非生長季降水對青藏高原高寒草甸優勢種生物量穩定性的影響
2022-06-28 03:11:32劉琪源董世魁肖建南武勝男周秉榮
生態學報 2022年10期
李 帥,劉琪源,董世魁,肖建南,武勝男,周秉榮,李 甫
1 山西農業大學資源環境學院,太谷 030801 2 北京師范大學環境學院,北京 100875 3 北京林業大學草業與草原學院,北京 100083 4 青海省氣象科研所,西寧 810002
穩定性作為生態學和全球變化生物學中最具影響力的概念之一,是指生態系統在外界干擾(比如氣候變化)下維持自身結構和功能的能力,且一直以來都是研究的熱點[1—5]。穩定性是對種群和群落相互之間以及對環境變化的反應的綜合衡量,盡管有許多方法可以測量穩定性,但一般采用生態變量(如生物量)的均值除以標準差來表征穩定性[1,6]。穩定性不僅可以反映外界干擾強度,而且對生態系統的功能和服務價值(如養分循環、生物量生產、畜牧產業的可持續性等)的維持具有很重要的作用[7—8]。
“質量比”假說認為,群落穩定性和生態系統功能在很大程度上取決于優勢物種[9—14],群落間種間相互作用的強度也取決于優勢物種[14—15]。有研究表明優勢種的某些特征,比如數量可能會影響物種間的相互作用[16]。于此同時越來越多的研究表明,次優勢物種可能也很重要[16—19]:比如有研究指出,相比于優勢物種,次優勢物種不僅直接響應氣候變化,而且還可以間接對優勢種產生影響[20—22]。
氣候變化導致的氣溫升高和降水模式的變化將會對群落組成和生態系統功能產生重要影響[23—27]。根據IPCC的報告,全球平均升溫的趨勢為0.03—0.06℃/10a[23],并且預計會在未來進一步上升。處于高緯度區域的青藏高原氣候變化會更加劇烈:有證據表明青藏高原的升溫速率為0.37℃/10a顯著高于全球速率[28];另外有研究表明在過去的五十年間,青藏高原的降水量增加了0.67mm/a[29],并且也將在未來持續增加[30],因此青藏高原整體呈現暖濕化的趨勢[9,31]。相比于年際之間溫度和降水的變化,生長季和非生長季氣候變化模式的差異可能會對生態系統產生更重要的影響。有研究表明,非生長季的氣候變暖會影響生態系統的穩定性[32—34]。在過去幾十年里,非對稱季節變暖在青藏高原上表現得十分明顯[35—38]。不對稱變暖可能與植物生產與消耗之間的平衡有關,也會導致植物生長過程中的水分有效性降低[39—42]。溫度和降水的季節性不對稱變化會引起物種組成[20,43]、營養循環和相互作用[44]的變化,最終影響生物量或穩定性[45—48]。
青藏高原是世界上最高的高原,平均海拔超過4000m,高寒草甸作為青藏高原最典型的生態系統之一,也對氣候變化十分敏感[49]。但是關于高寒草甸對于氣候變化的響應的研究結果并不一致,一方面這可能與單一的控制性實驗,比如單獨增溫和單獨增水的實驗多于復合型的實驗[50—51],另一方面,鑒于研究年限的差異,短期的控制性實驗得到的結果相比于長時間序列觀測實驗得到的結果還存在差異,這可能會造成對于氣候變化的響應被低估或高估[47,52—53]。關于高寒草甸植物群落對生長季/非生長季不對稱增溫的響應的相關研究時間年限較短(少于5a)[40]。因此,本研究利用青藏高原東部的高寒草甸17年(即2001年至2017年)的觀測數據,研究優勢物種本身的特征(包括高度和多度)以及次優勢物種生物量穩定性,結合生長季/非生長季不對稱增溫和降水,探究對于優勢種的生物量穩定性的影響。
1 材料與方法
1.1 研究區概況
研究區位于青海省海北藏族自治州海晏縣西海鎮(36.93°N,100.95°E,海拔3100 m)。草地類型為高寒草甸,優勢物種為紫花針茅和洽草,植被蓋度大于80%,土壤類型為砂壤土,年均降水量 400 cm,年蒸發量為 1538 mm,年平均氣溫-0.45 ℃。
1.2 實驗設計
從 2001 年至 2017 年,在研究區將地勢平坦的面積 100 m×100 m的區域分為 50 m×50 m的 4塊,每一塊進一步被分為25 m×25 m的 4 個小區,生長季圍封,非生長季放牧,地表無明顯徑流。每年9月底在4塊中每一塊隨機選擇一個小區,小區按順時針方向輪換每四年一輪。從中選取1m×1m的對優勢物種(紫花針茅)及次優勢物種群(洽草)的地上生物量進行監測。同時記錄紫花針茅的多度、高度,收集到的植物樣品在烘箱中高溫(65℃)烘干至恒重。在離最近的海晏縣氣象站取得日平均氣溫以及日降水量。
1.3 統計分析
計算2001年至2017年的年平均氣溫、生長季平均氣溫(4—10月)、非生長季平均氣溫(10月—第二年的4月)、年降水量、生長季降水量(4—10月)、非生長季降水量(10月—第二年的4月)。
計算了紫花針茅和洽草各年處理間的平均生物量(μ)與其處理間的標準差(σ)的比值為生物量穩定性[1,4—6]。分別計算了年平均氣溫、生長季平均氣溫、非生長季平均氣溫、年降水量、生長季降水量、非生長季降水量、紫花針茅的高度、多度、紫花針茅生物量穩定性、洽草生物量穩定性隨時間的變化趨勢;采用線性回歸包括研究了環境因子包括,年平均氣溫、生長季平均氣溫、非生長季平均氣溫、年降水量、生長季降水量、非生長季降水量對生物量穩定性的影響,以及生物因子,包括紫花針茅的高度、多度、洽草生物量穩定性與紫花針茅生物量穩定性的關系。
鑒于2010年紫花針茅生物量穩定性和2007年洽草生物量穩定性的值異常高(與平均值的偏差超過三倍標準差),本研究分別進行了剔除。依據線型回歸找到對穩定性影響較大的因子,考慮到年均溫和生長季平均溫度,以及年降水和生長季降水量呈現顯著的正相關且相關性系數大于0.7,為了避免共線性,本研究只將生長季平均溫度和生長季降水納入到模型的分析之中,并據此用建立理論模型。為了識別環境和生態因素影響植物群落的生物量穩定,本文結合Yang[29]和Ma[5]等已有研究結果構建了的先驗模型——結構方程模型,包括年平均氣溫、生長季平均氣溫、非生長季平均氣溫、年降水量、生長季降水量、非生長季降水量、紫花針茅生物量穩定性、洽草生物量穩定性、紫花針茅高度、紫花針茅多度。利用R語言‘basicTrendline’包來進行簡單線性回歸,利用Amos 21 (IBM, Chicago)進行結構方程模型分析,非顯著χ2檢驗(P>0.05)、擬合優度指數(GFI)(>0.90)和近似均方根誤差(RMSEA)(<0.05)[40]。
2 結果
2.1 氣候因子、生物因子的變化趨勢
2001年到2017年,年平均氣溫以k=0.041的趨勢顯著升高(P<0.05),生長季平均氣溫升高趨勢不顯著(P=0.068),非生長季平均氣溫升高卻不顯著(P>0.05)。年降水量和生長季降水量均增加了100 mm,非生長季降水量沒有明顯的趨勢(圖1)。非生長季降水量的變化趨勢與整個溫度降水上升趨勢不一致,沒有明顯的趨勢;年降水量集中在生長季,生長季降水較多,非生長季降水較少(圖1)。
不同于氣溫與降水變化趨勢,兩種植物的生物量穩定性均呈下降趨勢,其中洽草呈顯著下降趨勢(P<0.05),紫花針茅下降趨勢不顯著(P>0.05);紫花針茅的高度和多度升高緩慢但不顯著(P>0.05)(圖2)。
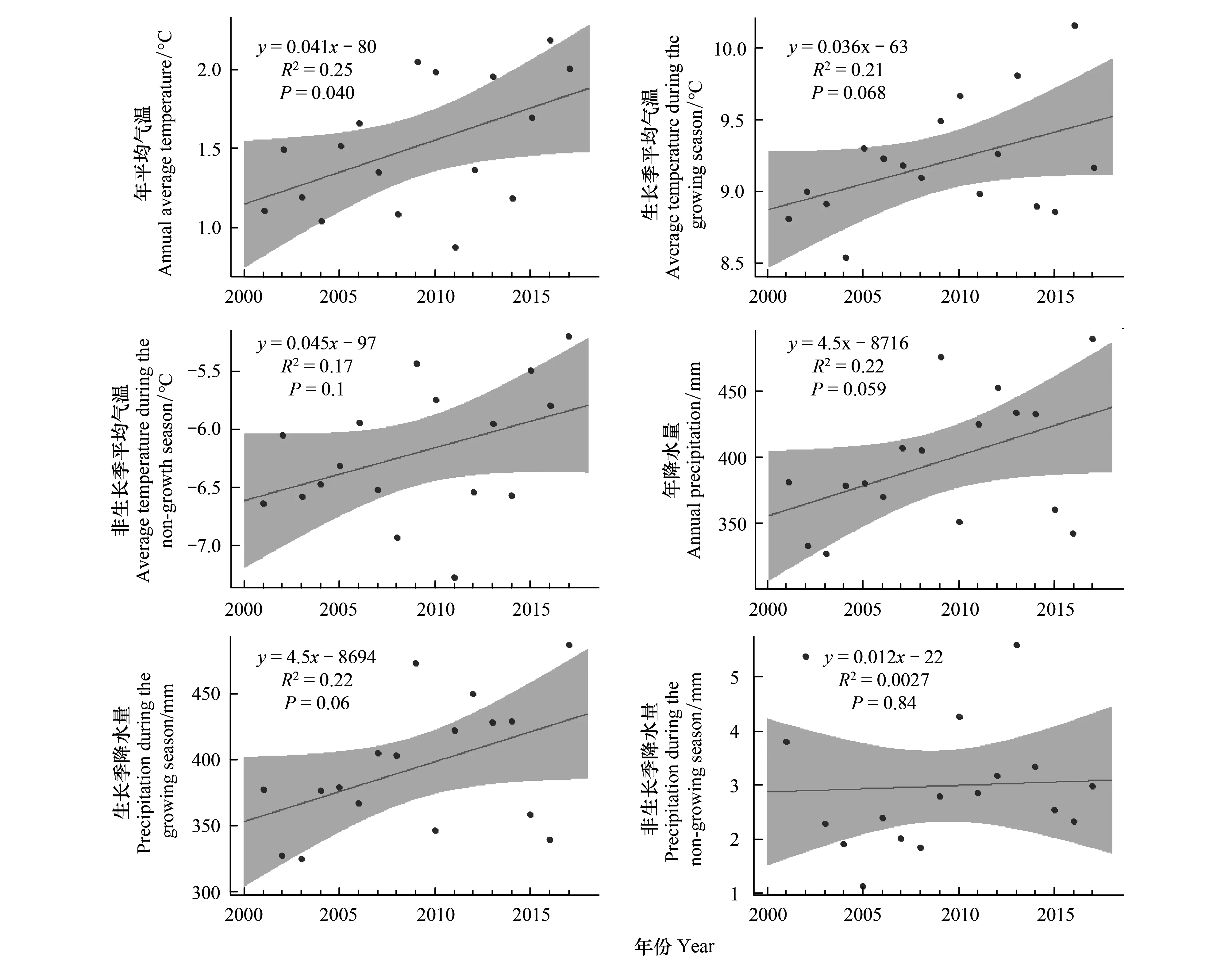
圖1 年平均氣溫、生長季平均氣溫、非生長季平均氣溫、年降水量、生長季降水量、非生長季降水量的變化趨勢Fig.1 The change trend of annual mean temperature, growth season average temperature, non-growth season average temperature, annual precipitation, growth season precipitation, non-growth season precipitation灰色區域是置信水平為95%的置信區間
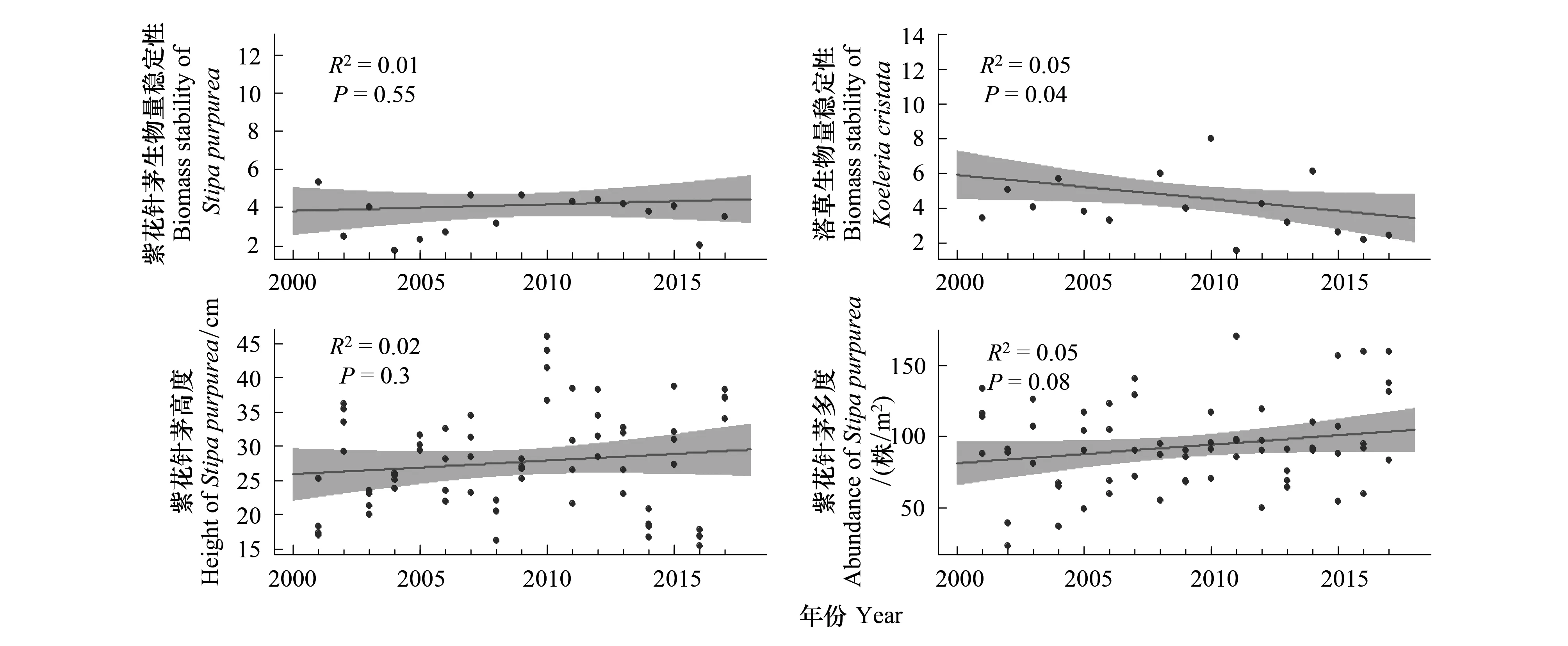
圖2 紫花針茅生物量穩定性、洽草生物量穩定性、紫花針茅高度、紫花針茅多度的變化趨勢Fig.2 The change trend of biomass stability of Stipa purpurea, and Koeleria cristata, the change trend of height and abundance of Stipa purpurea灰色區域是置信水平為95%的置信區間
2.2 環境因子、生物因子與紫花針茅生物量穩定性的關系
線性回歸分析顯示,與紫花針茅生物量穩定性顯著相關的有3個因子,即非生長季年降水、紫花針茅的高度以及洽草的生物量穩定性呈極顯著相關(P<0.05)(圖3)。非生長季降水量增加會促進紫花針茅的生物量穩定性(圖3)。
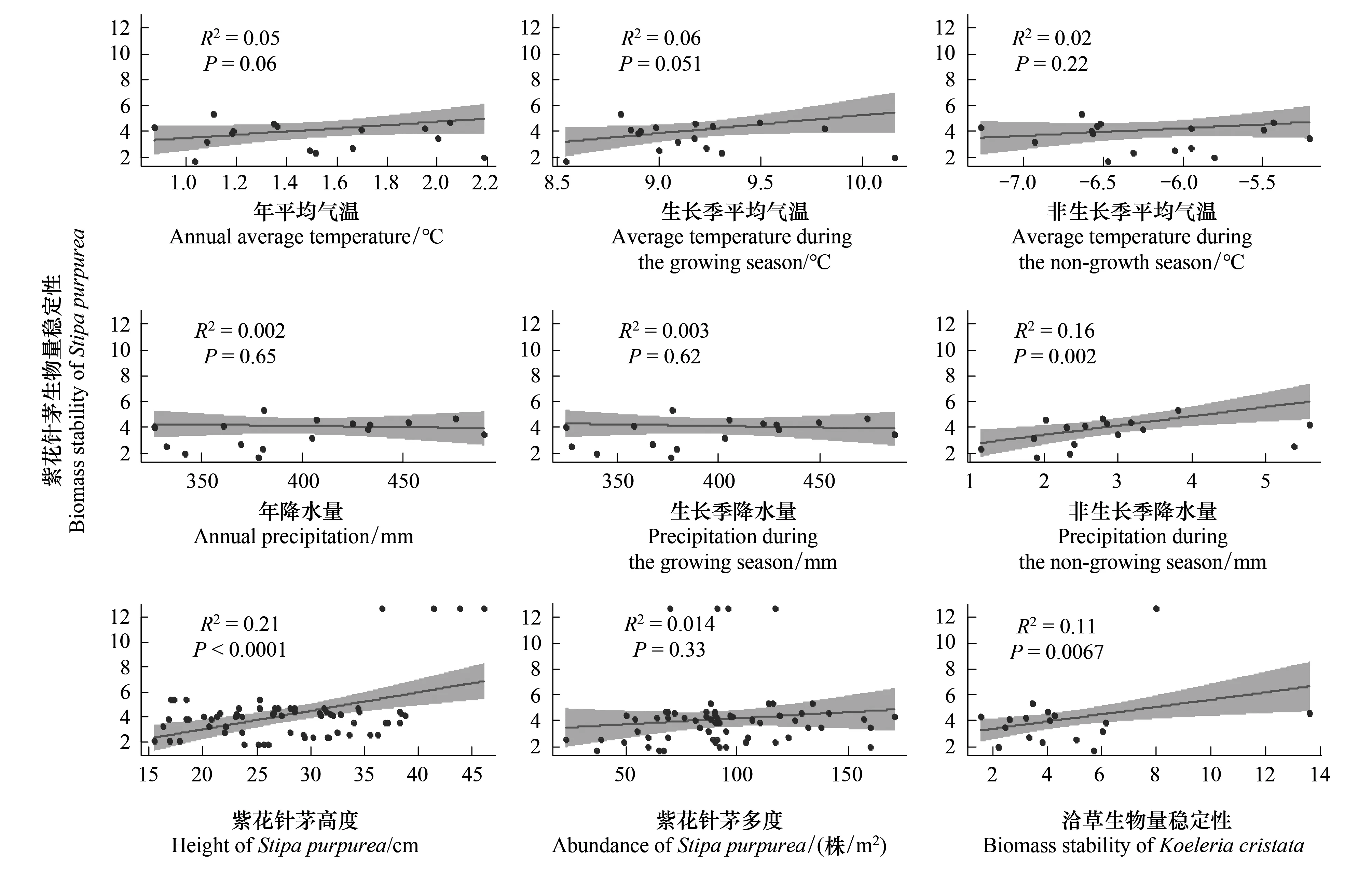
圖3 環境因子、生物因子與優勢種生物量穩定性的關系Fig.3 Relationship between environmental factors, biological factors and biomass stability of dominant species灰色區域是置信水平為95%的置信區間
2.3 環境因子、生物因子對紫花針茅生物量穩定性的影響
通過路徑分析(根據Chi-square=13.00,df=7,P=0.08),紫花針茅的生物量穩定被非生長季降水、紫花針茅的高度和多度以及洽草的生物量穩定性解釋了45%,其中高度、非生長季降水和洽草的生物量穩定性的貢獻最大。紫花針茅的高度和多度,以及洽草的生物量穩定性都與生長季/非生長季溫度和降水的變化,沒有顯著的關系(圖4)。
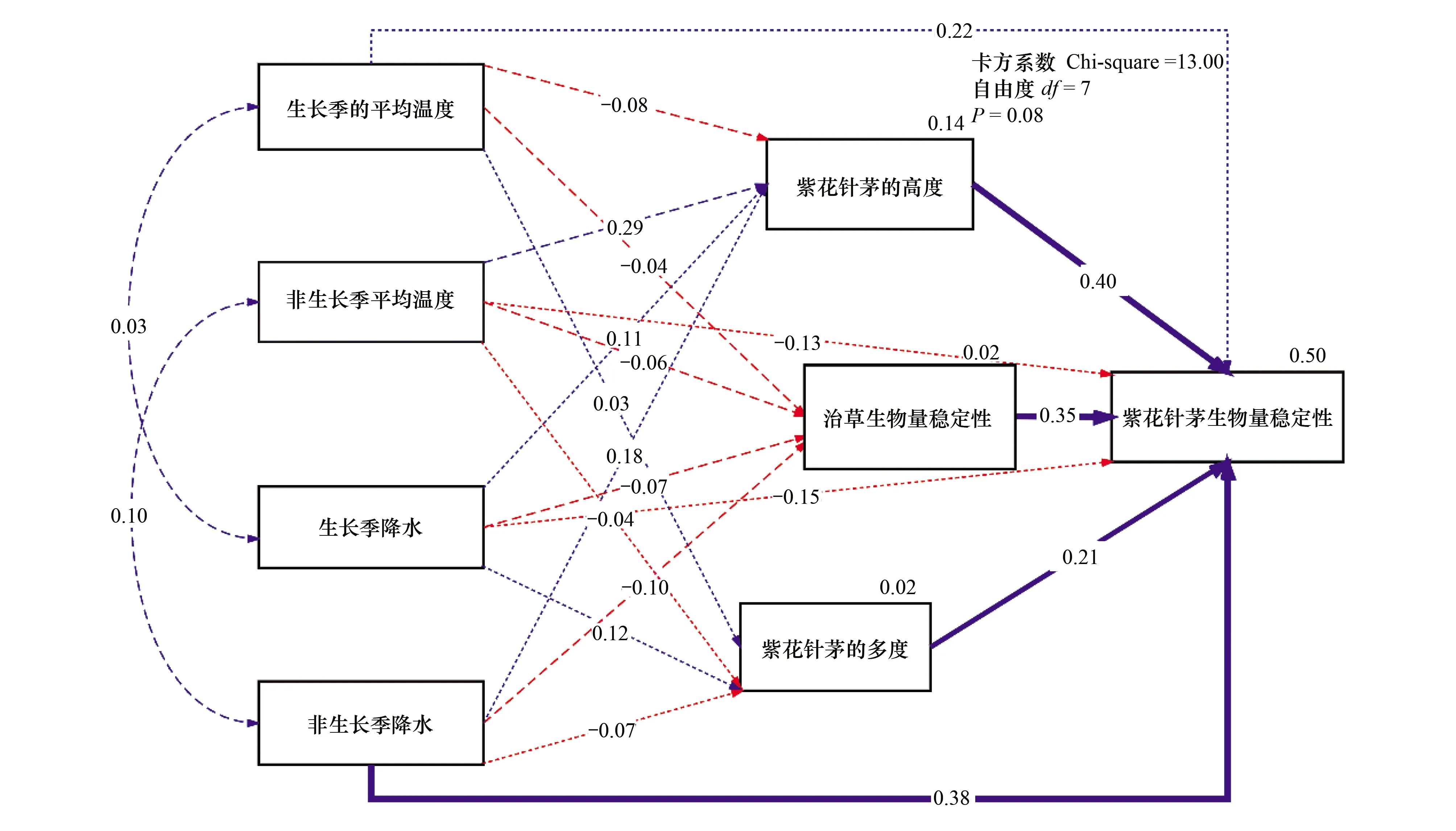
圖4 紫花針茅生物量穩定性的環境因子和生物因子的結構方程模型Fig.4 Structural equation model of environmental factors and biological factors for biomass stability of Stipa purpurea響應量(紫花針茅生物量穩定性、洽草生物量穩定性、紫花針茅高度、紫花針茅多度)右上角的百分比表示模型(R2)能解釋的方差;虛線表示不顯著的影響;單向箭頭上的數字為解釋度系數,雙向箭頭上的數字為相關系數。每個結構內變量與響應量連線上的百分比是被解釋的變量,紅色代表負效應,藍色代表正效應
3 討論
隨著全球氣候變化的加劇,青藏高原的氣候變化更加明顯,高山草甸生態系統也受到了較為嚴重的影響[5,51,54]。本研究通過長期觀測資料,分析生長季平均氣溫、非生長季平均氣溫、生長季降水量、非生長季降水量的數據,旨在厘清生長季/非生長季的氣溫和降水的不同步變化對于生態系統穩定性的影響。
近二十年的觀測數據與其它研究結果一致,表明受全球氣候變化影響,青藏高原上整體氣候呈現暖濕化趨勢[55],本研究發現,生長季的溫度和降水與全年的溫度降水均呈現顯著的正相關,這也與青藏高原往往雨熱同季的氣候特征相吻合,即在生長季呈現暖濕化的趨勢。但是,本研究并沒有發現非生長季溫度和降水呈現顯著的趨勢,這表明青藏高原過去十幾年的季節不對稱氣候變化呈現增加的趨勢,這與相關研究的結論一致[35—38]。
有研究表明,氣候變化對植物的影響表現為:在高緯度地區,低溫對植物生長的限制作用大于低緯度和高海拔地區[55],因此高寒植物的生物量穩定性隨著氣候變化的變化(特別是溫度的增加)可能會發生比較大的變化[56]。但本研究發現,優勢物種的生物量穩定性及其生態特征(高度和多度)在過去十幾年期間,并沒有發生顯著的變化,與優勢物種不同,本研究發現次優勢物種的生物量穩定性在過去十幾年期間呈現顯著下降的趨勢,這可能與優勢物種之間的相互作用有關系。
有研究表明,優勢物種在溫度升高時優勢度會升高[57—59],從而增加同時期優勢種與次優勢物種競爭作用的強度[60]。探究優勢物種和次優勢物種之間相互作用的方向和強度對氣候變化的響應是理解氣候變化對植物群落影響的基礎。優勢物種決定了群落中種間相互作用的強度并能對氣候變化做出及時的響應[24],優勢種群對生態系統結構和功能具有潛在的影響[41—42]。但是本研究發現,在環境因子里面只有非生長季降水與紫花針茅生物量穩定性呈現顯著的正相關,此外紫花針茅自身的生態特征,如高度也與其生物量穩定呈現顯著的正相關,以上表明紫花針茅作為優勢種對氣候變化更敏感且受到的影響是正面的[34],此外優勢物種的生物量穩定性也會受到自身特征以及次優勢物種的影響。
在高寒地區,溫度是平衡競爭和促進關系的決定因素[61]。生長季平均氣溫的升高直接緩解高寒地區低溫脅迫[62]。非生長季平均氣溫升高減少了霜凍災害的發生頻率,提高了植物的抗旱性[63],同時還提高了幼苗的存活率。其次,部分永凍層和積雪隨著冬季溫度升溫而融化,增加了土壤在冬季的含水量,保護了植物地下掩埋部分,有利于植物在其后春夏兩季的生長發育[63—64]。但是,本研究并沒有發現生長季或者非生長季溫度并沒有對優勢物種或者次優勢物種穩定性產生影響,這與Fu等人[41]的研究結果并不一致,這可能與時間的年限有關系,即短期的控制性實驗與長期氣候變化記錄是存在一定的差異的,本研究猜測優勢物種與次優勢物種的相互作用緩沖了季節不對稱增溫導致的效應,但是相關的結果還需要進一步的實驗驗證。此外,本研究并沒有考慮次優勢物種的高度和多度對于次優勢物種和優勢物種生物量穩定性的影響,這主要是其數據的缺失導致無法滿足統計學要求,這會造成一定的誤差,但是本文的研究結果可以反映生長季和非生長季不同步氣候變化背景下,優勢物種本身的特征(包括高度和多度)以及次優勢物種生物量穩定性對于優勢種的生物量穩定性的影響。
降水是高寒群落生產的主要驅動力,影響群落生物量穩定性[62,65],尤其高寒地區,降水往往成為植物生長的限制因子[66]。降水量的季節變化會影響生物量的穩定性[67],分析得出非生長季降水量與紫花針茅生物量穩定性的正相關性,大于生長季平均氣溫、非生長季平均氣溫,甚至大于生長季降水量。然而紫花針茅多度對降水增加的響應與年降水量無關,這與Wu等人的研究一致[68],只與生長季/非生長季降水量有關。本研究猜測,降水量的增加會在某種程度上解除溫度升高導致的水分限制,特別是在植物開始返青的時候,最終生物量的穩定性就會隨著暖濕化的氣候變化而增加[69,70—71]。于此同時,本研究也從優勢物種和次優勢物種穩定性與氣候變化的關系中驗證了本研究的推論,即優勢物種決定了群落中種間相互作用的強度并能對氣候變化做出及時的響應[38,61]。
4 結論
青藏高原上高寒草甸對于氣候變化較為敏感,但形成了對持續的暖濕化進行適應的策略,以保持自身生物量穩定性;優勢種和次優勢種對生長季/非生長季不對稱升溫的響應并不顯著;前人的研究很少關注非生長季降水量對于生態系統穩定性的影響,本研究發現非生長季的降水會促進次優勢物種的生長,最終促進高寒草甸中優勢物種的生物量穩定性,預計在青藏高原未來的暖濕化的氣候變化下,非生長季的降水結合物種間尤其是優勢物種之間相互作用,并最終決定高寒草甸的穩定性。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
小讀者(2021年2期)2021-03-29 05:03:48
趣味(語文)(2020年3期)2020-07-27 01:42:46
華人時刊(2019年13期)2019-11-17 14:59:54
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
文苑(2018年22期)2018-11-19 02:54:14
汽車觀察(2018年10期)2018-11-06 07:05:26
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
