荒漠植物珍珠豬毛菜根際土壤細菌多樣性與土壤理化性質相關性分析
2022-07-13 09:04:02魏艷晨陳吉祥王永剛孟彤彤韓亞龍李美
中國農業(yè)科技導報 2022年5期
魏艷晨, 陳吉祥*, 王永剛, 孟彤彤, 韓亞龍, 李美
(1.蘭州理工大學石油化工學院,蘭州 730050;2.蘭州理工大學生命科學與工程學院,蘭州 730050;3.蘭州理工大學能源與動力工程學院,蘭州 730050)
西北荒漠區(qū)作為我國典型的中-重型生態(tài)脆弱區(qū),是氣候變化響應的敏感帶,也是西北生態(tài)安全的重要屏障。景泰位于甘肅中部,河西走廊東端,地處甘肅、內蒙、寧夏交界處,屬于黃土高原與騰格里沙漠過渡地帶。氣溫多變、干旱多風少雨、沙化嚴重,是典型的干旱生態(tài)脆弱區(qū)[1]。珍珠豬毛菜(Salsola passerina)屬藜科半灌木,廣泛分布于景泰縣干旱荒漠區(qū)礫石質、沙礫質戈壁和鹽堿湖盆地,具有抗風沙、耐旱和耐寒的特性,是維護區(qū)域生態(tài)平衡的重要植物。研究荒漠區(qū)土壤細菌多樣性對荒漠植物生態(tài)適應及生態(tài)系統(tǒng)修復有重要意義。不同荒漠區(qū)優(yōu)勢植物類群根際土壤的微生物類群及豐度有明顯差異。康寶天等[2]研究祁連山高寒草甸和荒漠草原土壤細菌群落組成發(fā)現(xiàn),不同草地土壤的細菌群落存在顯著差異,放線菌門和綠彎菌門在高寒草甸土壤中的相對豐度顯著低于荒漠草原,而變形菌門、酸桿菌門、擬桿菌門和Rokubacteria則相反。黃耀龍[3]研究發(fā)現(xiàn),荒漠植物梭梭(Haloxylon ammodendron)與白刺(Nitraria tangutorum)根際土壤的細菌多樣性差異顯著。單立山等[4]和李善家等[5]對聯(lián)生珍珠豬毛菜和聯(lián)生紅砂、單生珍珠豬毛菜和單生紅砂的比根長、比表面積、根系生物量的垂直分布特征及葉片功能性狀進行比較,驗證了紅砂-珍珠豬毛菜聯(lián)生群落適應干旱脅迫環(huán)境。然而,在荒漠環(huán)境脅迫下,單生珍珠豬毛菜根際土壤和荒漠土壤的細菌在數(shù)量和多樣性均可能具有差異,且二者土壤理化性狀也可能存在差異。為此,本研究選取珍珠豬毛菜根際土壤和荒漠土壤(周邊非根際土壤)為研究對象,采用高通量測序對珍珠豬毛菜根際和周邊非根際土壤的細菌群落結構進行研究;通過對比根際和非根際土壤的可培養(yǎng)細菌數(shù)量、細菌總數(shù)、理化性質、微生物群落結構的變化和區(qū)別,分析土壤細菌群落組成和土壤理化性質的關系,為珍珠豬毛菜生態(tài)適應及荒漠草原生態(tài)修復提供理論基礎。
1 材料與方法
1.1 研究區(qū)概況及樣品采集
研究區(qū)地處甘肅省景泰縣境內荒漠草原區(qū)(E 103°55′,N 36°55′),屬溫帶大陸性干旱氣候,海拔1 276~3 321 m,年均氣溫8.2℃,年均降水量185 mm,年蒸發(fā)量達3 038.5 mm,無霜期191 d。該區(qū)主要地貌為風沙地貌,土壤以荒漠土和灰鈣土為主。植被結構簡單且稀疏,主要有珍珠豬毛菜、紅砂,其中珍珠豬毛菜分布較為廣泛。
在研究區(qū)內取珍珠豬毛菜根際土壤(rizosphere soil,GJ)和非根際土壤(non-rizosphere soil,NG)。其中,GJ:選取長勢一致的珍珠豬毛菜3株做重復,隨機選取離植株莖稈2~5 cm處的3個采樣點,用土鉆法取樣,垂直深度0—60 cm,將所取土樣混勻后裝入無菌保鮮袋于低溫保存待測;NG:自同一采樣點周邊區(qū)域,去除地表0—5 cm處土壤,收集5—60 cm土壤裝于無菌保鮮袋,于冷藏條件帶回實驗室進行分析。
1.2 土壤理化性質的測定
土壤pH采用pH計測定;土壤含水率(water content,WC)用烘干法測定;土壤全氮(total nitrogen,TN)用凱氏定氮法測定;土壤速效氮(available nitrogen,AN)采用堿解擴散法測定;土壤全磷(total phosphorus,TP)采用堿熔融-鉬銻抗比色法測定;土壤速效磷(available phosphorus,AP)采用NaHCO3浸提-鉬銻抗比色法測定;土壤全鉀(total potassium,TK)采用氫氟酸消解-原子吸收分光光度計法測定;土壤速效鉀(available potassium,AK)采用NH4Ac浸提-火焰光度計法測定[6]。
1.3 土壤細菌總數(shù)和可培養(yǎng)細菌數(shù)的測定
土壤細菌總數(shù)測定采用“LIVE/DEAD BacLight Bacterial Viability Kit”試劑盒,根據(jù)說明書進行。稱取10 g土樣于90 mL滅菌生理鹽水30℃振蕩30 min,使土壤中細菌均勻分散,制成10倍稀釋度的土壤懸液,離心收集菌體,用生理鹽水重懸。SYTO9和碘化丙啶核酸染色劑染色后,用激光共聚焦顯微鏡觀察[7]。可培養(yǎng)細菌數(shù)采用稀釋涂布平板法[8]測定。土壤懸液依次進行 10-1、10-2、10-3、10-4稀釋。取不同稀釋度土壤懸液100 μL,涂布LB平板,每個稀釋度設置3個平行。
1.4 土壤細菌多樣性分析
采用HiPure Soil DNA Kits試劑盒(D3142,廣州美基生物科技有限公司)提取土壤樣本的DNA進行PCR擴增,擴增區(qū)域為細菌16S rDNA的V3-V4區(qū),引物為341F(5′-CCTACGGGNGGCWGCAG-3′)和806R(5′-GGACTACHVGGGTATCTAAT-3′)。使用AMPure XP Beads對擴增產物進行純化,用ABI StepOnePlus Real Time PCR System(Life Technologies,產地美國)進行定量,最后根據(jù)Novaseq 6000的PE250模式pooling上機測序[9]。
1.5 數(shù)據(jù)處理
采用Uparse軟件對所有樣品的全部有效Tags序列聚類,以97%相似水平的OTUs序列進行分類分析。利用SPSS 22.0軟件進行統(tǒng)計分析。采用Image J軟件完成CLSM數(shù)據(jù)統(tǒng)計分析。
2 結果與分析
2.1 土壤理化性質分析
珍珠豬毛菜根際土壤和非根際土壤的理化性狀存在顯著差異(表1)。其中,珍珠豬毛菜根際土壤的全氮、全鉀、速效氮、速效磷和速效鉀均顯著高于非根際土壤;土壤含水量和pH顯著低于非根際土壤;土壤全磷含量在根際土壤和非根際土壤間差異不顯著。

表1 珍珠豬毛菜根際和非根際土壤的理化性質Table 1 Physicochemical properties of GJ and NG
2.2 土壤細菌總數(shù)和可培養(yǎng)細菌數(shù)
對不同土壤的可培養(yǎng)細菌數(shù)、活細菌數(shù)和細菌總數(shù)的測定結果(圖1)表明,根際和非根際土壤可培養(yǎng)細菌數(shù)分別為1.62×106CFU·g-1和5.40×105CFU·g-1;活細胞數(shù)分別為 2.52×106個·g-1和2.26×106個·g-1;細菌總數(shù)分別為1.33×107個·g-1和1.12×107個·g-1。由此表明,珍珠豬毛菜根際土壤的可培養(yǎng)細菌數(shù)、活細菌數(shù)和總細菌數(shù)均高于周邊非根際土壤;且兩種土壤樣品的細菌總數(shù)和活細菌數(shù)均高于可培養(yǎng)細菌數(shù)。
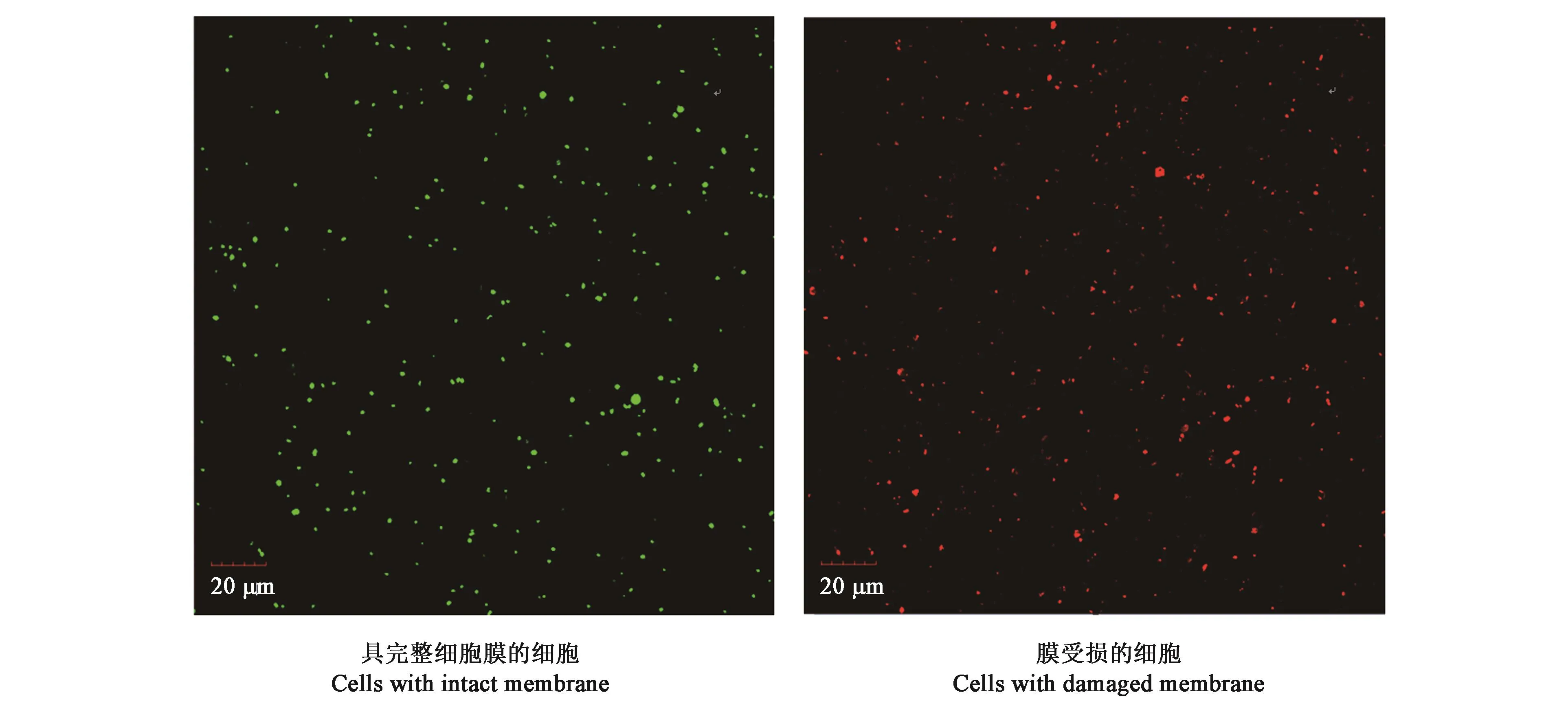
圖1 激光共聚焦顯微鏡下染色細胞圖Fig.1 Image of the stained cells with fluorescent dye under confocal laser microscope
2.3 土壤細菌群落多樣性分析
通過測序,珍珠豬毛菜根際和非根際土壤中OUT數(shù)分別為5 655和4 665個。其中,根際和非根際土壤特有的OUT數(shù)分別為2 580和1 590個,共同的為3 075個。多樣性分析結果(表2)表明,兩種土壤中細菌的Chao1指數(shù)無顯著差異;而珍珠豬毛菜根際土壤的Shannon指數(shù)顯著高于非根際土壤。非度量多維尺度(nonmetric multidimensional scaling,NMDS)分析(圖2)表明,根際和非根際土壤樣本被分離,表明兩種土壤的細菌群落組成具有明顯差異。

圖2 基于Weighted UniFrace NMDS分析的樣本二維排序圖Fig.2 NMDS plot based on weighted UniFrac distances

表2 土壤細菌的多樣性指數(shù)Table 2 Diversity indices of soil bacteria
2.4 不同土壤中細菌門組成
珍珠豬毛菜根際土壤和非根際土壤樣品中共檢測出33門細菌(圖3)。其中,放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、浮霉菌門(Planctomycetes)、芽單胞菌門(Gemmatimonadetes)、酸桿菌門(Acidobacteria)、綠彎菌門(Chloroflexi)和擬桿菌門(Bacteroidetes)等在2種土壤樣本中均有分布。放線菌門相對豐度最大,成為最優(yōu)勢菌門,在根際土壤中約占40.6%~43.3%,在非根際土壤中約占46.5%~52.0%。變形菌門、浮霉菌門和芽單胞菌門的相對豐度也較高。其中,浮霉菌門在根際土壤中的相對豐度(15.3%)顯著高于非根際土壤(9.1%)。而WPS-2和Latescibacteria為珍珠豬毛菜根際土壤特有的2個菌門;軟壁菌門(Tenericutes)和Halanaerobiaeota為非根際土壤特有的菌門。
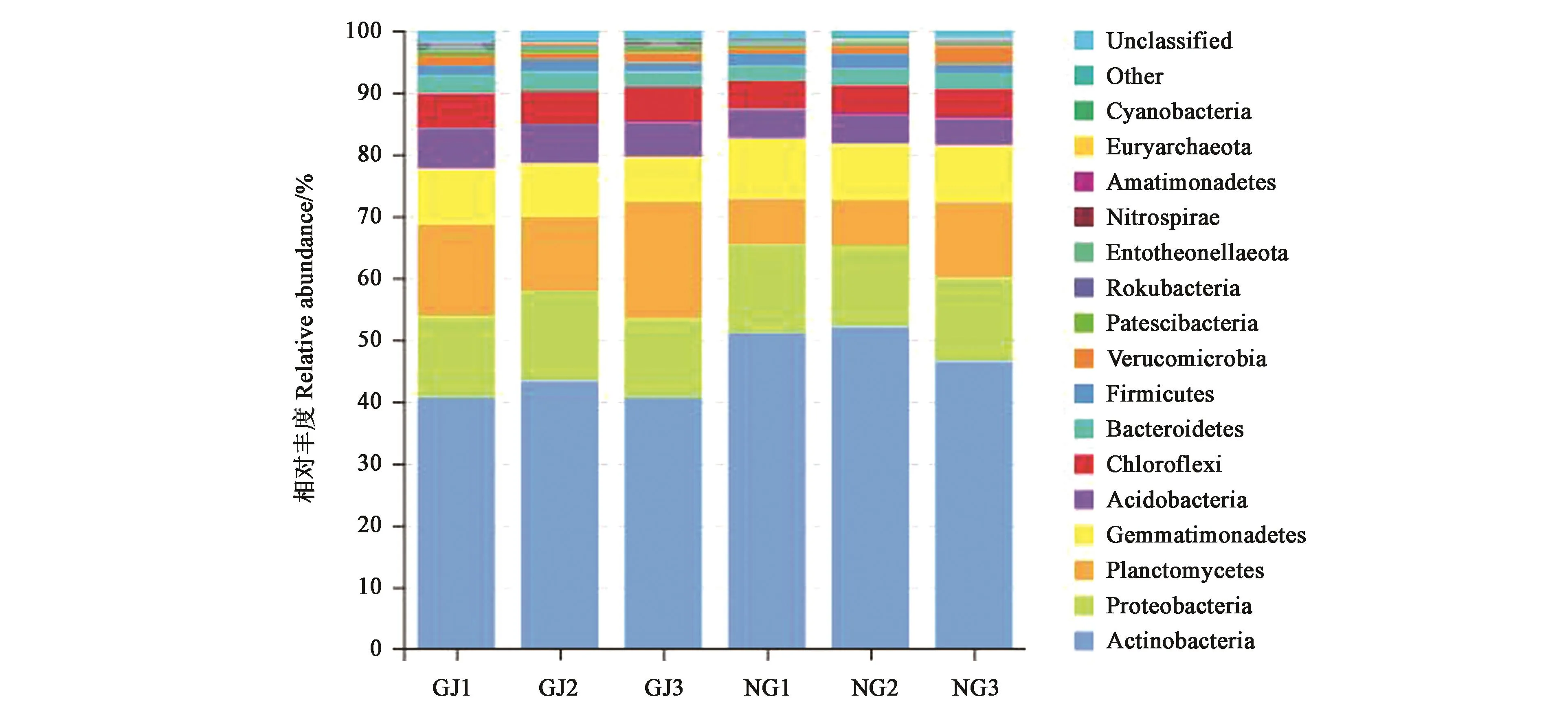
圖3 珍珠豬毛菜根際和非根際土壤細菌門水平的相對豐度Fig.3 Relative abundance of bacterial communities at the phylum level in different soils of Salsola passerine
2.5 不同土壤中細菌屬組成
珍珠豬毛菜根際土壤和非根際土壤分別包含367和398個細菌屬。其中,紅色桿菌屬(Rubrobacter) 、RB41、類 諾 卡 氏 菌 屬(Nocardioides)、鏈霉 菌 屬(Streptomyces)、Pir4_lineage、芽 孢 桿 菌 屬(Bacillus)、紅 桿 菌 屬(Solirubrobacter)、芽單胞菌屬(Gemmatimonas)、小梨形菌屬(Pirellula)假諾卡氏菌屬(Pseudonocardia)、Subgroup_10、分 支 桿 菌 屬(Mycobacterium)、半角藻屬(Haliangium)、MND1、Gaiella和Pelagibius在2種土壤中的相對豐度均較高(圖4)。其中,根際土壤相對豐度最高的優(yōu)勢菌屬為紅色桿菌屬,占3.8%;而非根際土壤的主要優(yōu)勢菌屬為類諾卡氏菌屬(Nocardioides),占2.9%。芽球菌屬(Blastococcus)和鞘氨醇單胞菌屬(Sphingomonas)在根際土壤中的相對豐度也較高(>1%);而擬無枝酸菌屬(Amycolatopsis)和糖霉菌屬(Glycomyces)在非根際土壤中的相對豐度較高(>1%)。
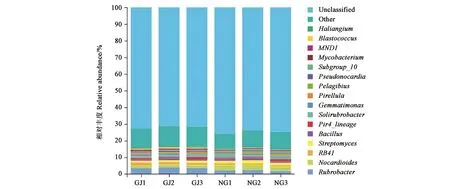
圖4 珍珠豬毛菜根際和非根際土壤細菌屬水平的相對豐度Fig.4 Relative abundance of bacterial communities at the genus level in different soils of Salsola passerine
2.6 土壤細菌與土壤理化性狀間的關系
2.6.1 土壤細菌數(shù)量與理化因子相關性分析對土壤細菌數(shù)量與土壤理化性狀進行相關性分析,結果(表3)表明,可培養(yǎng)細菌數(shù)量與土壤pH呈極顯著負相關,與其他理化性狀相關不顯著;而細菌總數(shù)與土壤養(yǎng)分含量、pH和土壤含水率相關均不顯著。
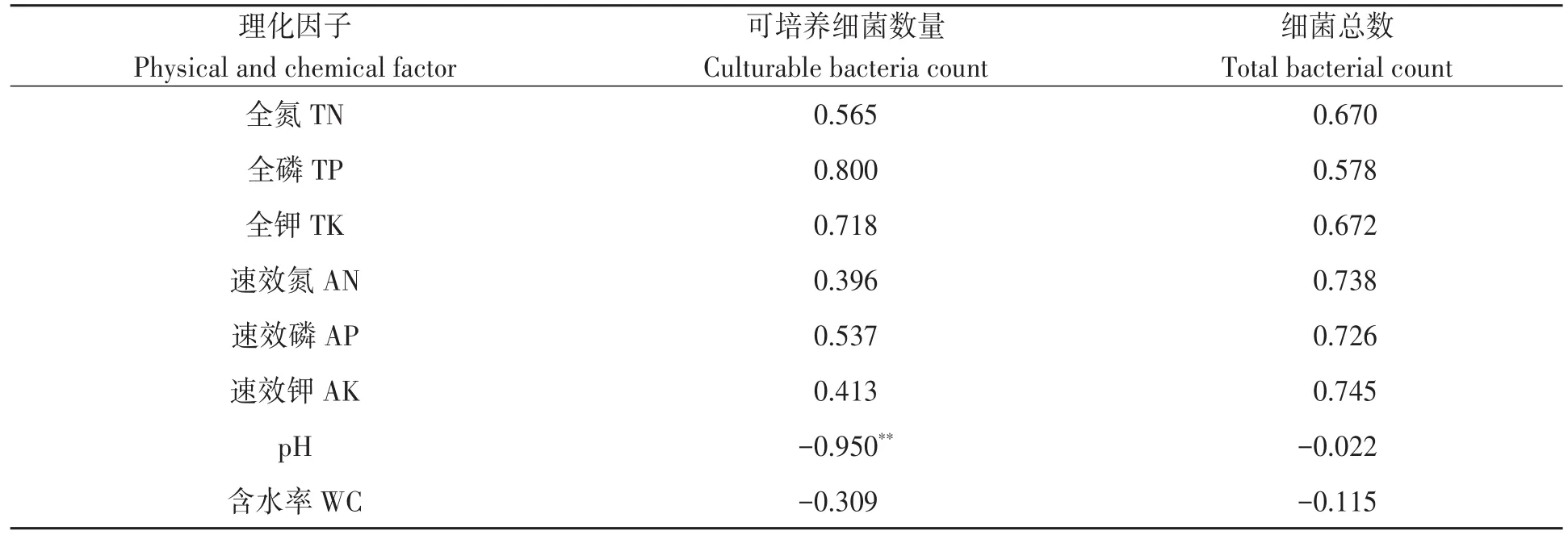
表3 土壤可培養(yǎng)細菌和細菌總數(shù)與理化因子之間的相關性Table 3 Correlation between the number of bacteria and soil physical and chemical properties
2.6.2 細菌多樣性指數(shù)與土壤理化因子相關性分析 對細菌多樣性與土壤理化性狀進行相關分析,結果(表4)表明,Shannon指數(shù)和Simpson指數(shù)與土壤速效氮、速效磷和速效鉀含量均達到極顯著正相關;Simpson指數(shù)與土壤全氮含量呈顯著正相關;ACE指數(shù)和Chao l指數(shù)均與土壤速效鉀含量呈顯著正相關。
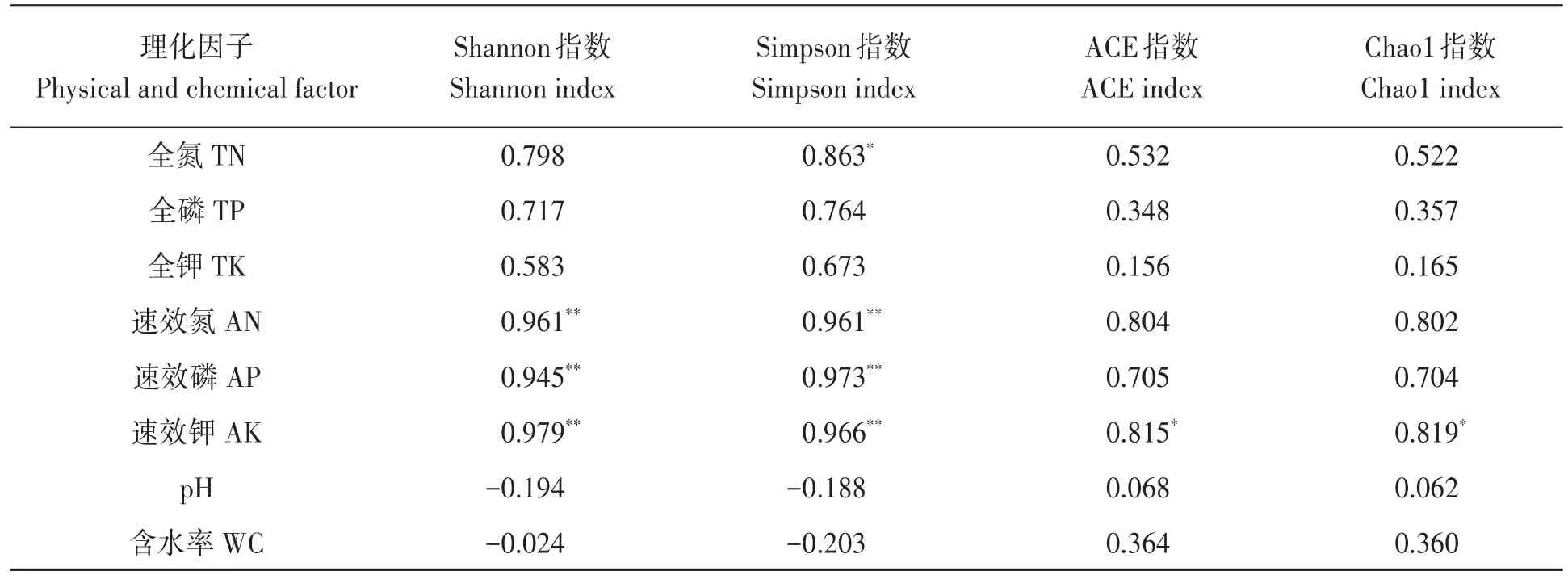
表4 細菌多樣性與土壤理化因子之間的相關性Table 4 Correlation between physical and chemical properties and diversity index of bacteria
2.6.3 細菌優(yōu)勢菌門與土壤理化因子相關性分析 對細菌優(yōu)勢菌門與土壤理化性狀進行相關性分析,結果(表5)表明,放線菌門的相對豐度與土壤全磷、速效氮、速效鉀含量呈顯著負相關,與土壤pH呈極顯著正相關;酸桿菌門與土壤全氮、速效氮、速效鉀含量呈顯著正相關,與土壤pH和含水率呈顯著負相關;綠彎菌門與土壤全氮和全磷含量呈顯著正相關,與土壤含水率呈顯著負相關,與速效氮和速效鉀含量呈極顯著正相關,與pH呈極顯著負相關。
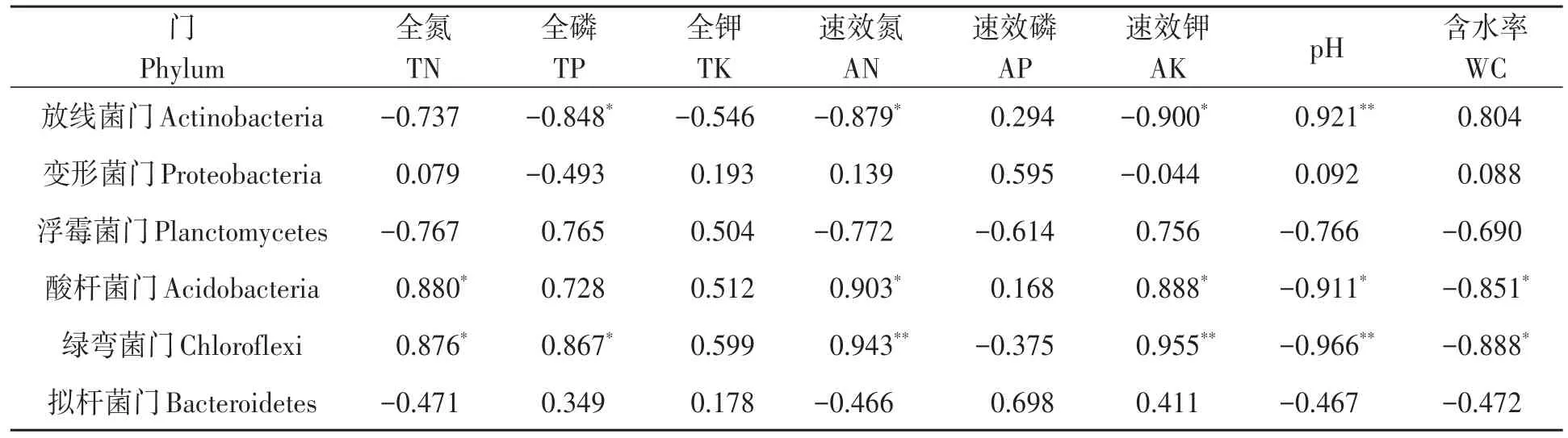
表5 不同土壤細菌優(yōu)勢菌門與土壤理化因子的相關性Table 5 Correlation of dominant communities of bacteria at the phylum level with soil physical and chemical factors
2.6.4 細菌優(yōu)勢菌屬與土壤理化因子相關性分析 對細菌優(yōu)勢菌屬與土壤理化性狀進行相關性分析,結果(表6)表明,細菌優(yōu)勢菌屬與不同土壤理化性狀具有一定的相關性。鏈霉菌屬與土壤pH呈顯著正相關。紅色桿菌屬與土壤全氮含量呈顯著正相關,與速效氮和速效鉀含量呈極顯著正相關,與土壤pH和含水率呈極顯著負相關。RB41與土壤速效氮和速效鉀含量呈顯著正相關,與土壤pH呈顯著負相關。類諾卡氏菌屬和Pelagibius與土壤全氮含量呈顯著負相關,與土壤含水率呈顯著正相關,與速效氮和速效鉀含量呈極顯著負相關,與土壤pH呈極顯著正相關。Pelagibius與土壤全磷含量呈顯著負相關。由此表明,土壤養(yǎng)分含量、pH和含水量是影響土壤細菌群落組成的重要因子。
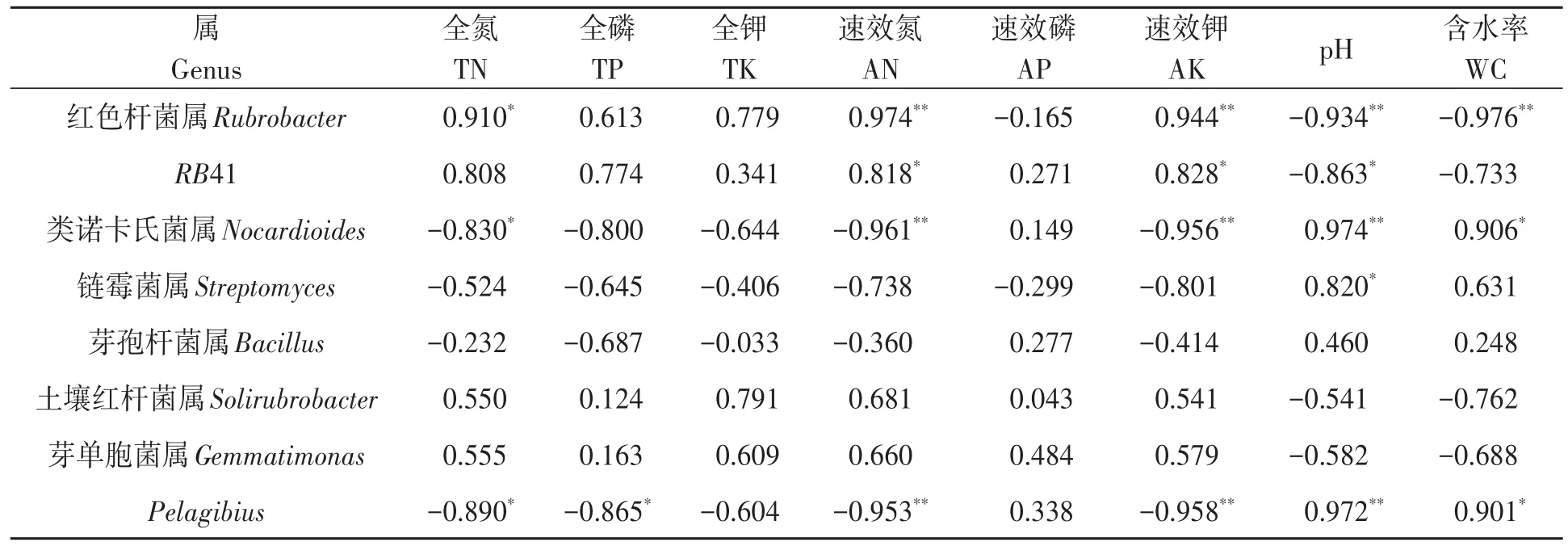
表6 不同土壤細菌優(yōu)勢菌屬與土壤理化因子的相關性Table 6 Correlation of dominant communities of bacteria at the genus level with soil physical and chemical factors
3 討論
景泰位于黃土高原與騰格里沙漠過渡地帶,為典型荒漠草原,土壤pH 8.23~8.93。研究表明,荒漠土壤pH顯著高于珍珠豬毛菜根際土壤,與楊陽等[10]研究結果相一致,可能是由于植物呼吸作用產生CO2及代謝作用產生的有機酸降低了土壤pH[11];而根際土壤含水率低于非根際土壤,可能與植物吸收水分有關。除全磷含量外,珍珠豬毛菜根際土壤養(yǎng)分含量均顯著高于非根際土壤,說明植物根系從環(huán)境攝取養(yǎng)分并促進了根際養(yǎng)分的積累,體現(xiàn)了“根際效應”有利于增加土壤養(yǎng)分[12]。根際微生物為植物提供養(yǎng)料和水分,增強植物對氮素攝入,促進土壤磷富集,有利于植物生長[13-14]。
珍珠豬毛菜根際土壤可培養(yǎng)細菌數(shù)為1.62×106CFU·g-1,活細胞數(shù)為 2.52×106個·g-1,細菌總數(shù)為1.33×107個·g-1;與非根際土壤相比,都有不同程度增加,表明根際土壤有利于微生物活動。值得注意的是,不同土壤的可培養(yǎng)細菌數(shù)量比活細菌數(shù)和細菌總數(shù)低,可能是在營養(yǎng)貧瘠的環(huán)境中存在著較多難培養(yǎng)細菌或不可培養(yǎng)細菌(Viable but Non-Culturable)[15-16],由此可見,純培養(yǎng)方法可能不能全面反映土壤微生物的類群和數(shù)量。
景泰荒漠草原土壤細菌具有豐富的多樣性,其中,荒漠土壤中細菌類群包含33門367屬,而珍珠豬毛菜根際土壤細菌類群涉及33門398屬。根際土壤和非根際土壤中OUT數(shù)、多樣性指數(shù)的顯著差異進一步驗證了兩種土壤中細菌在群落組成上存在差異。且根際土壤細菌的多樣性顯著高于非根際土壤,與前人研究結果相一致[17-18]。本研究發(fā)現(xiàn)放線菌門在兩種土壤中的豐度均最高,其中在非根際土壤中有更高的相對豐度(49.9%),可能是由于荒漠土壤為放線菌提供了木質素等營養(yǎng)物質[2]。放線菌門的許多屬種具有耐旱、耐鹽堿等特性,是干旱、鹽堿等類型土壤中的優(yōu)勢菌群[19-20],本研究表明,放線菌門與土壤理化因子間無顯著相關性。根際土壤中浮霉菌門的豐度僅次于放線菌門,其相對豐度從非根際土壤的9.1%增加到15.3%。酸桿菌門在根際土壤中的相對豐度從非根際土壤的4.7%增至6.2%,且與土壤pH呈顯著負相關,表明其更適應于pH較低的土壤環(huán)境[2]。綠彎菌門和擬桿菌門在珍珠豬毛菜根際土壤中的相對豐度亦高于非根際土壤;且綠彎菌門與土壤全氮、全磷、速效氮和速效鉀呈顯著正相關,與楊秉珣等[21]對川西北沙化地區(qū)的研究結果一致。植物根際可改變土壤微環(huán)境,使得某些細菌類群的豐度或多樣性增加或減少,形成有益于植物自身的根際細菌群落結構[22],如西瓜根際土壤中的優(yōu)勢類群是厚壁菌門、變形菌門和放線菌門[23];菊芋根際土壤中的優(yōu)勢類群是變形菌門、放線菌門、擬桿菌門和酸桿菌門[24],而造成不同植物根際土壤中優(yōu)勢菌群差異的機理尚需進一步研究。
在屬水平上,荒漠土壤主要優(yōu)勢菌屬為類諾卡氏菌屬,其次為紅色桿菌屬。紅色桿菌屬屬于放線菌門細菌,抗性較強,具有很好的固氮能力[25],與土壤全氮、速效氮和速效鉀含量呈顯著正相關,與土壤含水率和pH呈極顯著負相關。珍珠豬毛菜根際土壤的最優(yōu)勢菌為紅色桿菌屬,其次為RB41,芽單胞菌屬在根際土壤中相對豐度明顯增加,可能與多磷酸鹽積累有關[26],與土壤理化因子無顯著關系。
珍珠豬毛菜根際和非根際土壤的細菌群落結構存在顯著差異,土壤pH和含水率及全氮、全磷、速效氮、速效鉀含量是影響土壤細菌群落組成的主要因子,但這些菌群對珍珠豬毛菜生長的影響尚不清楚,故需進一步研究菌群的生長代謝,掌握細菌群-珍珠豬毛菜的協(xié)同規(guī)律和環(huán)境生存機制,為生態(tài)恢復提供理論依據(jù)。
