馬氏珠母貝混養紫海膽對污損生物防除效果的研究
2022-07-28 06:42:54葉孝飛陸慧琳曾靖于宗赫洪喆聰余祥勇王梅芳
水產科技情報 2022年4期
關鍵詞:生物
葉孝飛 陸慧琳 曾靖 于宗赫 洪喆聰 余祥勇 王梅芳
(華南農業大學海洋學院,廣東廣州 510642)
馬氏珠母貝(Pinctadamartensi)又稱合浦珠母貝,是我國重要的海水養殖貝類和珍珠生產的主要母貝。但是,在馬氏珠母貝養殖過程中,其殼表和養殖網籠很快會被污損生物覆蓋。污損生物是附著在船底、浮標和一切人工設施上的動、植物和微生物的總稱。據估計,水產養殖行業每年因為污損生物而造成的直接經濟損失占生產成本的5%~10%[1]。Ventilla[2]研究表明,纏繞盤踞在貝類殼表的污損生物會干擾貝殼的開閉。王春忠等[3]則觀察到,貝殼上附著的污損生物在特定情況下會影響貝類的生長。Wallace等[4]發現,污損生物能與貝類競爭養殖區域的溶解氧。Su等[5]則進一步探討了污損生物附著對馬氏珠母貝攝食、耗氧和排泄物的影響。此外,污損生物的某些種類(如鑿貝才女蟲Polydoraciliata)還能鑿穿貝類的貝殼,造成細菌感染,引發貝類病害[6]。使用化學藥品處理養殖網具如涂抹防污材料三丁基錫(TBT)、有機鋁、有機砷、銅化合物等,以及定期打撈養殖貝籠并用高壓水槍沖洗是目前國內外海上筏式養殖普遍使用的方法[7],但是這些方法費時費力,不僅對貝類損傷較大[8],還會污染養殖海域環境。近些年,利用生物法[9],即通過食物鏈之間的關系選擇污損生物的捕食者進行混養來控制污損生物的方法,因投入費用低、操作簡單、對環境友好等特點而具有很大的發展潛力。
紫海膽(Anthocidariscrassispina)隸屬于棘皮動物門、海膽綱、正形目、長海膽科、紫海膽屬,在我國廣東省沿海海域有廣泛分布[10]。海膽食性雜,其主食藻類,也攝食纖毛蟲、水螅蟲、端足類、橈足類和有機碎屑,且具有很強的運動能力,幾乎能到達養殖網籠全部區域,使污損生物難以附著[11]。利用生物防治法進行污損生物防治已經在實踐中得到應用。Lodeiros等[12]利用紫海膽與牡蠣混養,與對照組相比,網籠污損生物附著量減少了74%,貝殼上的污損生物減少了71%。研究表明,南海相較于渤海、黃海更適于污損生物的生長[13]。但關于在南海用生物防治法的研究,如利用海膽去除貝類養殖過程中的污損生物,以及混養紫海膽對馬氏珠母貝生長的影響等鮮有報道。基于紫海膽是南海海域常見生物,本試驗選用紫海膽與馬氏珠母貝在深圳大亞灣海域進行混養試驗,通過在固定養殖密度的馬氏珠母貝養殖籠中放置不同數量的紫海膽,以馬氏珠母貝和紫海膽的生長指標、存活率和養殖籠上污損生物的質量為測量指標,比較不同混養密度對馬氏珠母貝及紫海膽生長和存活的影響,以及對籠中污損生物的去除效果,以探究大亞灣海域馬氏珠母貝更加合理、高效的養殖模式。
1 材料和方法
1.1 試驗地點和材料
本試驗在大亞灣自然海域(東經114.52°,北緯22.553°)馬氏珠母貝養殖區進行。試驗用養殖籠具為直徑30 cm、高15 cm的6層珠母貝養殖籠,網籠孔目大小為10 mm×10 mm。養殖網籠每層之間用黑色塑料擋板避光,擋板厚3 mm,板上鉆有直徑為5 mm的小圓孔,孔間距10~15 mm。將養殖網籠懸掛在大亞灣內灣水深5 m以上的珠母貝筏式養殖區。
試驗用馬氏珠母貝和紫海膽均取自大亞灣海洋生物綜合實驗站。紫海膽初始殼徑(4.00±0.12)cm,馬氏珠母貝初始殼長(4.00±0.37)cm,均選擇活力旺盛、健康狀況良好的個體。
1.2 試驗方法
設置4個試驗組,即混養組(PA3、PA6、PA9)和對照組(P),每組在養殖馬氏珠母貝30只/層的基礎上,分別混養紫海膽3、6、9、0個/層。另外,設置3個紫海膽單養試驗組(A3、A6、A9),養殖密度分別為3、6、9個/層。以上每個處理組設置5個平行。
馬氏珠母貝和紫海膽的日常管理均參照“大亞灣馬氏珠母貝海上養殖工藝”[14],按常規方法進行。試驗時間為2020年10月—2021年2月。
1.3 數據處理
為了評估單養組與各混養組紫海膽和馬氏珠母貝的生長狀況,在試驗結束時統計各組紫海膽及馬氏珠母貝的存活數量、體質量、殼長,計算各組的存活率、特定生長率、殼長日增長率、肥滿度。采用Ross等[15]的方法評估紫海膽對馬氏珠母貝養殖網籠污損生物的去除效果。試驗前,將養殖網籠完全浸沒在養殖海水中12 h,瀝干水分5 min后稱量,即為干凈網籠質量。將養殖區吊養的試驗網籠撈上岸,用干凈海水沖洗掉淤泥,在日光下晾曬2 h后分別稱取質量,此質量減去干凈網籠質量即為網籠污損生物的質量。計量貝體附著污損生物的方法為:用刀刮取各組的貝體污損生物后,用干凈海水沖洗濾網去除淤泥,然后編號封裝,-4 ℃冷凍,帶回實驗室進行分類鑒定并統計生物量。馬氏珠母貝和紫海膽相關指標的計算公式如下。
存活率Rs(%)=(nt/n0)×100
(1)
特定生長率RSG(%/d)=100×(lnwt-lnw0)/t
(2)
殼長日平均增長率RLD(%/d)=100×(Lt-L0)/(L0·t)
(3)

(4)
式(1)~(3)中,nt為第t天時的存活個數,n0為初始存活個數,wt為第t天時的體質量(g),w0為初始體質量(g),Lt為第t天時的殼長(cm),L0為初始殼長(cm),t為試驗時長(d)。
1.4 數據分析
采用SPSS 19.0軟件對試驗數據進行單因素方差分析(one-way ANOVA),設P<0.05為差異顯著,采用Duncan’s法進行多重比較,用ORIGIN 2018軟件制圖。
2 結果
2.1 馬氏珠母貝和紫海膽的生長情況
各組馬氏珠母貝和紫海膽的生長指標見圖1。結果顯示,各混養組及P組間馬氏珠母貝的存活率沒有顯著差異(P>0.05),混養組紫海膽的存活率均接近100%,各組間無顯著差異(P>0.05)。PA6、PA9組馬氏珠母貝的特定生長率顯著大于P組(P<0.05),而PA3組與P組間差異不顯著(P>0.05)。就馬氏珠母貝的殼長日平均增長率而言,PA3、PA9組與P組間沒有顯著差異(P>0.05),但PA6組顯著大于P組和PA3組(P<0.05)。PA6、PA9組馬氏珠母貝的肥滿度顯著大于P組(P<0.05),PA3與P組間無顯著差異(P>0.05)。紫海膽的特定生長率、殼長日平均增長率、肥滿度在混養與單養的各組間均無顯著性差異(P>0.05)。
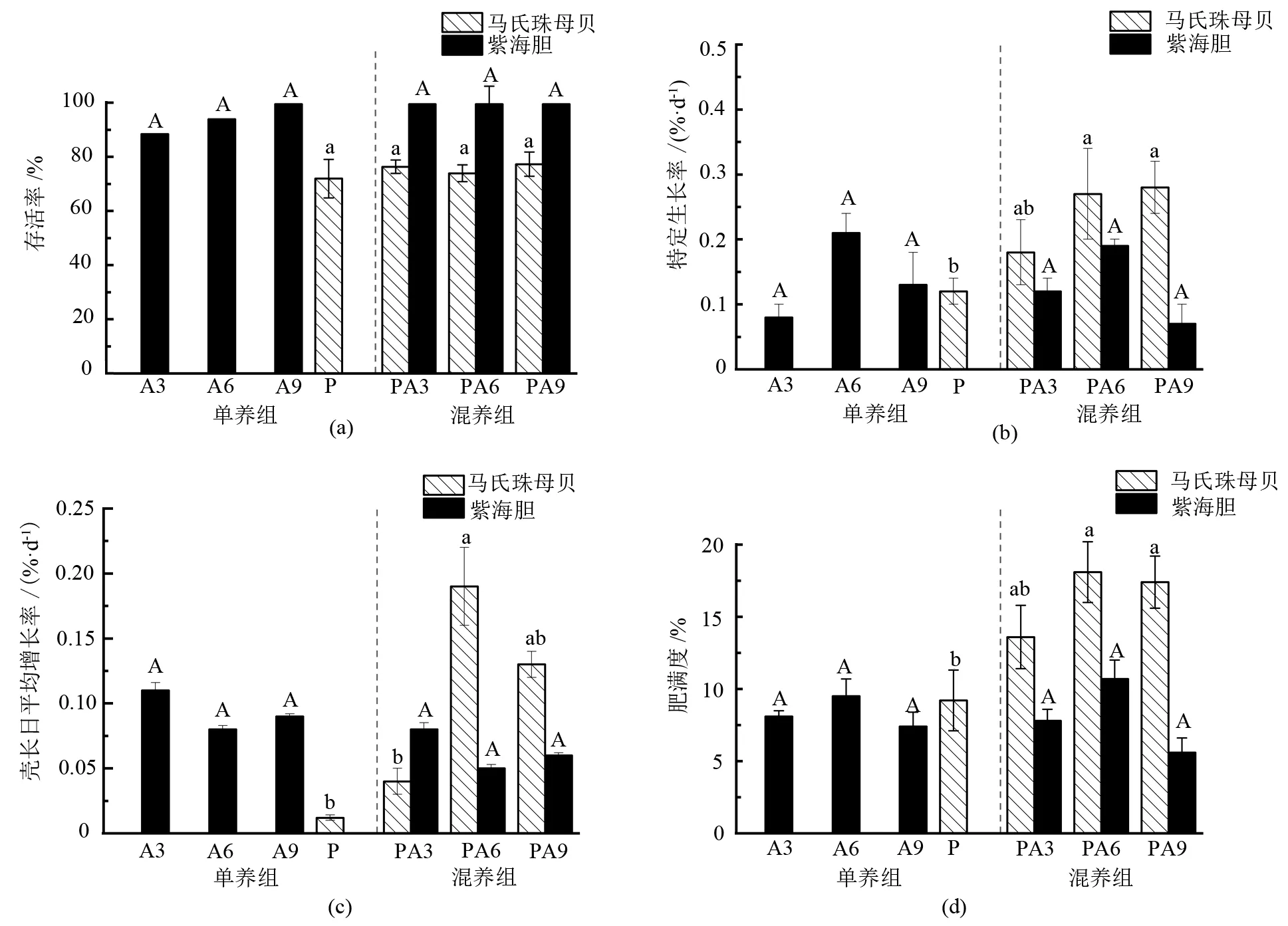
注:柱狀圖上方小寫字母不同表示馬氏珠母貝相關指標組間差異顯著(P<0.05),小寫字母相同表示差異不顯著(P>0.05);柱狀圖上方大寫字母不同表示紫海膽相關指標組間差異顯著(P<0.05),大寫字母相同表示差異不顯著(P>0.05)。
2.2 紫海膽對污損生物的去除效果
由圖2可見,紫海膽各混養組養殖網籠污損生物的覆蓋面積均小于單養網籠。特別是PA6、PA9組,其養殖網籠相比P組而言,污損生物覆蓋面積明顯減少,除了最底部隔板等紫海膽無法到達的區域外,網籠籠衣的清潔度都很高,基本沒有能造成網孔堵塞的污損生物存在。

注:a圖中從左至右依次為P、PA9、PA6、PA3組網籠;b圖中從左至右依次為P、PA3、PA6、PA9組網籠隔板。
由圖3可見,PA3、PA6、PA9組網籠的附著污損生物量均顯著小于P組(P<0.05)。與對照組P相比,混養組PA3、PA6、PA9網籠的污損生物量分別減少了(52.5±12.4)%、(63.3±15.7)%、(70.5±13.1)%。混養組馬氏珠母貝殼表附著的污損生物量分別減少了(45.4±9.6)%、(62.9±7.5)%、(68.1±11.9)%,其中PA6、PA9組殼表附著污損生物量顯著小于P組(P<0.05)。
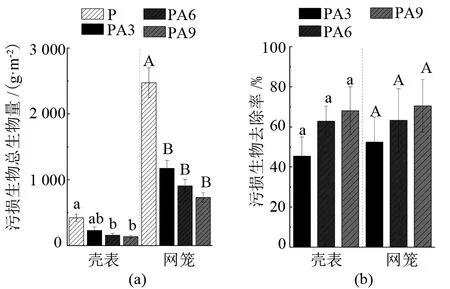
注:小寫和大寫字母分別代表殼表數據組間差異和網籠數據組間差異情況,字母不同表示差異顯著(P<0.05),相同表示差異不顯著(P>0.05)。
2.3 貝體殼表附著污損生物優勢種及紫海膽對污損生物優勢種的去除率
混養組網籠及貝體殼表附著污損生物優勢種的去除率見圖4~圖5。P組網籠附著污損生物優勢種為草苔蟲(Bugulasp.)、密鱗牡蠣(Ostreadenselamellosa)、網紋藤壺(Amphibalanusreticulates)、巴西埃蜚(Ericthoniusbrasiliensis)、粘附山海綿(Mycaleadhaerens)。馬氏珠母貝殼表污損生物優勢種為草苔蟲、密鱗牡蠣、粘附山海綿、瘤海鞘(Styelasp.)、齒緣牡蠣(Dendostreafolium)、河蜾蠃蜚(Corophiumacherusicum)、巴西地鉤蝦(Podocerusbrasiliensis)。從圖4可以看出,PA3、PA6、PA9組紫海膽對P組網籠的污損生物優勢種草苔蟲、巴西埃蜚、粘附山海綿均有很好的去除效果,對巴西埃蜚的去除率達到了100%。但PA3、PA6、PA9組優勢種網紋藤壺的生物量與P組相比分別增加了(15.7±3.2)%、(63.9±7.2)%、(70.2±9.1)%。紫海膽對馬氏珠母貝殼表污損生物優勢種草苔蟲、巴西地鉤蝦、河蜾蠃蜚、瘤海鞘、粘附山海綿的去除率均達到50%以上,但是PA3、PA9組密鱗牡蠣的生物量相較P組分別增加了(8.7±1.2)%、(28.6±2.7)%。
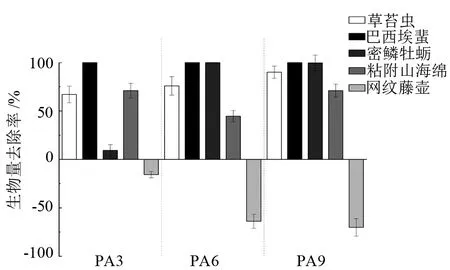
圖4 紫海膽混養組相比于馬氏珠母貝單養組網籠污損生物優勢種生物量去除率
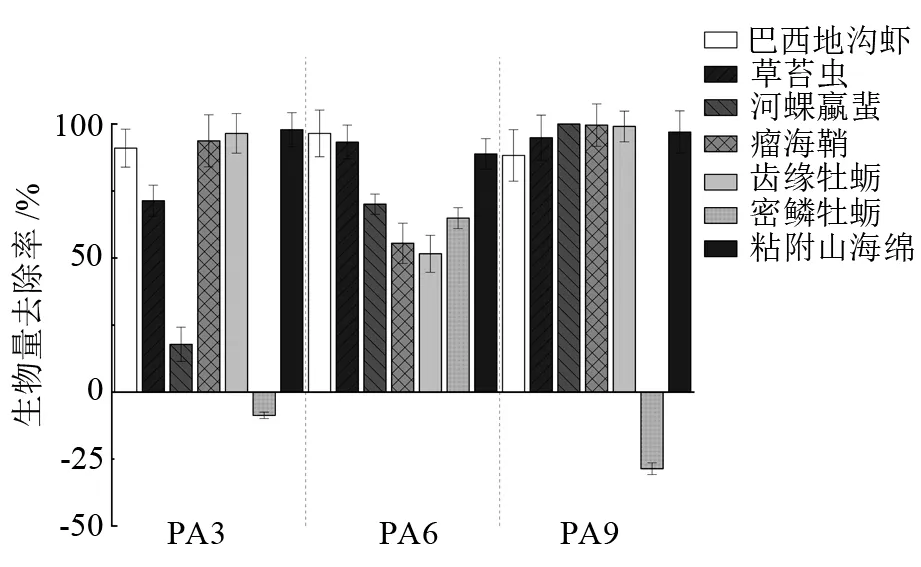
圖5 紫海膽混養組相比于馬氏珠母貝單養組殼表污損生物優勢種生物量去除率
3 討論
3.1 紫海膽不同混養密度對污損生物的去除情況
大亞灣屬于亞熱帶海域,污損生物常年棲息,通常第1個污損生物高峰期在5—8月,10—12月為第2個暴發期[16]。本試驗通過連續4個月的研究觀察,發現在污損生物中,草苔蟲、密鱗牡蠣、網紋藤壺、巴西埃蜚、粘附山海綿為網籠附著優勢種,草苔蟲、密鱗牡蠣、粘附山海綿、瘤海鞘、齒緣牡蠣、河蜾蠃蜚、巴西地鉤蝦為殼表污損生物優勢種。其中草苔蟲為這個時期馬氏珠母貝養殖網籠和貝體附著污損生物絕對優勢種,附著嚴重時甚至可以完全堵塞網眼,阻礙網籠內外的水體交換。鄭東強等[17]對大亞灣海水養殖箱、籠上附著污損生物的研究也發現,草苔蟲是這個時期珍珠貝養殖的重點防除對象。
海膽通常被認為是草食性動物,但隨著人們對其食性的深入研究,發現海膽有著廣泛的攝食選擇。莫寶霖等[18]利用碳氮穩定同位素技術對大亞灣紫海膽進行食性分析,發現其攝食種類包括顆粒有機物(POM)、有機沉積物(SOM)、裂葉馬尾藻、底棲硅藻、浮游動植物等。
本研究將大亞灣自然海域紫海膽與馬氏珠母貝混合養殖,結果表明,與馬氏珠母貝單養對照組(P組)相比,混養組PA3、PA6、PA9網籠附著污損生物的生物量分別減少了(52.5±12.4)%、(63.3±15.7)%、(70.5±13.1)%,而且混養組紫海膽對網籠內馬氏珠母貝殼表污損生物的清除率也達到了(45.4±9.6)%、(62.9±7.5)%、(68.1±11.9)%,PA3、PA6、PA9組網籠及PA6、PA9組貝體污損生物的生物量均顯著小于P組(P<0.05)。這說明利用紫海膽能有效防除污損生物在養殖網籠和貝體的附著,且對污損生物的清除率與紫海膽混養密度呈正相關。但是,PA3、PA6、PA9組網籠上的網紋藤壺及PA3、PA9組貝殼表面密鱗牡蠣的生物量與P組相比反而增加了,推測原因,可能是混養組紫海膽將其他污損生物大量去除,導致網紋藤壺和密鱗牡蠣競爭對手減少,生存空間增大。
3.2 混養試驗組和對照組養殖馬氏珠母貝和紫海膽的生長特性
本試驗中,各混養組及P組間馬氏珠母貝的存活率無顯著差異(P>0.05),而紫海膽的存活率均接近100%,組間無顯著差異(P>0.05),表明混養紫海膽并不會對馬氏珠母貝和紫海膽的存活造成影響。PA6、PA9組馬氏珠母貝的特定生長率、肥滿度均顯著高于P組(P<0.05),PA6組的殼長日平均增長率也顯著高于P組(P<0.05),究其原因,是混養紫海膽后,網籠附著污損生物特別是絕對優勢種草苔蟲被大量去除,使得通過網籠的水流加大,增加了馬氏珠母貝的食物供應(即浮游生物、溶解氧、有機碎屑的含量更高)[19-21],促進了馬氏珠母貝的生長。尤其當紫海膽混養密度為6、9個/層時,能顯著影響馬氏珠母貝的生長(P<0.05)。紫海膽的特定生長率、殼長日平均增長率和肥滿度在混養與單養的各組間均無顯著性差異(P>0.05),表明混養不會對紫海膽的生長造成影響。綜合污損生物去除率以及馬氏珠母貝和紫海膽的生長情況,建議紫海膽的混養密度以6個/層為宜。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
