榛子胚珠發育相關基因ChARF1的原核表達及多克隆抗體制備
2022-07-23 07:00:24黃立龍王秋緣劉春明劉劍鋒程云清
東北師大學報(自然科學版) 2022年2期
關鍵詞:檢測
黃立龍,魏 珩,王秋緣,劉春明,劉劍鋒,程云清
(1.吉林師范大學吉林省植物資源科學與綠色生產重點實驗室,吉林 四平136000;2.吉林師范大學生命科學學院,吉林 四平136000)
榛子(Corylusspp.)屬于樺木科(Betulaceae)榛屬(CorylusL.),是重要的經濟林木.隨著我國退耕還林政策的實施推廣,榛子已經成為山區經濟發展的優勢樹種.榛子種仁可直接食用或制成榛子油、榛子醬及利口酒等加工食品[1-2].榛子的營養價值和經濟價值主要來自于種仁.榛子種仁是否飽滿與胚珠發育過程是否能順利進行具有直接關系,而在實際種植中,榛子的胚珠發育障礙問題普遍存在,導致種仁干癟和空殼現象,造成種仁營養和經濟價值的損失[3].因此,深入了解榛子花果發育尤其是胚珠發育的分子機制,對于改良榛子種質,提升質量、提高產量具有重要意義.
生長素(auxin)是植物生長發育中的一種核心調控激素,在營養生長和生殖生長過程中具有重要調節作用.生長素反應因子(auxin response factor,ARF)通過與生長素/吲哚-3-乙酸(auxin/indole-3-acetic acid,AUX/IAA)相互作用實現對生長素的響應和信號調控,是調控下游響應基因的重要轉錄因子[4-5].ARFs的功能研究在模式植物擬南芥(Arabidopsisthaliana)中比較成熟,參與花果發育(AtARF1/2/3/5/6/8/17)[6-8]、葉片生長(AtARF3/4)[9]、根系生長(AtARF7/10/16/19)[10-11]和種子發育(AtARF12/13/14/15/10/21/22)[10]等多種過程[12],其中AtARF1/2在花的發端和衰老過程中具有重要作用[6].同時ARFs在植物抗逆境脅迫的調控中也扮演重要角色[13-14].目前,在對榛子進行的相關研究中,未見關于ARFs參與胚珠發育乃至花果發育的報道.本文通過對前期研究[15]的榛子胚珠發育轉錄組數據進行篩選,發現了1個顯著差異表達ARF同源基因,命名為ChARF1,通過構建原核表達載體獲得了ChARF1(N-380)多克隆抗體,為了解ChARF1基因在榛子胚珠發育中的作用提供了檢測材料,研究結果可為進一步發現ARFs在榛子花果發育中的功能機制奠定基礎.
1 材料與方法
1.1 樣品采集
實驗材料為平歐雜交榛(Corylusheterophylla×C.avellana),種植于吉林省四平市伊通滿族自治縣榛園,品種為“達維”,樹齡12年,株高3.0~3.5 m.2019年,收集榛子品種“薄殼紅”花粉并參照本實驗室的研究方法人工授粉[3].于同年8月20日(授粉后120 d),采集果實并現場解剖,取成熟期胚珠置液氮中,轉至實驗室后超低溫冰箱-80℃保存[15].
1.2 基因篩選及克隆
以前期榛子胚珠發育4個時期(Ov1:胚珠形成期,Ov2:早期胚珠生長期,Ov3:胚珠快速生長期和Ov4:胚珠成熟期)轉錄組測序數據[15]為基礎,篩選差異表達基因.相對表達量以FPKM值(Fragments per kilobase of transcript per million fragments mapped)為指標,篩選標準:|log2(倍數變化)|>1且錯誤發現率<0.05.使用SPSS 8.0 方差分析程序進行差異顯著性分析(3次生物學重復).使用Primer Premier 5.0平臺設計CDS全長序列引物(F:5′-AGAGAGATGAGTAGTGTTGG-3′,R:5′-ATTGGAATCTAATAGGGATT-3′,產物長度2 129 bp).樣品總RNA提取和檢測參考Liu 等[15]的方法.以純度和濃度符合標準的總RNA反轉錄合成cDNA第一條鏈,參照SuperScriptTMIV第一鏈合成系統試劑盒(Invitrogen,USA,18091050)說明書進行操作.使用設計的引物以cDNA為模板擴增篩選基因,純化后以T4DNA連接酶(寶生物工程有限公司,大連,2040A)連接至pMD18-T載體(寶生物工程有限公司,大連,D103A)并轉化大腸桿菌DH5α(唯地生物技術有限公司,上海,DL1001).提取陽性克隆質粒留存并送上海生工有限公司測序.
1.3 基因鑒定及系統發育分析
測序所得序列在NCBI(nucleotide collection nr/nt)和番茄數據庫(https:∥phytozome.jgi.doe.gov/pz/portal.html)進行比對查找同源序列.序列翻譯為蛋白后,使用NCBI保守序列數據庫CDD(https:∥www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)和Pfam(http:∥pfam.sanger.ac.uk/)驗證序列的保守結構域[16],隨后通過DNAMAN 9.0.1軟件進行結構域比對并作圖.使用在線工具ProtParam[17](https:∥web.expasy.org/protparam/)預測蛋白相對分子質量.將比對得到的同源序列以及Tair(https:∥www.arabidopsis.org/)網站下載的擬南芥編碼序列(Coding sequence,CDS)通過MEGA 7.0[18]軟件構建最大似然(maximum likelihood,ML)樹,進行系統發育分析[19].
1.4 原核表達載體構建及誘導表達
將包含目的基因的pMD18-T重組質粒送至武漢愛博泰克生物科技有限公司構建原核表達載體并制備多克隆抗體.以重組質粒為模板PCR擴增目的基因N端特異性片段,純化克隆至pET-28a-SUMO載體(含His-標簽,T7-標簽,SUMO-標簽,相對分子質量約18 000).經PCR和測序鑒定后,提取陽性重組質粒轉化E.coliRosetta菌株.將陽性單克隆擴大培養至D(600)=0.5~0.6.設實驗組:加入0.8 mmol/L IPTG,37 ℃誘導4 h;對照組:加入等量無菌水.收集少量菌液進行抗原蛋白表達SDS-PAGE(聚丙烯酰胺凝膠電泳)檢測.驗證表達成功后,將剩余菌液離心重懸并用超聲波破碎,離心并分離上清和沉淀.使用2和8 mol/L尿素溶解沉淀中的包涵體,期間取上清和溶解包涵體進行抗原蛋白可溶性SDS-PAGE檢測.相同條件下進一步擴大培養,收集包涵體蛋白,使用組氨酸標簽蛋白純化試劑盒(碧云天生物技術有限公司,上海)純化后進行SDS-PAGE檢測和濃度測定.
1.5 抗體制備和純化及蛋白質免疫印跡檢測
將純化后的抗原蛋白分5次免疫兩只實驗級雄性日本大耳白兔,具體流程見表1.酶聯免疫效價檢測:設空白對照;陰性對照:免疫前抽取兔耳靜脈血清;陽性實驗:第52天抽取兔頸部抗血清.酶標板每孔加入200 ng純化抗原蛋白和不同稀釋濃度 (體積比為1∶1 000~1∶512 000) 的抗血清,加入二抗進行顯色反應,檢測D(450)進行效價分析.構建抗原蛋白與瓊脂糖耦連的抗原親和純化層析柱,抗血清上樣洗脫后得到純化濃縮抗體.以ChARF1(N-380)蛋白作抗原,分別取500 pg,10 ng進行聚丙烯酰胺凝膠電泳.抗體1∶1 000(體積比)倍稀釋后做蛋白質免疫印跡檢測[20].
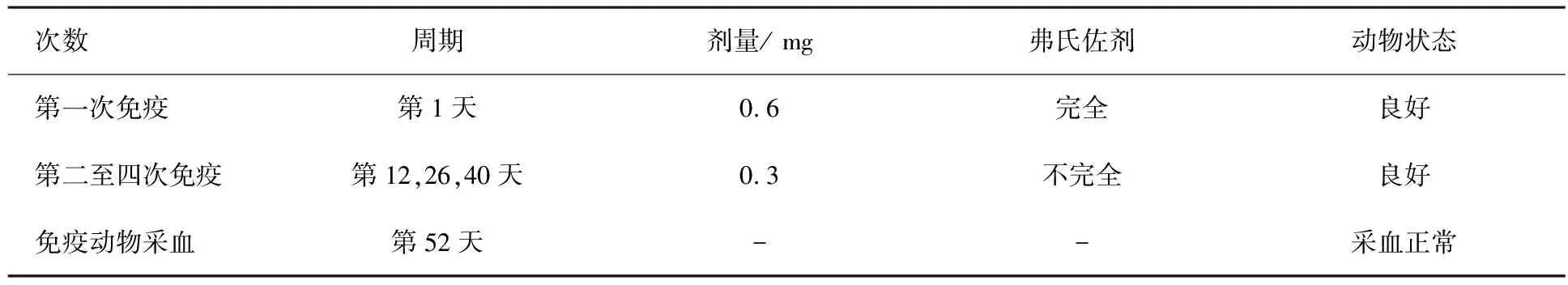
表1 免疫流程
2 結果與分析
2.1 目的基因的鑒定與進化分析
通過對比轉錄組[15]中胚珠4個時期的相對表達量(FPKM值),發現1個Ov4顯著上調表達基因(unigene)g22071(P<0.05,見圖1A).該基因CDS長度為2 115 bp,編碼704個氨基酸,預測編碼蛋白相對分子質量為78 770.通過核苷酸序列比對程序篩選同源序列以及保守結構域預測,發現該基因同源聚類于胡桃(Juglansregia,XM_035695460.1)、闊葉櫟(Quercuslobata,XM_031092776.1)、歐洲栓皮櫟(Quercussuber,XM_024058295.1)和扁桃(Prunusdulcise,XM_034371489.1),參考ARF基因序列(E=0,重合率(identity)>75%),顯著匹配Auxin_resp(pfam06507)、B3(pfam02362)和不顯著匹配AUX_IAA(CL0072)結構域[4]見圖1B,鑒定g22071屬于ARF基因家族,命名為ChARF1(Genbank登錄號:MW369441).
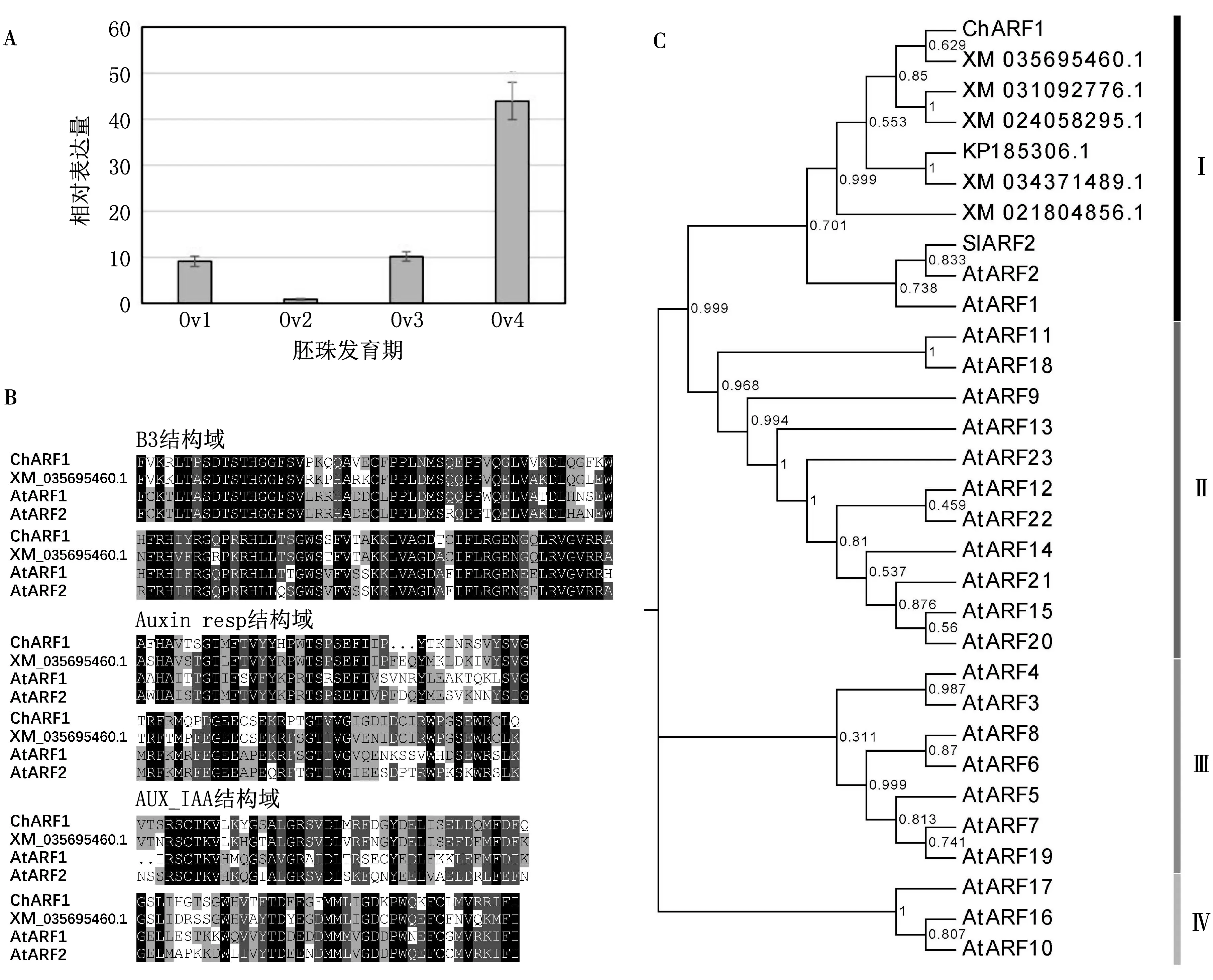
A:g22071基因表達分析.相對表達量以FPKM值為標準;a—c表示差異表達顯著水平,P<0.05.B:保守結構域分析.C:系統發育分析.AtARF1-23,擬南芥(Arabidopsis thaliana);ChARF1,榛子(Corylus heterophylla × C.avellana);SlARF2,番茄(Solanum lycopersicum);XM_035695460.1,胡桃(Juglans regia);XM_031092776.1,闊葉櫟(Quercus lobata);XM_024058295.1,歐洲栓皮櫟(Quercus suber);KP185306.1,梅(Prunus mume);XM_034371489.1,扁桃(Prunus dulcise);XM_021804856.1,橡膠樹(Hevea brasiliensis).
將ChARF1及核苷酸序列比對獲得的同源序列和模式植物擬南芥ARF基因家族共31條序列構建ML進化樹,結果(見圖1C)顯示,31條不同物種的ARFs聚類為4個進化分枝(Ⅰ—Ⅳ).ChARF1基因與胡桃、闊葉櫟等的ARFs以及擬南芥AtARF1/2、番茄SlARF2基因聚類于分枝Ⅰ.在分枝Ⅰ內,ChARF1基因與胡桃的ARF(XM_035695460.1)同源性最高,而與已報道的擬南芥AtARF1/2基因[6-8]和番茄SlARF2基因[21]同源關系相對較遠.擬南芥AtARF3-23基因則聚類于Ⅱ,Ⅲ,Ⅳ分支.
2.2 原核表達載體的構建及表達
選擇ChARF1基因序列N端包含Auxin_resp和B3保守結構域(見圖1B)的前380個氨基酸為抗原特異性片段,以測序正確的pMD18-T-ChARF1質粒為模板進行PCR擴增,聚丙烯酰胺凝膠電泳檢測產物大小正確(見圖2A).構建pET-28a-SUMO-ChARF1(N-380)重組載體后,測序鑒定結果顯示目的片段成功克隆至重組載體,載體構建成功.
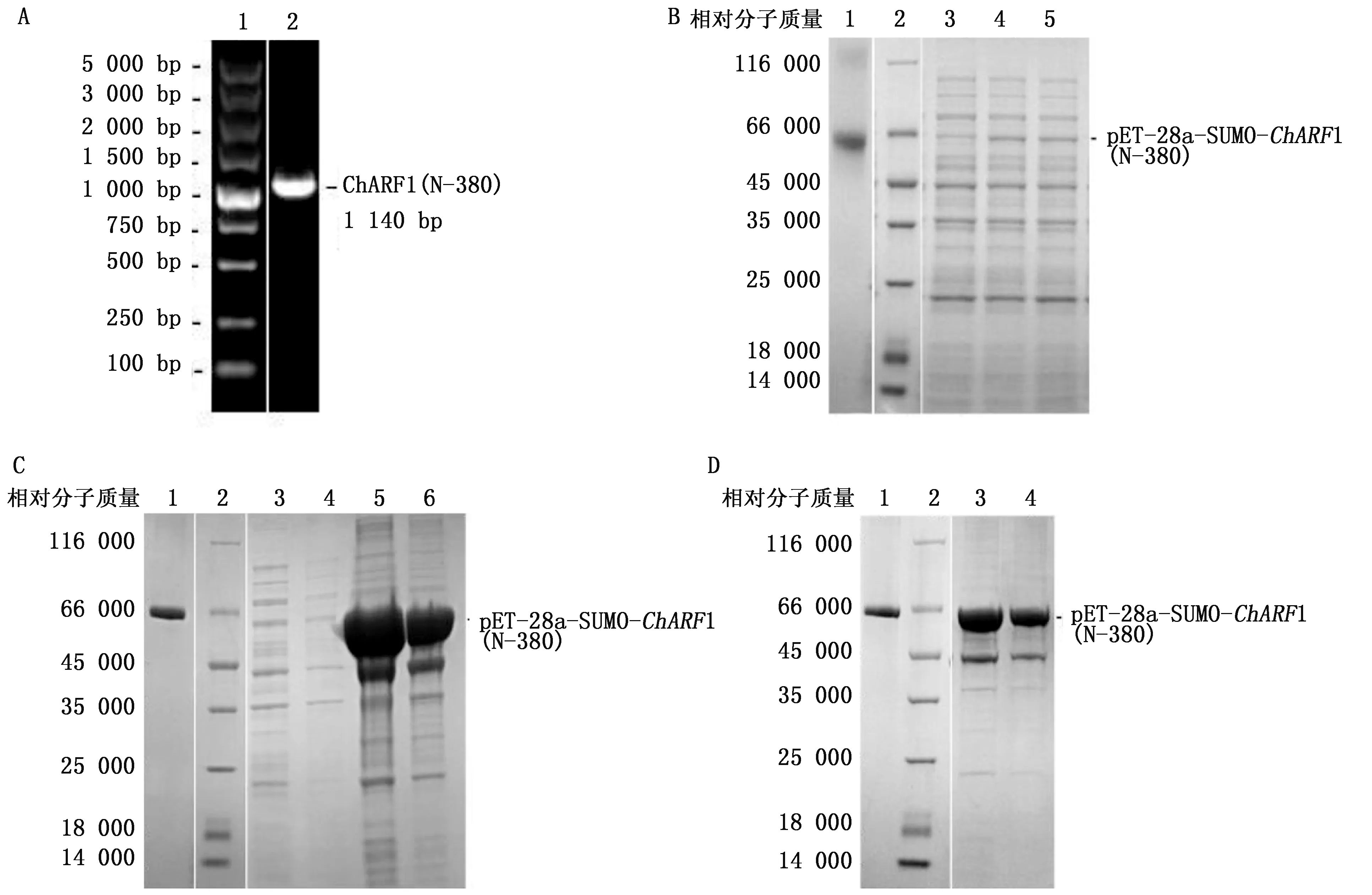
A:PCR擴增結果.1:Marker(相對分子質量標準),2:ChARF1(N-380).B:蛋白表達檢測.1:0.4 mg/mL 牛血清白蛋白,2:Maker,3:pET-28a-SUMO-ChARF1(N-380)未誘導表達,4:pET-28a-SUMO-ChARF1(N-380)誘導表達,5:pET-28a-SUMO-ChARF1(N-380)誘導表達.C:可溶性檢測.1:0.4 mg/mL 牛血清白蛋白,2:Marker,3:上清,4:上清2(2 mol/L尿素溶解),5:包涵體2倍稀釋(8 mol/L尿素溶解),6:包涵體5倍稀釋(8 mol/L尿素溶解).D:純化檢測.1:0.4 mg/mL 牛血清白蛋白,2:Marker,3:包涵體10倍稀釋(8 mol/L尿素溶解),4:包涵體20倍稀釋(8 mol/L尿素溶解).
將陽性pET-28a-SUMO-ChARF1(N-380)載體轉化至E.coliRosetta菌株進行表達.完整的ChARF1蛋白相對分子質量大小推測為78 770,而ChARF1(N-380)相對分子質量推測為43 000.重組蛋白融合了相對分子質量為18 000的His-標簽、T7-標簽、SUMO-標簽,實際表達產物預期相對分子質量高于61 000.聚丙烯酰胺凝膠電泳結果顯示,實驗組與對照組相比,在相對分子質量為66 000的條帶處可觀察到特異性條帶(見圖2B),符合預期大小,表明ChARF1(N-380)在E.coliRosetta菌株中成功表達.
2.3 抗原蛋白的可溶性檢測及純化
經超聲波破菌后,取上清和沉淀進行ChARF1(N-380)抗原蛋白可溶性檢測,結果(見圖2C)顯示在相對分子質量為66 000處,上清及上清2(2 mol/L尿素溶解包涵體),條帶不清晰;而在沉淀中,通過使用8 mol/L尿素溶解包涵體,可觀察到明顯的條帶.表明ChARF1(N-380)抗原蛋白表達主要在包涵體中,具有不溶性.
抗原蛋白ChARF1(N-380)具有6個氨基酸的His-標簽,可使用His標簽蛋白純化試劑盒純化.純化前非特異性條帶較多(見圖2C);純化后非特異性條帶基本消失,在相對分子質量為66 000處抗原蛋白條帶清晰(見圖2D).依據BSA蛋白條帶進行蛋白濃度估算約為12 mg/mL,包涵體20倍稀釋液仍可觀察到明顯條帶.結果證明,ChARF1(N-380)抗原蛋白的純度和濃度已經達到免疫要求.
2.4 多克隆抗體的制備及效價分析
將純化后ChARF1(N-380)抗原蛋白免疫2只雄性日本大耳白兔,獲得E7200和E7201抗血清.對抗血清進行酶聯免疫效價檢測,結果見表2.在不同稀釋濃度下,E7200和E7201血清中抗體與抗原蛋白發生了特異性結合,顯色反應明顯.根據(D陽性-D空白)/(D陰性-D空白)>2.1標準,E7200和E7201抗血清效價皆可達到1∶512 000.

表2 ChARF1(N-380)抗血清ELISA效價檢測
2.5 多克隆抗體的純化及WB檢測
親和純化用蛋白ChARF1(N-380)經檢測濃度為4 mg/mL,與破菌純化后的濃度和純度差異不大,可進行抗原親和純化.親和純化得到抗體E7200和E7201,濃度分別為4.09,4.67 mg/mL.將抗體稀釋(體積比為1∶1 000)后進行WB檢測,抗體E7200和E7201檢測抗原印記在相對分子質量為67 000左右,隨抗原上樣量減少印記變淺,最低可檢測到500 pg抗原(見圖3A,B).抗體E7200和E7201濃度正常,能夠特異性檢測ChARF1(N-380)抗原蛋白.

圖3 ChARF1(N-380)抗體的蛋白質免疫印跡檢測
3 討論
生長素在植物花果發育過程中扮演重要角色.ARFs作為重要的轉錄因子可直接與下游靶基因結合并激活或抑制靶基因在花果發育過程中的表達,從而影響植物的生殖生長[22-23].在本文的研究中,榛子ChARF1基因表達水平從Ov2期開始隨著胚珠的發育逐步上升,在Ov4期達到最大,推測ChARF1基因在胚珠發育過程中具有正調控作用.系統發育分析表明,榛子ChARF1基因與胡桃JrARF基因(XM_035695460.1)高度同源(見圖1B),二者同為可食用干果,種仁含油量較高.其他比對到的同源序列涉及闊葉櫟、歐洲栓皮櫟和扁桃等,則為干果類林木.目前同源性高的匹配序列多為預測參考序列,未見相關的功能研究的報道,通過對榛子ChARF1基因與胚珠發育的研究,將有助于加深對此類植物胚珠發育機制的理解.此外,ChARF1基因與擬南芥ARFs基因家族的AtARF1/2基因、番茄SlARF2基因聚類于進化分枝Ⅰ(見圖1B),AtARF1/2基因影響花的發端和衰老,二者在部分功能上存在冗余,AtARF2基因還具有獨立于乙烯和細胞分裂素外調控花器官脫落功能[6-8];SlARF2基因在番茄中參與花器官的衰老調控和果實發育過程[21].綜上推測,ChARF1基因在胚珠發育過程中發揮重要作用,同時還可能具有AtARF1/2和SlARF2基因的類似功能,參與榛子花的發育和花器官脫落過程.
為進一步驗證ChARF1基因的功能機制,本研究通過構建pET-28a-SUMO-ChARF1(N-380)原核表達載體成功誘導表達ChARF1(N-380)抗原蛋白,首次獲得了能夠特異性檢測榛子ChARF1基因相關蛋白的多克隆抗體E7200和E7201,為后續研究提供了檢測材料.由于本次克隆的ChARF1基因是以cDNA為模板擴增的CDS序列,后續還應以gDNA為模板通過5′-RACE和3′-RACE檢測基因的完整性,在獲得基因完整的序列信息后,進行ChARF1基因序列以及榛子ARF基因家族的系統生物信息學分析和分子功能研究,將有助于深刻揭示ARFs在榛子胚珠發育中所扮演的角色.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
