馬尾連堿乙抑制谷氨酰胺代謝影響胃癌細胞遷移及侵襲的機制
2022-07-15 03:12:48王洪濤梁育芩李海峰
河北醫藥 2022年13期
關鍵詞:胃癌
王洪濤 梁育芩 李海峰
胃癌為最常見的癌癥,是全世界癌癥相關死亡率的排名第二的誘因,也是東亞最常見的癌癥[1,2]。全世界每年約有99萬人被診斷出患有胃癌,其中約73.8萬人死于該疾病,使胃癌成為第四大常見癌癥,胃癌也是導致患者負擔最高的癌癥之一[3]。腫瘤細胞通過糖酵解而不是氧化磷酸化以獲得能量是腫瘤細胞的特征。這種代謝現象被稱為有氧糖酵解或“Warburg效應”。腫瘤細胞中增加的葡萄糖消耗、增加的糖酵解活性和乳酸的積累是腫瘤細胞的重要特征[4]。與主要通過線粒體氧化磷酸化產生能量的正常細胞相比,即使在有氧條件下,腫瘤細胞也主要通過增加糖酵解獲得能量。乳酸的積累形成酸性微環境,對腫瘤細胞有保護作用[5]。此外在過去30年中,越來越多的證據表明谷氨酰胺是許多腫瘤細胞的重要代謝底物和能量來源,這些腫瘤細胞需要谷氨酰胺才能持續生長和存活,表現出所謂的“谷氨酰胺成癮”。谷氨酰胺代謝不僅為腫瘤細胞提高能量,同時為其提供細胞增殖的原料[6]。馬尾連為毛茛科唐松草屬植物多葉唐松草Thalictrum foliolosum DC的根莖,該藥具有清熱燥濕,瀉火解毒的作用,用于腸炎、痢疾、黃疸、目赤腫痛,生物堿是其主要活性成分,近年研究發現馬尾連提取物及其生物總堿對小鼠肺癌的抑制率達74%,對腹水淋巴瘤小鼠生命延長率為137.2%[7,8],研究發現馬尾連生物堿主要為甲、乙、丙三類,其中馬尾連生物堿甲可阻斷G1細胞向S期過渡,其殺細胞作用似為細胞周期特異性,生物堿乙及丙也具有一定的抗腫瘤作用[9,10],筆者總結后發現研究較少有待進一步明確其作用。本研究擬探討馬尾連堿乙在胃癌中的作用及其對胃癌細胞能量代謝的影響,報告如下。
1 材料與方法
1.1 材料與主要儀器 人胃癌SGC-7901、BGC-823 細胞系購自中國科學院上海細胞研究所,馬尾連堿乙(SIGMA,美國),四甲基偶氮唑鹽(MTT,95%,SIGMA 公司美國);二甲基亞砜(DMSO,百靈威試劑公司);乳酸含量試劑盒,葡萄糖試劑盒(Bivision公司,英國),ATP檢測試劑盒(南京建成生物工程研究所,A095-1-1,中國),谷氨酰胺試劑盒(南京建成生物工程研究所,A047-1-1,中國),實驗其他試劑均為化學純。主要儀器為蛋白印跡系統(Bio-rad公司,美國),多功能酶標儀(sigma公司,美國)。
1.2 方法
1.2.1 正常細胞模型:采用人胃癌SGC-7901,BGC-823細胞,將不同細胞系37℃條件下,采用含有10%小牛血清培養基培養于二氧化碳培養箱,培養基為采用85 U/L青霉素、100 mg/L 鏈霉素的DMEM。將細胞接種于96孔板中,設立復孔,分為4組,空白組細胞正常培養基培養,馬尾連堿乙組(濃度為5 μmol/L、25 μmol/L、75 μmol/L)。采用MTT法觀察馬尾連堿乙對胃癌細胞活力,遷移,侵襲能力的影響。
1.2.2 谷氨酰胺剝奪細胞模型:建立谷氨酰胺剝奪細胞模型,去除正常DMEM,用PBS一次性沖洗或轉染正常細胞。添加不含谷氨酰胺的DMEM(Thermo Fisher Science,A14431)。此外,向培養基中添加10%的DFBS(美國加利福尼亞州薩克拉門托,Gemini Bio Products)。細胞在此培養基中培養24 h。細胞適應培養基后,正常傳代,繼續使用。
1.3 MTT抑制率實驗 將不同的胃癌細胞SGC-7901,BGC-823分別接種在96孔板中,濃度為1×105個細胞,24 h 后加入含有馬尾連堿乙的培養液繼續培育,每組6孔。12 h、24 h、48 h后,加入30 μl MTT繼續在培養箱中孵育4 h,加入150 μl DMSO溶解結晶物。采用酶標儀在550 nm下檢測吸光度(A)值,計算細胞活力。
1.4 三磷酸腺苷(ATP)及谷氨酰胺含量實驗 采用ATP及谷氨酰胺試劑盒檢測細胞培養基中的乳酸含量,將不同的胃癌細胞SGC-7901、BGC-823分別接種在6孔板中,濃度6×107個細胞,24 h后加入含有馬尾連堿乙的培養液繼續培育24 h后檢測,每組3孔。ATP及谷氨酰胺檢測方法按照試劑盒說明書進行。
1.5 葡萄糖消耗及乳酸含量實驗 采用葡萄糖及乳酸試劑盒檢測細胞培養基中的乳酸含量,將不同的胃癌細胞SGC-7901、BGC-823分別接種在6孔板中,濃度6×107個細胞,24 h 后加入含有馬尾連堿乙的培養液繼續培育24 h后檢測,每組3孔。葡萄糖及乳酸試劑盒檢測方法按照試劑盒說明書進行。
1.6 細胞遷移實驗 置于24孔板中浸泡1 h,將不同的胃癌細胞SGC-7901、BGC-823于對數生長期消化,計數制備細胞懸液,加入24孔板中,每孔加入100 μl細胞懸液,24孔板中加入Transwell,加入藥物干預,37℃孵育24 h,取出Transwell,用PBS洗滌2次,用0.4%多聚甲醛在4℃固定30 min,用PBS洗滌2次,室溫下用0.1%結晶紫染色15~30 min,用PBS洗滌2次,顯微鏡下觀察計數。
1.7 細胞侵襲實驗 用無血清培養基將基質凝膠稀釋至最終濃度1 mg/ml。在Transwell培養箱中,向上培養箱中加入100 μl基質凝膠,在37℃下培養4~5 h,使其凝膠化。在上腔分別加入的胃癌細胞。下室加入含血清培養基600 μl,孵育2 h后取出培養箱,甲醇固定30 min,Giemsa染色30 min,隨機抽取9個視野進行顯微鏡下計數。
1.8 蛋白免疫印跡實驗 分別將胃癌細胞SGC-7901和BGC-823接種于6孔板中,濃度為5×106個細胞,給藥干預24 h后,提取細胞蛋白,BCA試劑盒測定蛋白濃度。加入50 ng樣品蛋白,電泳分離后,用濕轉移印花法將蛋白轉移到PVDF膜上,用含1%脫脂奶粉和0.1%TBST的TBS密封過夜,TBST洗滌3次,每次5 min,加入一抗體,4℃孵育過夜,TBST洗滌3次,每次5 min。加入辣根過氧化物酶(1∶2 000)標記的二抗體,室溫孵育2 h,TBST洗滌膜,每次5 min。以GAPDH為內參照。用ECL化學發光試劑顯色。
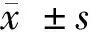
2 結果
2.1 馬尾連堿乙顯著抑制胃癌細胞活力 給予中、高劑量馬尾連堿乙干預后,SGC-7901細胞在干預24 h及48 h后細胞活力與對照組比較差異均有統計學意義(P<0.05),中劑量馬尾連堿乙干預BGC-823細胞48 h后,細胞活力與對照組比較差異有統計學意義(P<0.05),高劑量馬尾連堿乙干預24 h后,兩種胃癌細胞活力與對照組比較差異有統計學意義(P<0.05)。見表1。
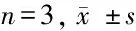
表1 馬尾連堿乙對胃癌細胞活力的影響
2.2 馬尾連堿乙抑制胃癌細胞的遷移及侵襲能力 給予馬尾連堿乙后,細胞遷移及侵襲能力增加,細胞穿膜數量明顯增加,其中SGC-7901細胞中,給予中、高劑量的馬尾連堿乙干預后,遷移及侵襲細胞穿膜數與對照組比較差異均有統計學意義(P<0.05)。BGC-823細胞給予中、高劑量馬尾連堿乙干預后,與對照組比較,差異有統計學意義(P<0.05),高劑量組干預后,遷移及侵襲細胞穿膜數與對照組比較,差異有統計學意義(P<0.05)。見表2。
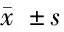
表2 馬尾連堿乙對胃癌細胞遷移和侵襲實驗中的穿膜細胞數的影響 n=3,個,
2.3 馬尾連堿乙顯著抑制胃癌細胞能量代謝水平及谷氨酰胺含量 給予馬尾連堿乙后,馬尾連堿乙可以明顯抑制細胞的能量代謝水平,在SGC-7901細胞中,馬尾連堿乙中、高濃度均可以顯著抑制細胞能量代謝水平,同時顯著抑制細胞中谷氨酰胺含量,與對照組比較差異有統計學意義(P<0.05);在BGC-823細胞中,給予馬尾連堿乙干預后,細胞能量代謝水平明顯減低,中、高劑量組與對照組比較差異有統計學意義(P<0.05),中、高劑量組干預后,均可以抑制BGC-823細胞中的谷氨酰胺含量,差異有統計學意義(P<0.05)。見表3。
表3 馬尾連堿乙對胃癌細胞ATP及谷氨酰胺含量的影響
2.4 馬尾連堿乙對胃癌細胞糖酵解水平的影響 給予馬尾連堿乙干預后,對兩種胃癌腫瘤細胞的乳酸含量及葡萄糖消耗水平具有一定的影響,但與對照組比較差異無統計學意義(P>0.05)。見表4。
表4 馬尾連堿乙對胃癌細胞糖酵解水平的影響
2.5 馬尾連堿乙抑制胃癌細胞谷氨酰胺酶的表達 蛋白實驗結果表明,馬尾連堿乙對SGC-7901、BGC-823胃癌腫瘤細胞谷氨酰胺酶表達均具有明顯的抑制作用,且隨計量的升高作用更加顯著。見圖1。

圖1 馬尾連堿乙在不同細胞中抑制胃癌細胞谷氨酰胺酶的表達
2.6 馬尾連堿乙在無谷氨酰胺環境下對胃癌細胞活力抑制作用 將兩種不同的胃癌細胞接種于無谷氨酰胺的培養基中,實驗結果表明,馬尾連堿乙對細胞活力的抑制作用顯著降低,其中僅在高劑量組對SGC-7901在干預48 h后,具有一定抑制作用,且差異有統計學意義(P<0.05)。見表5。
表5 馬尾連堿乙在無谷氨酰胺環境下對胃癌細胞活力的影響
2.7 馬尾連堿乙在無谷氨酰胺環境下對胃癌細胞遷移及侵襲能力的影響 將兩種不同的胃癌細胞接種于無谷氨酰胺的培養基中,實驗結果表明,馬尾連堿乙對細胞的遷移及侵襲能力影響作用顯著降低,其中在高劑量組對SGC-7901的遷移實驗具有抑制作用,差異有統計學意義(P<0.05)。見表6。
表6 在無谷氨酰胺環境下馬尾連堿乙對胃癌細胞遷移和侵襲的穿膜細胞數的影響 n=3,個,
2.8 馬尾連堿乙在無谷氨酰胺環境下對胃癌細胞ATP含量的影響 將兩種不同的胃癌細胞接種于無谷氨酰胺的培養基中,實驗結果表明,馬尾連堿乙對細胞的能量代謝水平無顯著影響,不同濃度的馬尾連堿乙干預后,細胞的能量代謝水平與對照組比較差異均無統計學意義(P>0.05)。見表7。
表7 在無谷氨酰胺環境下馬尾連堿乙對胃癌細胞ATP含量的影響
3 討論
胃癌是消化系統最常見的惡性腫瘤之一。外科手術目前被認為是唯一的根治性治療。隨著手術技術的進步和傳統放療、化療及新輔助治療的實施,早期胃癌的5年生存率可達95%以上[1]。然而,早期診斷率低意味著大多數患者在診斷時都有晚期疾病,因此錯過了最佳手術窗口。因此輔助放化療、分子靶向治療和免疫治療是晚期胃癌的主要治療方法。
癌細胞的新陳代謝與正常細胞有很大的不同。糖酵解和谷氨酰胺分解途徑在癌細胞中的能量供應中均顯著增強。當糖酵解增強以滿足癌細胞不斷增加的能量需求時,谷氨酰胺分解增強以提供癌細胞的生物合成前體[11]。
本研究發現馬尾連堿乙可以顯著抑制胃癌細胞的增殖活力,同時降低胃癌細胞的遷移及侵襲能力,給予馬尾連堿乙可以明顯降低胃癌細胞ATP合成能力,但其不影響細胞乳酸及葡萄糖消耗水平。糖酵解是腫瘤細胞能量代謝的主要來源,而谷氨酰胺同樣參與腫瘤細胞的的能量代謝,谷氨酰胺是一種豐富的多功能營養素,參與癌細胞的能量形成、氧化還原穩態、大分子合成和信號傳導。谷氨酰胺作為營養素在癌癥中的重要性源于它可以將氮和碳提供給腫瘤促進腫瘤細胞增殖[12,13]。谷氨酰胺是血漿中最豐富的氨基酸之一。盡管大多數組織可以合成谷氨酰胺,但在快速生長或其他應激期間,谷氨酰胺成為腫瘤細胞快速增殖的關鍵因素[14,15]。
本研究發現馬尾連堿乙隨對糖酵解反應沒有顯著干預作用,但可顯著抑制胃癌細胞谷氨酰胺代謝能力,降低腫瘤細胞中的谷氨酰胺含量。體內循環的谷氨酰胺通過轉運蛋白如SLC1A5進入細胞。癌細胞通過谷氨酰胺酶介導的谷氨酰胺分解將谷氨酰胺轉化為谷氨酸,可被谷氨酸脫氫酶(GLUD)或谷氨酸轉氨酶進一步脫氨基為α-酮戊二酸(α-KG),進入三羧酸(TCA)循環,α-酮戊二酸是TCA循環的中間產物,也是修飾蛋白質和DNA的雙加氧酶的底物。近年研究發現,沉默GLS表達或抑制GLS活性可以明顯延緩腫瘤生長[16-18]。有研究發現GLS2是p53靶基因,在腫瘤抑制中起發揮作用。近期的研究發現上皮細胞-細胞粘附分子E-cadherin參與腫瘤細胞誘導遷移、侵襲,其機制與谷氨酰胺代謝相關[19,20]。
本研究發現馬尾連堿乙可以顯著抑制谷氨酰胺酶的表達,當將馬尾連堿乙給予缺乏谷氨酰胺培養基的胃癌細胞后其對腫瘤細胞的調節能力消失,其對腫瘤細胞的抑制作用顯著降低,同時其對細胞遷移,侵襲的能力顯著降低。
綜上所述,馬尾連堿乙可以抑制谷氨酰胺酸的表達,影響胃癌細胞谷氨酰胺代謝,抑制其能量代謝水平,從而發揮抑制胃癌細胞增殖活力及減少遷移及侵襲的作用,但其具體的調控分子機制還有待于進一步的研究。
猜你喜歡
昆明醫科大學學報(2022年1期)2022-02-28 07:43:36
昆明醫科大學學報(2021年5期)2021-07-22 07:32:22
基層中醫藥(2020年2期)2020-07-27 02:46:06
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
中外醫療(2015年18期)2016-01-04 06:51:55
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:25
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫藥(2015年20期)2015-03-01 02:04:29
