流速對光棘球海膽行為和生長的影響
2022-06-28 03:11:58施棟濤尹東紅常亞青
生態學報 2022年10期
施棟濤,趙 沖,尹東紅,陳 陽,常亞青
農業農村部北方海水增養殖重點實驗室,大連海洋大學,大連 116023
流速作為重要的環境物理因子,對海洋生物的生存、生長及種群分布等均有重要的影響[1—4]。海膽是一類常見的棘皮動物,多生活在水流變化較大的淺海和潮間帶區域,具有重要的生態和經濟價值[5]。研究表明流速對海膽的影響在個體和種群層面都有表現。在個體層面,海膽的移動[6—9]、覓食[10—11]及生殖[12—13]等行為在不同流速下均會表現出顯著的差異。在種群層面,水流速度影響海膽的棲息區域,不同種海膽對棲息地的選擇不同[14—16]。
正形海膽幼膽和成膽以大型藻類為食(如海帶和石莼)。該類海膽的覓食過程主要涉及管足活動[17],移動[18—19],口器咬合[20—21]等行為。海膽利用管足進行移動,管足的粘附能力可以幫助海膽粘附在大型海藻上,海膽進而利用活動口器進行咬合來完成覓食活動[17,22]。Tamaki等[23]的野外研究表明流速限制了海膽的覓食區域[24]。這可能是由于高流速的增加嚴重影響了海膽的運動和攝食[19]。實驗室研究也證明了這一點,在24 h的單向水流作用下,海膽的攝食率隨著流速的增加而下降[11]。翻正行為是海膽在口面向上的情況下,利用管足使其口面向下的行為模式,是測試海膽活力的有效性狀[25—26]。本研究之前的短期實驗結果表明,海膽的翻正行為受到高流速抑制[9]。海膽的體尺也受到不同區域流速的影響。Cohen Rengifo等[27]在不同流速海區采集的同種海膽在體尺大小,管足的數目和韌性均出現了顯著差異。目前關于流速對海膽影響的報道主要集中在實驗室短期(0—2 d)研究[9,11,19]和野外觀察上[19,23,26]。短期研究難以闡明流速的長期影響,而野外海區環境復雜,海膽易受到流速之外的多種非實驗因素的干擾。因此,開展相對長時間的實驗室研究對揭示流速對海膽行為和生長的影響具有重要意義。
光棘球海膽(Mesocentrotusnudus)作為我國北方沿海潮間帶地區的常見物種,具有重要的生態和經濟價值。國家海洋信息中心流速數據表明,大連海洋牧場相關海域大部分時間流速穩定(<20 cm/s)且有良好的水流交換,適合海洋經濟生物的底播增殖[28]。Kawamata[19]和Tamaki[23]的短期實驗表明20 cm/s流速下,海膽的攝食率顯著下降,超過20 cm/s流速區域海膽數目顯著減少。依據Shi等[9]之前的結果,流速穩定時進行底播可減小流速對海膽增殖放流的影響,但其在長期水流影響下的生長和攝食能力仍缺乏研究。因此,本文根據現有研究和海區流速環境確定10 cm/s和20 cm/s為本研究的中和高流速,2 cm/s為自然海區較低的流速。本實驗主要研究了光棘球海膽1)在不同流速下的生存和生長情況;2)在不同流速下的覓食和攝食行為;3)在不同流速下管足附著和翻正行為。本研究有助于進一步理解流速對海膽的長期影響,為進一步闡明海膽底播增殖的適宜流速提供理論參考。
1 材料與方法
1.1 海膽
本實驗用光棘球海膽全部來源于大連市金州某育苗場(殼徑 =(21.57±0.14)mm,殼高=(10.12±0.10)mm,體重 =(4.19±0.08)g)。實驗前海膽暫養于農業農村部北方海水增養殖重點實驗室。實驗使用海膽均健康,無疾病癥狀。暫養池為玻璃鋼水槽(長×寬×高=180 cm×100 cm×70 cm),期間飼以新鮮海帶(足量投喂,每2 d約10 g/只)。此外,本研究每兩日換水1/3,清理糞便。實驗室光照為自然光,暫養期間連續充氣確保海膽不會缺氧。
1.2 實驗設計
為了研究流速對海膽的長期影響,實驗設置在低(2 cm/s),中(10 cm/s)和高(20 cm/s)3種流速下進行。

圖1 長期流速實驗裝置示意圖 Fig.1 Diagram of the equipment for the long-term flow velocity experiment 水流為單向,由入水口進入后,經整流板進入實驗區后自出口流出。實驗區有5個小室,每一個小室為一個實驗區(長×寬×高=20 cm×15 cm×10 cm),其間用網相隔,底部放置有底質模擬板
本實驗使用環形跑道(圖1)進行,跑道由網分割出5個體積相等小室(長×寬×高=20 cm×15 cm×10 cm)。每一小室底部放置一塊底質模擬板。底質板由沙子(采集于黑石礁海邊,由1 mm孔徑篩網篩出,約50 g)和石頭(采集于黑石礁海邊,約40 g,約15個)作為底質,使用環氧樹脂(20 g)經行粘結。水流為單向流動(如圖1),由入口進入后,經過各小室流至出水口。出水口掛置一個過濾網袋過濾海膽糞便。食物由綁在底質板上的塑料夾子夾住。本研究對實驗過程中其它發現也進行記錄。海膽體尺性狀使用游標卡尺(Mahr,德國)測量,重量使用電子天平(G & G,USA)測量。實驗開始前利用流速儀(JDC,Swiss)調整流速至目標流速,并保證每個小室的流速差距在±0.5 cm/s。為保證試驗期間水質,保證充氣充足并每日換水1/3,日常測量其溫度,pH和鹽度。實驗進行時,每一小室放置5只海膽,每一流速和每一小室間的海膽在體尺測量上均無統計學差異。
1.3 攝食
為了評估長期水流對海膽攝食的影響,本研究測量了海膽每日的攝食量。每個小室每次投食均使用相同部位并切成大小約11 cm×5 cm的新鮮海帶(約20 g)。每日定時投喂海帶,并稱量前一日投喂海帶的剩余重量,以確定其每日的攝食情況,測量共39次(n=5)。在測量攝食期間,對每一流速下的海膽的糞便留存量進行了3次測量。
1.4 生長
實驗進行中,為了研究海膽在不同流速下的生長的情況,每周對其進行生長相關性狀的測量。本實驗中,測量性狀包括海膽的殼徑(mm),殼高(mm)和體重(g)。每個小室中的5個海膽平均值作為一個獨立樣本數值進行分析(n=5)。最后,本研究中計算了海膽的增重率(WGR),特定生長率(SGR)和飼料轉化率(FCR),公式如下:
式中,w0表示海膽初始體重,w表示養殖后體重,t1表示開始養殖的時間,t2表示終末時間,F表示總攝食量。
1.5 翻正行為
在長期流速影響的養殖實驗結束后,立即對海膽的翻正行為進行測試。這一實驗分別在原位(攝食實驗時所暴露的流速)和靜水中進行。將海膽口面朝上放置于裝置底部,記錄海膽利用其管足和棘調整至反口面朝上的正常體態所需時間,若海膽超過300 s仍未恢復到口面向下的姿態,則翻正時間記為300 s[9]。
1.6 覓食時間和附著時間
在長期流速影響的養殖試驗后,本研究測量了海膽在不同流速下的覓食能力。在原流速下進行了7 d禁食處理后,將新鮮海帶置于上游區域,并將海膽放置于下游區域中間(兩個小室大小,長×寬=40 cm×15 cm),觀察海膽的移動和覓食情況。本實驗全程使用攝像機(SONY,FDX- 55)進行攝像,實驗后進行計算和分析。在共5個小室中,每個小室內隨機挑選2只海膽進行實驗(n=10)。覓食行為時長持續為480 s,若超過這一時間海膽仍未到達食物點,則將覓食時間記為480 s[29]。
海膽覓食實驗前,進行海膽管足附著能力的測量。本研究測量了海膽附著的時間以確定水流對管足的附著能力的影響。將3種不同流速下養殖的海膽放置于與其暴露流速相同的流速下,記錄其用管足將自身附著于底面所需的時長。實驗開始時將海膽放置于底面并開始計時,當其管足附著于底面不被水流移位且可移動時記為附著完成,期間為附著時間。
1.7 口器咬合行為
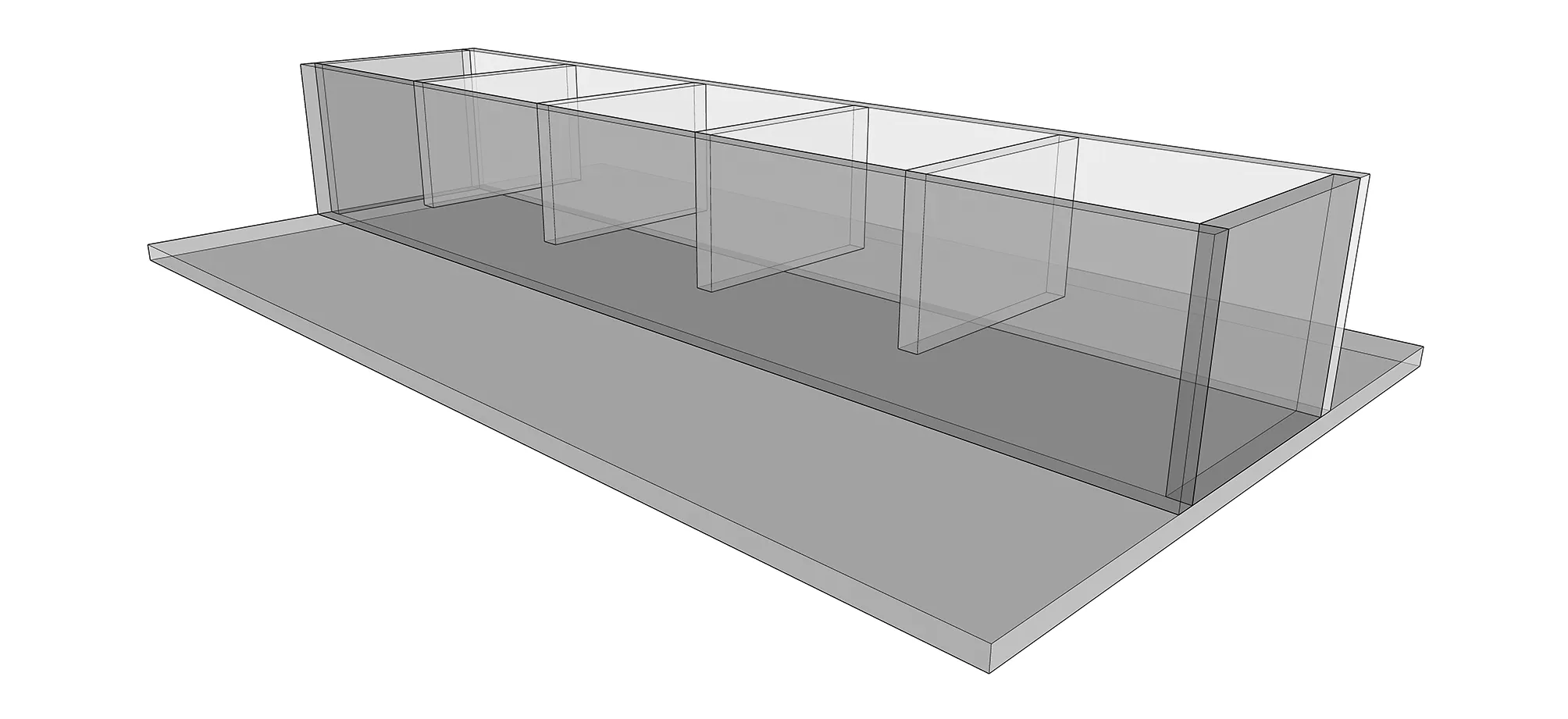
圖2 口器咬合行為實驗裝置示意圖 Fig.2 Experimental facility for Aristotle′s lantern reflex of Mesocentrotus nudus
為了研究長期暴露在不同流速下的海膽的攝食能力,在覓食行為后5 d(期間正常投喂)進行了口器咬合行為的實驗。實驗使用定制的亞克力器具如圖2所示。實驗開始前,根據Shi等[30]的方法配置海藻膠并鋪于容器內,待其凝固后加滿相同溫度海水。實驗時,將海膽放入容器中,并在上方蓋一聚乙烯塑料泡沫固定。使用攝影機(SONY,FDX- 55)在容器下方進行拍攝。在共5個小室中,每個小室內隨機挑選2只海膽進行實驗(n=10)。實驗后通過錄像計數海膽在300 s內的口器咬合次數。
1.8 數據分析
本研究數據均使用SPSS(vision 22)進行分析。分析前分別用Kolmogorov-Smirnov和Levene′s的方法檢查數據的正態分布情況和方差齊性。海膽的初始殼徑、殼高和體重使用單因素方差分析。養殖期間海膽攝食量、生長性狀和之后測試的翻正行為使用單因素方差分析,如有顯著差異則進一步使用LSD方法進行兩兩比較。對不符合正態分布或方差齊性的覓食行為和管足附著時間數據進行非參數檢驗分析(Kruskal-Wallis方法)。顯著性概率水平為P< 0.05。
2 實驗結果
2.1 攝食量
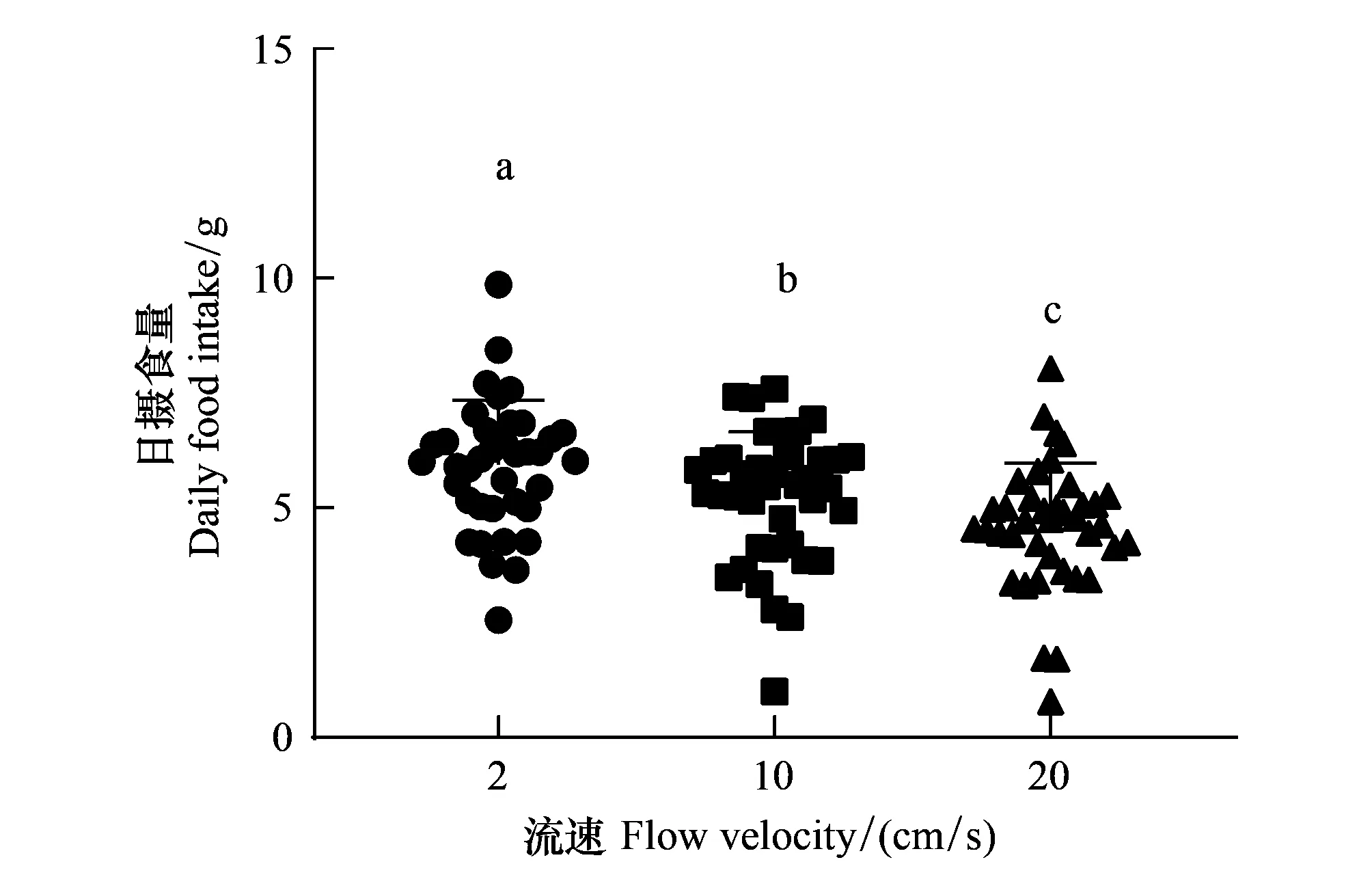
圖3 光棘球海膽在2 cm/s,10 cm/s和20 cm/s下的每日平均攝食量(平均值±標準差)Fig.3 Daily feeding of Mesocentrotus nudus at 2 cm/s, 10 cm/s and 20 cm/s (mean±SD)不同小寫字母表示在不同流速下日平均攝食量存在顯著性差異

圖4 光棘球海膽于2 cm/s時,殼面部分留有糞便Fig.4 Left feces on the top of the urchin test at 2 cm/s
養殖試驗中測量海膽每日攝食量39次,期間無海膽死亡。結果表明在流速為2 cm/s,10 cm/s和20 cm/s時的每日平均攝食量分別為(5.94±1.87)g,(5.22±1.90)g和(4.59±1.81)g。其中流速為2 cm/s時的每日攝食量顯著大于10 cm/s和20 cm/s(P< 0.001和P< 0.001),而10 cm/s時的攝食量顯著大于20 cm/s(P=0.001)(圖3)。
此外,在實驗中發現一定的水流對海膽的糞便有清理作用。當流速為2 cm/s時底板和海膽反口面部會殘留部分糞便((4.66±1.85)g,圖4),而在共3次的測量中流速為10 cm/s和20 cm/s時則沒有糞便殘留(n=3)。
2.2 生長
在對海膽殼徑和體重的7周連續測量中,自第2周和第3周,海膽的體重和殼徑就開始在不同流速組間出現顯著差異(表1)。
在49 d中,光棘球海膽在2 cm/s時的增重率(圖5)顯著大于10 cm/s和20 cm/s組(P=0.015和P=0.001),而后兩者間無顯著差異(P=0.099)。特定增長率(圖5)也表明流速顯著地影響了光棘球海膽的生長。流速為2 cm/s時海膽的特定增長率顯著大于10 cm/s和20 cm/s(P=0.007和P< 0.001),而后兩者無顯著差異(P=0.060)。飼料轉化率(圖5)在流速為20 cm/s時要顯著大于2 cm/s(P=0.045),而2 cm/s和10 cm/s,10 cm/s和20 cm/s間無顯著差異(P=0.057和P=0.899)。
2.3 附著時間
流速顯著影響海膽的管足的粘附時間(圖6)。在10 cm/s和20 cm/s下,海膽的管足粘附時間顯著長于在2 cm/s下(P< 0.001和P< 0.001)。在10 cm/s和20 cm/s組之間無顯著差異(P=0.051)。
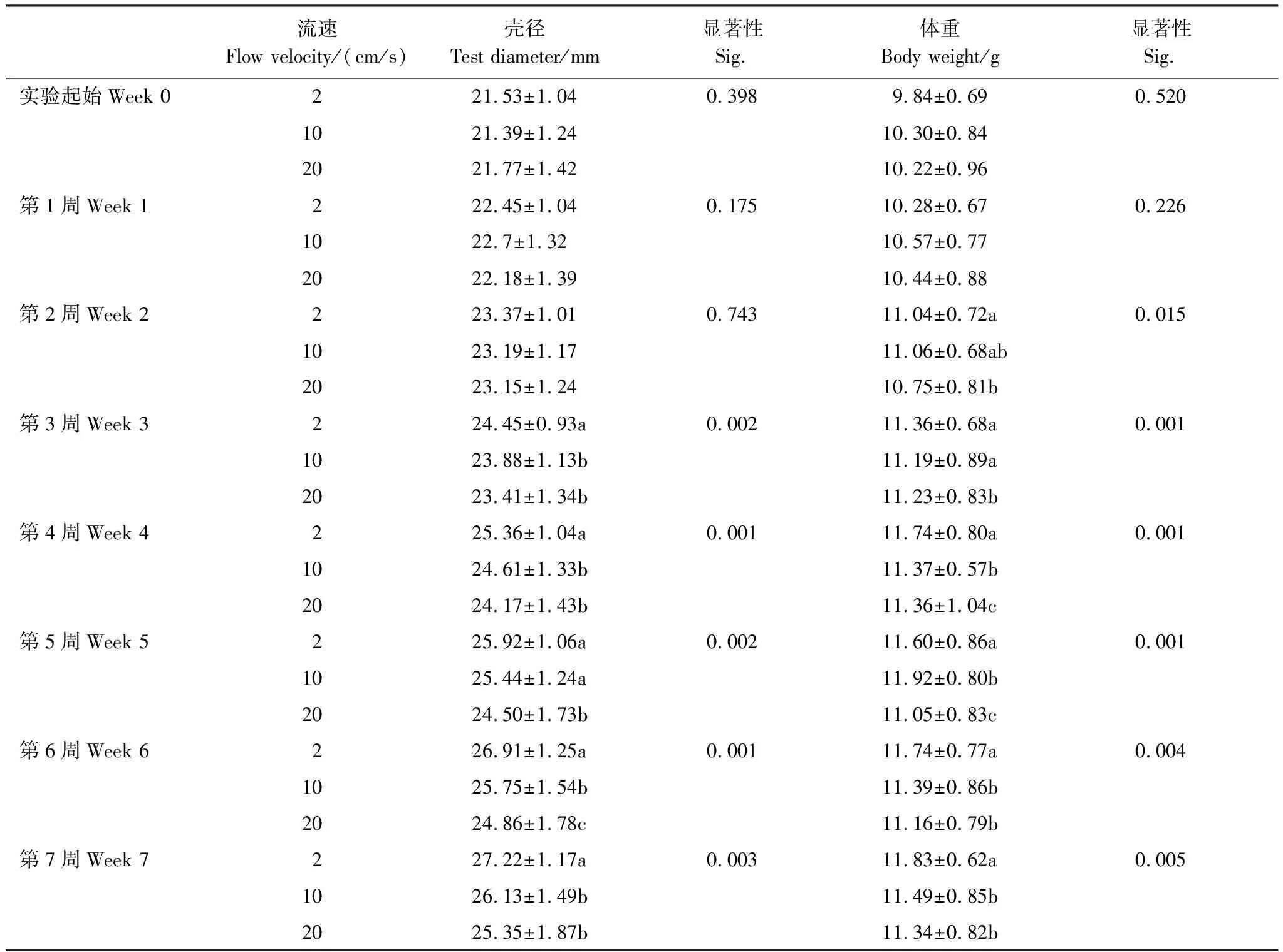
表1 光棘球海膽在2 cm/s,10 cm/s和20 cm/s流速下7周內的殼徑與體重(平均值±標準差)
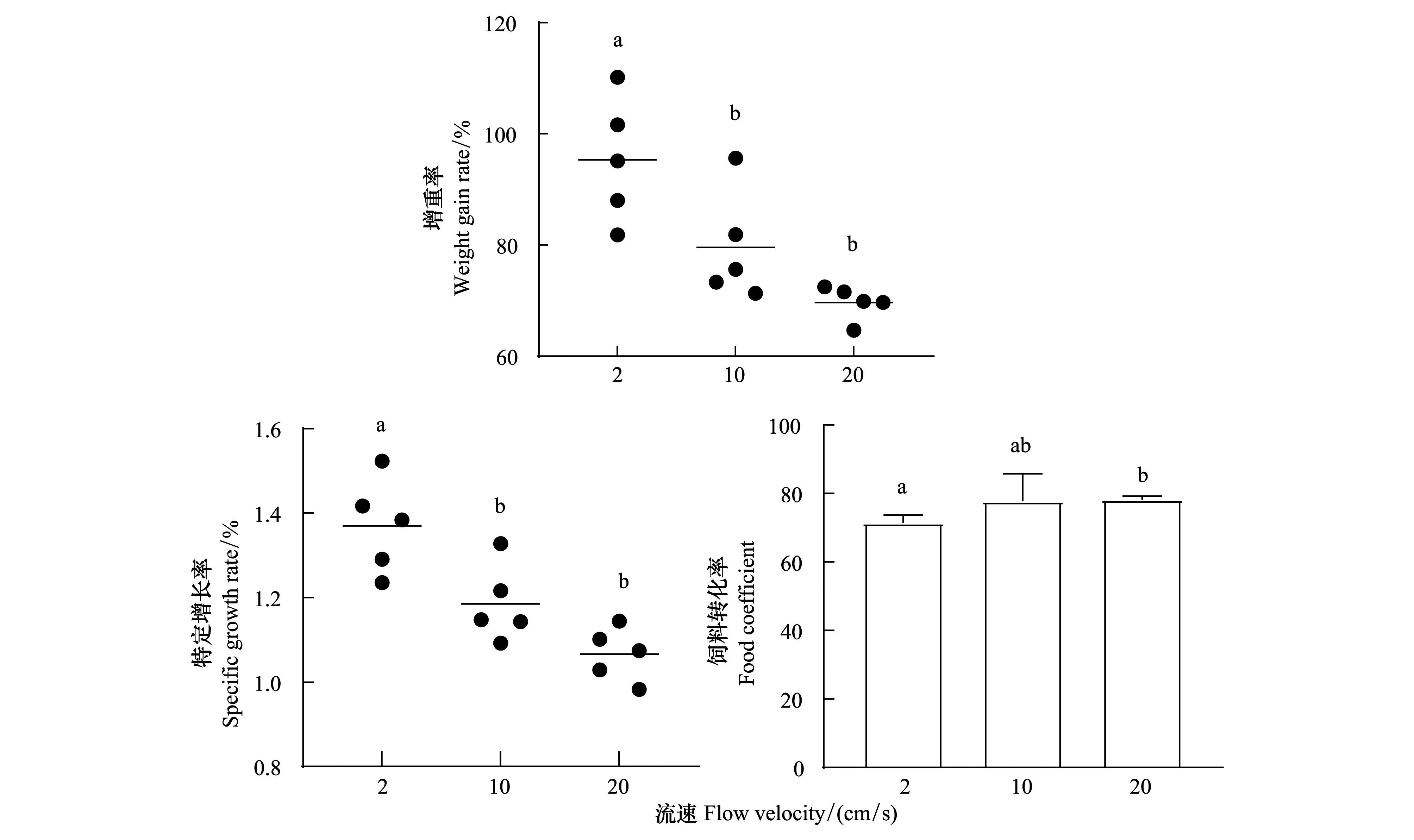
圖5 在2 cm/s,10 cm/s和20 cm/s流速下光棘球海膽的增重率、特定生長率、飼料轉化率變化Fig.5 Weight gain rate, specific growth rate and food coefficient of Mesocentrotus nudus at flow velocities of 2 cm/s, 10 cm/s and 20 cm/s不同小寫字母表示在各實驗組中存在顯著性差異
2.4 覓食時間
光棘球海膽在不同流速下的覓食時間存在顯著差異。在20 cm/s流速下,海膽的覓食時間顯著多于在2 cm/s時(P=0.004),但是在10 cm/s和20 cm/s組之間無顯著差異(P=0.200)(圖6)。
2.5 口器咬合
海膽口器咬合在2 cm/s,10 cm/s和20 cm/s流速下無顯著差異,但是在20 cm/s的流速下咬合次數相對更多(P=0.113)(圖6)。
2.6 翻正行為
在原位翻正行為測試中,相對于2 cm/s(P=0.02)和10 cm/s(P=0.03),當流速為20 cm/s時海膽的翻正時間顯著增加。海膽原位翻正時間在2 cm/s和10 cm/s之間沒有顯著差異(P=0.840,圖6)。
在靜水中測試的海膽翻正時間結果表明,在2 cm/s,10 cm/s和20 cm/s下長時間暴露的各組海膽翻正時間之間無顯著差異(P=0.208)。
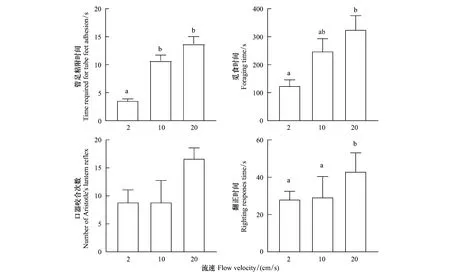
圖6 光棘球海膽在2 cm/s,10 cm/s和20 cm/s流速下長期暴露后的行為Fig.6 The behaviors of Mesocentrotus nudus at flow velocities of 2 cm/s, 10 cm/s and 20 cm/s不同小寫字母表示在不同實驗組中存在顯著性差異
3 討論
棲息于潮間帶和淺海,海膽的分布、行為和生長更容易受到水流環境變化的影響[11,26,31]。因此,研究海膽在長期不同流速影響下的生存、生長和行為,具有重要的生態和經濟價值。本實驗研究了長時間(49 d)不同流速(2 cm/s,10 cm/s和20 cm/s)對光棘球海膽的生存、生長和行為的影響,為進一步闡明海膽底播增殖的適宜流速提供理論參考。
野外調查顯示流速影響海膽的生長[27]。本研究表明海膽的殼徑和體重的增長在高流速下均受到了顯著抑制,這一現象在第2、3周已表現出來。海膽的增重率為2 cm/s組>10 cm/s組和2 cm/s組>20 cm/s組。這表明高流速抑制了海膽體重的增長。特定生長率與增重率結果同樣為2 cm/s組>10 cm/s組和2 cm/s組>20 cm/s組。這進一步表明高流速對海膽生長的抑制作用。該結果與野外調查有一定相似性,在流速較高的區域收集的海膽在體尺上小于流速較低區域[27]。本文推測海膽增重受到抑制可能是攝食量受到影響的結果。進一步研究發現,海膽攝食量在不同流速下為2 cm/s組>10 cm/s組>20 cm/s組。這說明在不同流速下的攝食量對海膽生長可能具有較大影響。海膽的覓食或者攝食(口器咬合)能力下降可能進一步導致了攝食量的下降。然而,海膽口器咬合次數并未在3種流速下產生顯著差異,這說明海膽的口器攝食功能穩定性好,與Shi等[30]的研究結果相似。Tamaki等[23]的野外調查結果顯示光棘球海膽幾乎不會出現在較高流速(14.5±3.4)cm/s但食物更為豐富的區域。這與本實驗結果相一致,光棘球海膽在低流速下的覓食所需時間隨著流速的升高而增加(10 cm/s組>2 cm/s組,20 cm/s組>2 cm/s組)。這表明較高流速削弱了海膽覓食能力。因此,高流速下每日攝食量的減少可能是由于流速對光棘球海膽覓食能力的影響,這也與Tamaki等[11]的短期實驗結果相一致。較低流速下海膽持續增長以較高的生長率為特征,而體型的增長可能會幫助其建立抵抗水流的優勢[11,19]。因此,中低流速區域(2—10 cm/s)更適合海膽生存、攝食和生長。低流速海域的大型藻類可能會因海膽的大量攝食而減少,進而影響低流速區域的生態環境[24]。
海膽的覓食行為需要依靠管足的粘附—脫離—粘附過程來完成。這也是其附著、移動等行為的基礎[32]。在高流速下海膽生長更緩慢,可能是因為管足消耗額外的能量用來抵御水流影響,而不能用于生長[33—34]。本研究發現飼料轉化率在20 cm/s下顯著大于低流速(2 cm/s),這表明光棘球海膽的增重所消耗的食物(海帶)隨著流速的增加而增加。管足附著時間實驗表明管足吸附能力受水流顯著影響,長時間流速暴露后海膽的覓食能力仍會受到水流的顯著影響。海膽管足附著所需時間在3種流速下存在著顯著差異,中高流速暴露下海膽要消耗更長時間才能完成附著,這可能是能量消耗的主要原因之一。而飼料轉化率在不同流速下的海膽中無顯著差異。這說明當流速升高到一定程度后,海膽需要用于保持附著所需的消耗可能會趨于平衡[35]。此外,較高流速(10 cm/s和20 cm/s)下海膽的糞便幾乎不會在原地殘留。本研究表明,在保證食物供應的前提下,流速為2—10 cm/s的區域有利于海膽生長。
翻正行為幫助海膽在不利環境(如海流)造成姿態失常后恢復正常。這一行為主要由管足活動影響,是判斷海膽環境適應性的重要指標[26]。在原位實驗中,海膽的翻正所需時間隨著流速的增加而增加。這表明海膽的翻正能力會被削弱,這也是管足吸附能力減弱的具體體現[36]。與之前的短期研究[9]結果不同,長期的流速適應下的海膽均成功完成翻正并耗時更短。這說明海膽在長期高流速下,或已經出現了一定適應性的表現,但也無法排除體型差距可能造成的影響。Cohen Rengifo等[27]研究表明,在不同流速區域生長的海膽在管足密度表現出了差別。這說明海膽對流速的適應很有可能會通過管足表現(例如,翻正行為)來實現[26,33]。而海膽在靜水中的翻正所需時間無顯著差異,這表明這一行為穩定性好,在10 cm/s和20 cm/s長期的流速下未受到顯著影響。
綜上所述,不同流速暴露顯著影響光棘球海膽(殼徑:約20 mm)殼徑和體重的增長,但對其生存無顯著影響。海膽的生長隨著流速的升高而顯著降低。高流速(20 cm/s)顯著削弱海膽管足的粘附能力,進而顯著降低其翻正,移動和覓食行為,但并不顯著影響其口器咬合行為。飼料轉化率結果表明高流速(20 cm/s)下海膽需要消耗更多飼料以維系其生長。這表明流速對海膽生長的影響可能不僅源于攝食量和覓食行為的差異,也源于其生存的基本消耗差異。根據我們結果和國家海洋信息中心關于適于海膽底播增殖海域的流速數據,本研究建議有關生產單位在綜合考慮各種環境因子和生產要素的前提下,在流速常年在2—10 cm/s的海區開展光棘球海膽幼膽底播增殖生產,或對相關海域進行海洋牧場建設,投放躲避物減少流速影響。
致謝:感謝國家科技資源共享服務平臺——國家海洋科學數據中心(http://mds.nmdis.org.cn/)提供數據信息。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
