc使紅系終末分化阻滯在較早期。
3 討論
本研究主要基于紅系終末分化單細胞轉錄組數據進行轉錄調控因子預測,并結合bulk轉錄組和功能實驗初步探究了轉錄因子NFYB和NFYC在紅系終末分化中的表達及作用。結果表明NFYB和NFYC在人與小鼠紅系終末分化較早期的BasoE和PolyE高表達,分別敲低后使紅系終末分化阻滯在早期,表明NFYB和NFYC是紅系終末分化早期的調控因子。
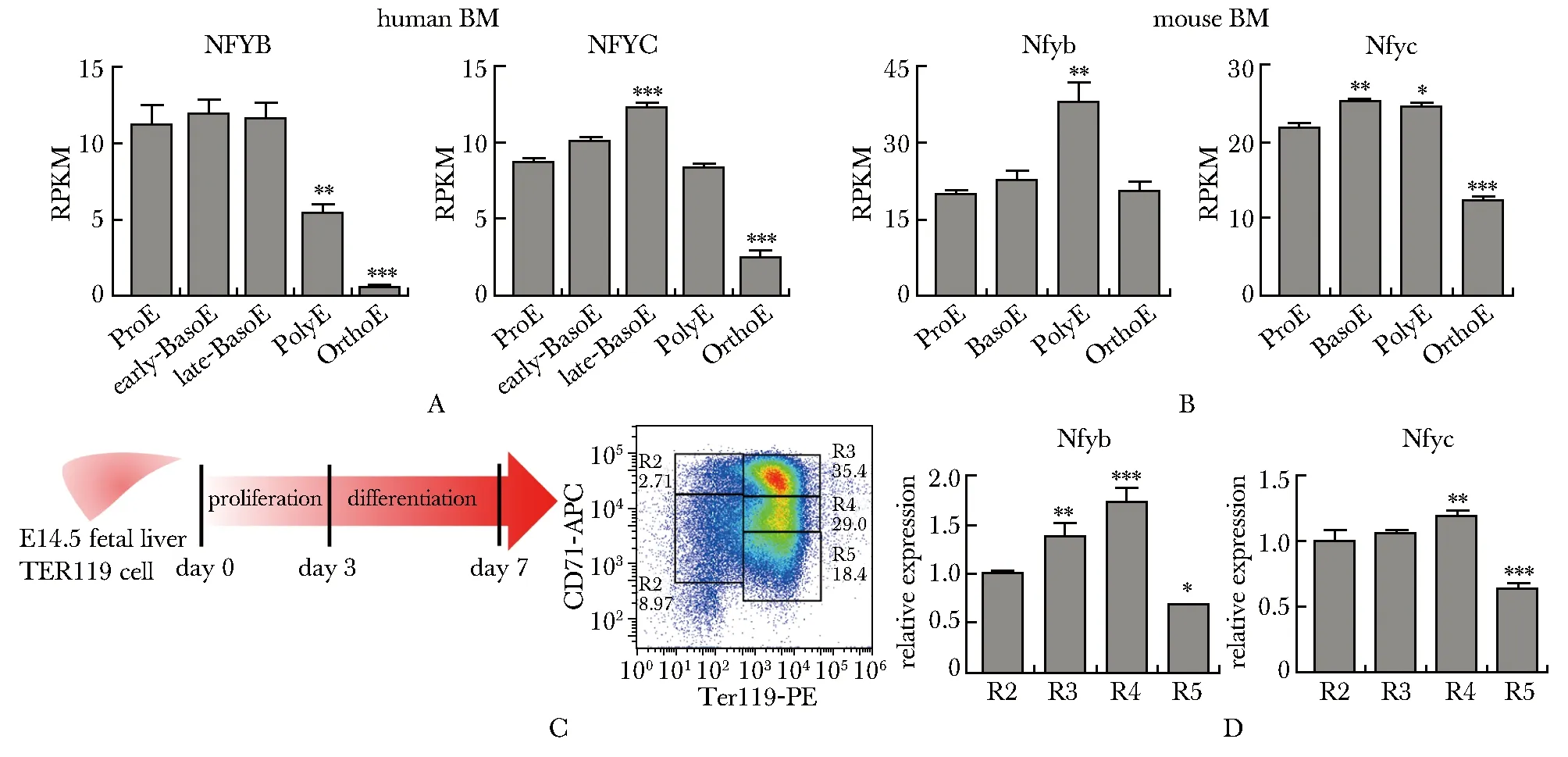
A,B.expression levels of NFYB and NFYC in human (A) and mouse (B) terminal erythropoiesis were identified in published bulk transcriptional sequencing data(GSE53983); C.flowchart of mouse fetal liver in vitro erythroid differentiation and the soring of cells at different maturation stages (R1-R5) by flow cytometry;D.expression levels of NFYB and NFYC at different stages of terminal erythroid differentiation of mouse fetal liver; *P<0.05, **P<0.01, ***P<0.001 compared with ProE or R2
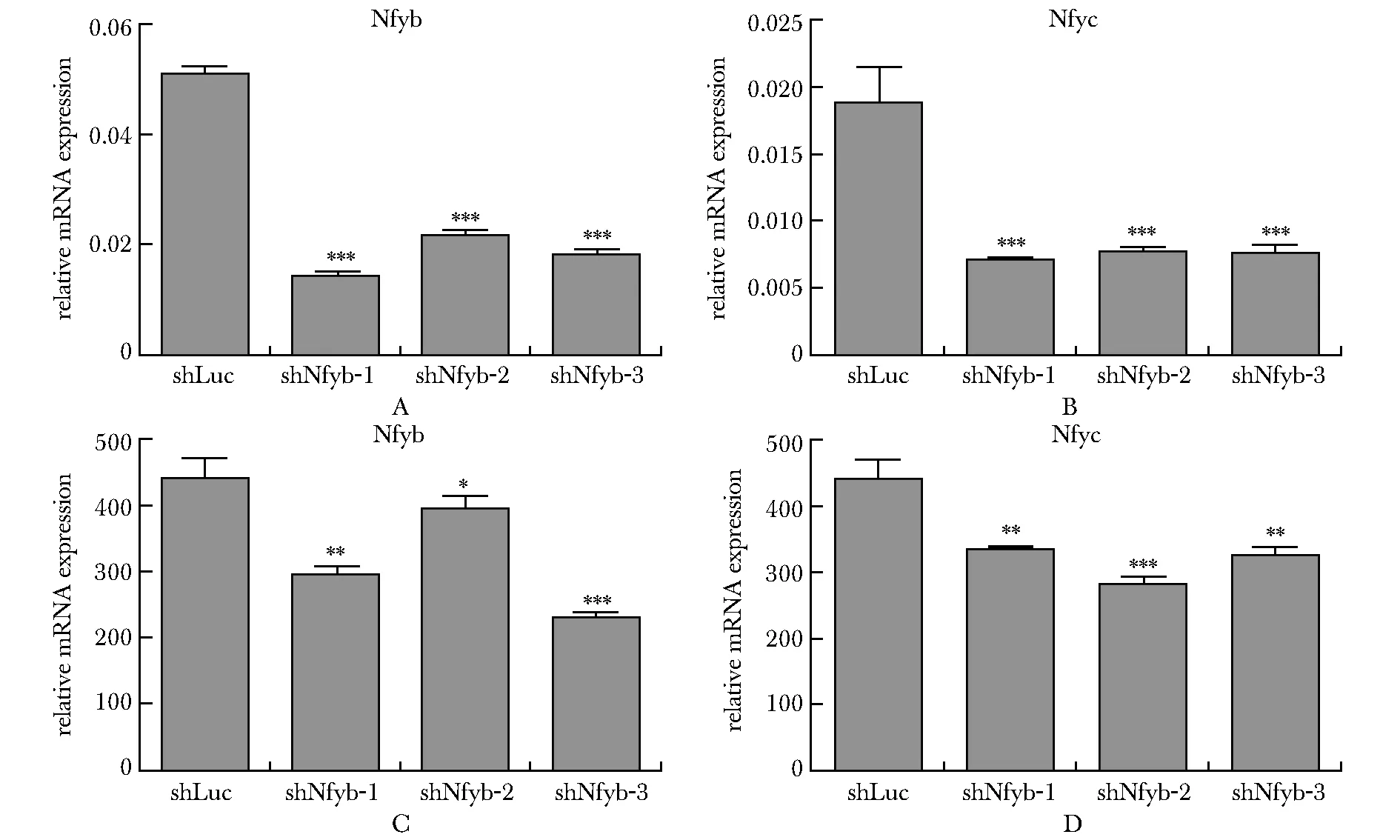
A,B.knockdown efficiency of Nfyb and Nfyc;C,D.RT-qPCR detection of hemoglobin gene (Hbb) expression level;*P<0.05, **P<0.01, ***P<0.001 compared with shLuc group

A.representative flow cytometry plots showing the effects of Nfyb and Nfyc knockdown on erythroid differentiation and enucleation; B.knockdown of Nfyb and Nfyc blocked erythroid terminal differentiation; C.knockdown of Nfyb and Nfyc decreased enucleation rate; *P<0.05, **P<0.01, ***P<0.001 compared with shLuc group
NF-Y復合物由NFYA、NFYB和NFYC組成,其中NFYA識別啟動子中CCAAT 框的序列,NFYB和NFYC形成類核小體結構[10],招募 TATA 盒結合蛋白(TBP)和RNA聚合酶II到啟動子[11]。NF-Y通過直接結合CCAAT 框激活γ-珠蛋白[12],參與珠蛋白的轉錄[13]。此外,NF-Y是造血干細胞增殖和存活所必需的[14],還具有調節線粒體活性的功能[15]。關于NF-Y在紅系終末分化中調控的分子機制尚有待進一步研究。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
電子制作(2018年18期)2018-11-14 01:48:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
終身教育研究(2014年5期)2014-02-28 01:23:06
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
