魚類對飼料可消化糖的利用和代謝特征
2022-07-28 06:43:02程開敏李沛洋鄭石軒朱學芝馬學坤李新宇
水產科技情報 2022年4期
程開敏 李沛洋 鄭石軒 朱學芝 馬學坤 李新宇
(廣東粵海飼料集團股份有限公司,廣東湛江 524017)
自20世紀80年代以來,國際水產養殖總量以每年6%~12%的速率增長,目前已經提供超過一半的全球漁業產量[1]。根據相關統計數據,全球接近70%的水產養殖依賴于投喂商業飼料,因此水產動物營養與飼料科技的進步對水產養殖健康發展至關重要[2]。為了達到較高的產量和利潤,主流商業配合飼料往往呈現高蛋白高能量的特征。近年來,膨化飼料因具有較好的物理性狀和可消化率,已成為許多水產飼料的主流產品[3]。為達到理想的生產效率和膨化效果,目前膨化飼料的淀粉推薦水平約為20%[4]。
糖類是魚類三大營養物質之一[5-6]。糖類作為非蛋白能量物質,有節省蛋白質的作用,并且在代謝過程中能夠產生一定的有益代謝產物,飼料中保持適量的可消化糖(主要是淀粉)水平能夠提高養殖對象的生長速度和飼料利用表現[7-8]。為了節省成本和減少蛋白質的浪費,商業飼料往往盡可能地提高非蛋白能(如可消化糖和脂肪)的使用比例。另一方面,不同食性的魚類對飼料淀粉的利用率差異較大,草食性或者部分雜食性魚類可以充分利用40%淀粉水平的飼料而不影響生長和健康[4,9],而肉食性魚類飼料的淀粉水平一般在20%以下,個別品種甚至要求低于10%[3]。有研究表明,過量攝入糖類將影響魚類生長、攝食和健康[10]。雖然近年來配合飼料技術發展迅速,但給魚類投喂這些高能量高淀粉的商業配合飼料往往會導致一定的健康問題。因此,針對不同品種的魚類,應該謹慎評估飼料的淀粉水平,以保證魚類健康快速生長。
近年來,國外已有學者就魚類對飼料可消化糖類的消化、吸收和利用進行了總結[11]。本研究在過去研究的基礎上,總結了不同魚類對糖類的代謝特征和常見的研究方法,并歸納了不同食性魚類對飼料淀粉的利用效果,皆在為商業配合飼料開發提供理論指導依據。值得注意的是,廣義糖類物質涵蓋范圍較廣,本研究主要總結的是可消化糖類(如淀粉、葡萄糖等)在魚類營養中的研究進展。
1 糖類
蛋白質、脂肪和糖類是魚類飼料中的三大營養物質。糖類作為重要的能量來源,不僅價格低廉,還具有節省蛋白質、保護環境等作用。糖類的準確定義為:多羥基醛或多羥基酮以及水解后能夠產生多羥基醛或多羥基酮的一類有機化合物[12]。糖類是自然界中分布極為廣泛的一類有機化合物,大多數植物體糖含量可達干質量的80%。植物種子中的淀粉,根、莖、葉的纖維素,動物組織中的糖原、黏多糖,以及蜂蜜和水果中的葡萄糖、果糖等都是糖類。其中,淀粉(如小麥淀粉、木薯淀粉、玉米淀粉等)是魚類配合飼料的主要可消化糖來源。
2 淀粉的消化和吸收
飼料淀粉的消化和吸收(葡萄糖形式)是動物利用可消化糖作為能量源的初始步驟。在腸道淀粉酶的作用下,多糖(如淀粉和糖原)被分解為單糖(葡萄糖)被腸道吸收。在哺乳動物中,淀粉的消化從口腔開始,在唾液淀粉酶的作用下被消化成麥芽糖或糊精[13]。但是,由于魚類口腔中缺乏淀粉酶,該步驟在魚類中并不重要。糊精和麥芽糖可被麥芽糖酶進一步水解成葡萄糖。在魚類體內,淀粉酶主要由胰腺產生,并分泌到腸道和幽門盲囊中。例如,在黃尾魚師(Seriolalalandi)的幽門盲囊和腸道中可檢測到較高活性的α-淀粉酶[14]。值得注意的是,α-淀粉酶只可以水解淀粉內部的α-1,4-糖苷鍵,而不能水解α-1,6-糖苷鍵。大量研究已經證明,淀粉酶活性受魚類食性的影響,草食性魚類的淀粉酶活性往往高于肉食性魚類,如鯉魚和金魚的消化道淀粉酶活性明顯高于海鯛、鰻魚和鱒魚的相應酶活性[15]。同時,淀粉酶活性也受多種其他因素影響,如營養狀態、溫度和飼料成分等[3]。此外,葡萄糖作為一個極性物質不能直接穿透細胞膜,因此需要轉運蛋白協助吸收。
魚類對飼料淀粉的消化還受到不同淀粉源及其水平的影響,并且整體低于蛋白質和脂肪的消化率[3]。淀粉顆粒越小越有利于其消化。因此,糊化淀粉一般比生淀粉擁有更佳的消化率[3,16]。另外,淀粉原料中的直鏈淀粉與支鏈淀粉比例也會影響對淀粉的消化。一般認為,支鏈淀粉更容易被消化[3]。通常而言,糖類鏈越短,越有利于被魚類消化吸收(葡萄糖>麥芽糖>淀粉),但不同品種略有差異。如團頭魴(Megalobramaamblycephala)對葡萄糖、麥芽糖和糊精擁有較高的消化率,其次為小麥淀粉和玉米淀粉,最差為纖維素[17]。由于魚類淀粉酶分泌有限,往往會有一個最高的飼料淀粉可消化水平[18],如隨著飼料淀粉水平從8%上升至17%,大西洋比目魚對淀粉的消化率從84%降至53%[19]。對草魚的研究也表明,其對淀粉的消化率隨著飼料淀粉水平的上升而下降[20]。
3 葡萄糖轉運
葡萄糖轉運體存在于身體的各個組織細胞中,一般分為兩類。一類是鈉依賴的葡萄糖轉運體(SGLT),以主動方式逆濃度梯度轉運葡萄糖;另一類是易化擴散的葡萄糖轉運體(GLUT),以易化擴散的方式順濃度梯度轉運葡萄糖,其轉運過程不消耗能量。與哺乳動物一樣,糖在魚體消化道被分解成單糖后,通過腸道黏膜SGLT1被組織吸收。目前,對魚類的不同葡萄糖轉運體有一定的研究。GLUT1和GLUT3是一類廣泛分布于眾多組織器官中的葡萄糖轉運體,其中個別魚種的GLUT3在腦部表達較高[21]。GLUT2主要在肝臟、腸道和腎臟中表達較高[18,22-23]。GLUT4則主要在肌肉組織中表達,包括白肌、紅肌和心肌[18,24],考慮到肌肉組織占比較大,該轉運體對魚體內糖穩態調節起到重要作用。GLUT4轉運活性受限或者對相關激素調節不敏感是魚類高糖不耐癥的主要原因之一[25-26]。部分常見魚類的GLUTs轉運體在體內的分布情況見表1[24]。

表1 GLUTs在不同魚類體內的組織分布
4 葡萄糖的代謝
血液流經各組織時,葡萄糖被各組織吸收,一部分被直接氧化利用,另一部分被轉變為糖原或脂肪。整體上,體內葡萄糖的去路可分為3條路徑(見圖1):①糖酵解(glycolysis)產生丙酮酸進入三羧酸循環徹底氧化,或進一步合成脂肪和乳酸;②戊糖磷酸途徑(pentose phosphate pathway)代謝產生還原型輔酶Ⅱ(NADPH)和核糖-5-磷酸;③合成糖原,儲存在肌肉和肝臟組織中。此外,動物體能將簡單的非糖前體(如乳酸、甘油、生糖氨基酸等)轉變為糖(葡萄糖或糖原),又稱為糖異生。
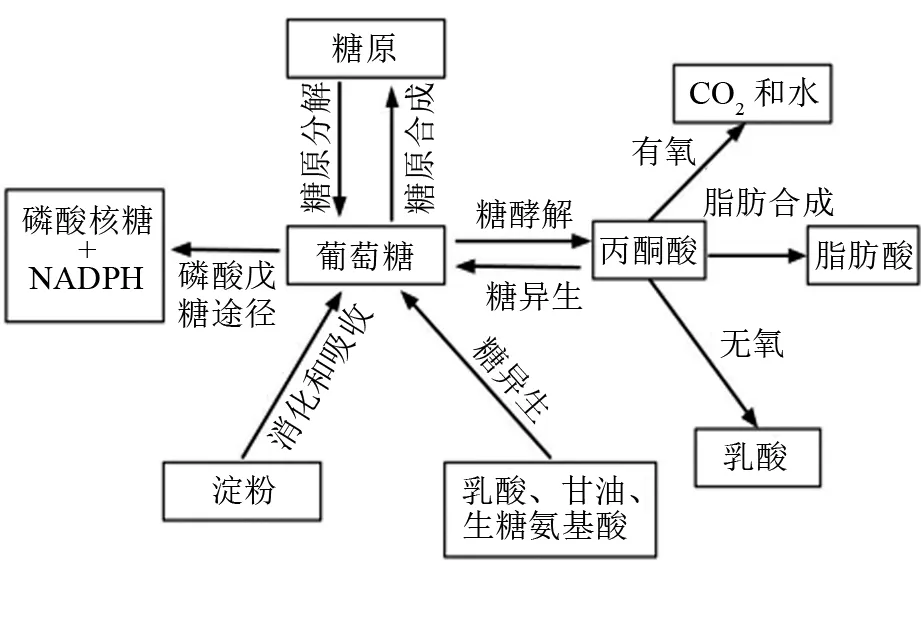
圖1 動物體內糖類消化、吸收和代謝的基本途徑
4.1 糖的降解和異生
糖酵解和糖異生是維持機體糖穩態的兩條重要代謝通路。糖酵解是從葡萄糖開始分解生成丙酮酸的過程,全過程共有10步酶催化反應。該通路主要通過3個關鍵酶反應步驟進行調控:①己糖激酶(HK)參與的葡萄糖磷酸化,需要Mg2+作為輔助因子;②磷酸果糖激酶-1(PFK-1)參與的催化6-磷酸果糖生成1,6-二磷酸果糖;③丙酮酸激酶(PK)參與的催化磷酸烯醇式丙酮酸分子高能磷酸基團轉移給ADP生成ATP,產生丙酮酸,需要Mg2+和K+參與,反應不可逆。己糖激酶目前已知有4種不同的型態,包括己糖激酶Ⅰ、己糖激酶Ⅱ、己糖激酶Ⅲ,以及與前三者差異較大的己糖激酶Ⅳ(又稱為葡萄糖激酶,GK)。其中,GK擁有較高Km值,對血糖調節意義重大,其表達往往受血糖水平的影響[27]。糖酵解產生的丙酮酸可進一步被分解成乙酰輔酶A,然后進入三羧酸循環,產生更多的ATP。另一方面,在乳酸脫氫酶的作用下,丙酮酸也可能被無氧酵解為乳酸。在魚類中,組織(尤其白肌)往往含有較高的乳酸脫氫酶活性[24],證明該途徑也是魚類葡萄糖代謝的主要路徑之一。草食性魚類能夠通過提高糖酵解酶活性來適應飼料中較高的淀粉水平[28],但肉食性魚類相關酶活性的調節能力有限,導致對飼料淀粉利用受限[29-31],如大黃魚投喂過量的飼料淀粉反而會降低GK和PK的活性[32]。研究也表明,肉食性魚類各個組織主要以氨基酸為能量來源,而非葡萄糖[33-34]。
在哺乳動物中,還存在一個乳酸循環(lactic acid cycle)。當肌肉收縮時(尤其是供氧不足時)通過糖酵解生成乳酸。肌肉內糖異生活性較低,所以乳酸通過細胞膜彌散進入血液后,再被肝臟組織攝入,在肝內異生為葡萄糖。然后,該葡萄糖釋入血液后又可重新被肌肉利用,這就構成了一個循環(肌肉-肝臟-肌肉),此循環也稱Cori循環。前文提到,大部分魚類白肌能夠產生大量乳酸,因此魚類也可能存在這種循環,但相關研究仍然十分有限。
糖異生的七步反應是糖酵解中的逆反應,它們有相同的催化酶。另一方面,糖酵解中有三步不可逆反應,因此糖異生時必須繞過這三步反應。整體上,糖異生主要由以下4種酶控制反應:①丙酮酸羧化酶,可以催化二氧化碳固定在丙酮酸上,生成草酰乙酸;②磷酸烯醇丙酮酸羧激酶(PEPCK),催化草酰乙酸形成磷酸烯醇式丙酮酸和二氧化碳;③果糖-1,6-二磷酸酶(FBPase),催化1,6-二磷酸果糖和水生成6-磷酸果糖;④葡萄糖-6-磷酸酶,催化葡萄糖合成。有研究報道,體內糖異生是維持葡萄糖穩態的重要路徑[35]。如在攝入足量淀粉的情況下,鯉魚可通過減少糖異生活性,預防血糖過高[36]。但對于某些肉食性魚類,它們缺乏這種調控機制,從而導致無法適應高糖飼料[9]。也有研究者發現,虹鱒糖異生活性不受飼料糖水平的影響[37-39]。
磷酸戊糖途徑是葡萄糖氧化分解的另一種方式,具有多種生理意義:①為核酸的合成提供核糖;②生成大量的NADPH+H+,作為供氫體參與多種代謝反應;③通過磷酸戊糖途徑中的轉酮醇基及轉醛醇基反應,促進糖在體內的轉變;④NADPH可用作脂肪酸合成,并參與抗氧化反應。目前,該通路在魚類中的研究相對較少。
4.2 糖原和脂肪合成
游離葡萄糖在糖原合酶的催化下與引物分子(未降解完全的糖原分子或糖原素)合成糖原。值得注意的是,游離的葡萄糖分子并不能直接作為糖原合酶的底物,需要在細胞內首先被激活為6-磷酸葡糖,再經磷酸葡糖變位酶催化異構化為1-磷酸葡糖。糖原合成能夠暫時儲存體內過多的葡萄糖,對維持血糖穩定意義重大。最近的研究表明,飼料可消化糖水平增加會引起部分魚類肝臟糖原快速積累,但長期過度積累將導致肝臟損傷[40-42]。
如前文所述,糖酵解后產生丙酮酸可被進一步分解為乙酰輔酶A,而乙酰輔酶A是合成脂肪酸的重要物質。乙酰輔酶A與碳酸氫根、ATP反應,羧化生成丙二酸單酰輔酶A。該反應由乙酰輔酶A羧化酶(ACC)催化,是脂肪酸合成的限速步驟。乙酰輔酶A和丙二酸單酰輔酶A在脂肪合成酶的作用下,通過連續的酮還原酶、脫水酶以及烯脂酰ACP還原酶的作用,完成脂肪鏈的延長。磷酸戊糖途徑產生的NADPH也參與了該反應,因此葡萄糖代謝對脂肪合成非常重要。對部分魚類的研究結果也證明,高淀粉飼料提高了體內脂肪蓄積或組織脂肪合成相關基因的表達水平,證明魚類可以通過調節脂肪合成適應高糖飼料[43-44]。
5 血糖和激素調節
盡管大部分魚類的空腹(24 h以上)基礎血糖水平差異不大,血糖濃度大多在3~7 mM(見表2)[28,43-44,52-71],但餐后或葡萄糖注射后的峰值水平差異較大,在5~40 mM[9]。血糖必須保持一定的水平才能維持體內各器官和組織的正常工作。胰島素和胰高血糖素是調節血糖穩定的主要兩種激素。胰島素是降低血糖的激素,能夠促進糖原、脂肪、蛋白質合成,并促進糖酵解。胰高血糖素具有很強的促進糖原分解和糖異生作用,能使血糖明顯升高。魚類血液的胰島素含量大多在0.2~5.0 nmol/L,但個別餐后可能更高。Kamalam等[11]報道,胰島素可刺激魚類組織提高葡萄糖的攝入效率,通過激活相關通路,刺激糖酵解、糖原和脂肪合成,并抑制糖異生,從而達到血糖下降的目的。對于部分魚類,血清胰島素水平明顯隨飼料糖水平的提高而提高[45]。但對虹鱒的研究表明,胰島素反而會抑制血糖清除速率[從(8.6±0.6)μmol·kg-1·min-1下降到(5.4±0.5)μmol·kg-1·min-1],從而證明某些肉食性魚類胰島素的作用可能與傳統認知不同[46]。Navarro等[47]研究表明,魚類胰島素對血糖刺激不敏感,尤其是肉食性魚類,胰島素主要受游離氨基酸刺激而分泌。對虹鱒的研究表明,精氨酸注射能使血清胰島素水平從12.5 ng/mL上升到76.8 ng/mL,并增加胰島素和組織的結合效率[48]。對加州鱸的研究表明,精氨酸能夠刺激胰島素的分泌,并且效果稍好于葡萄糖的刺激效果[49]。胰高血糖素在魚類中的作用基本與其在哺乳動物中的作用相似。對幾種魚類的研究表明,胰高血糖素能夠提高糖原分解和糖異生的效率[9]。在對虹鱒的研究中發現,該激素通常在餐后或高血糖情況下被抑制[50]。此外,該激素分泌也受飼料氨基酸組成和環境因素的影響[51]。
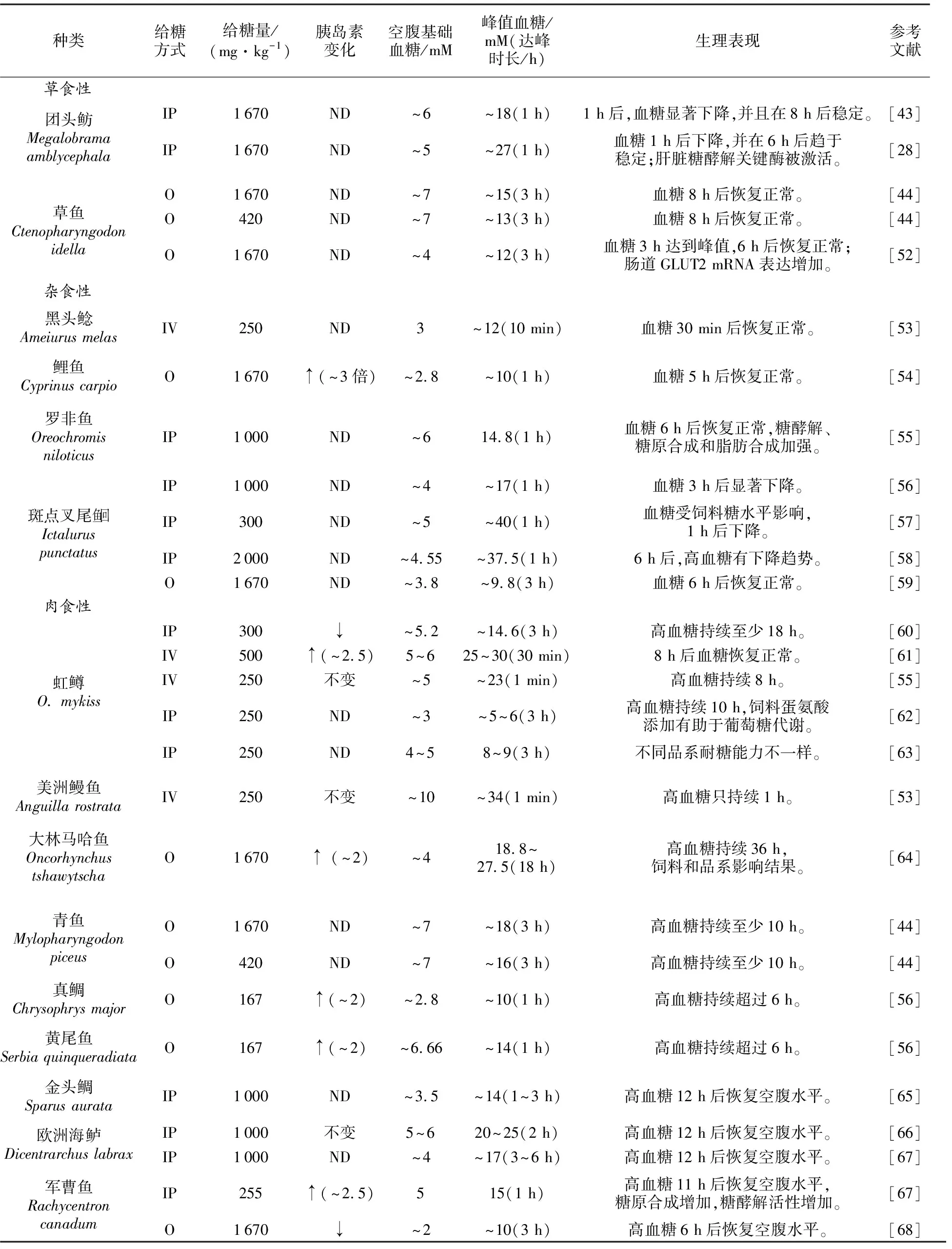
表2 部分魚類糖耐量試驗結果
6 試驗方法
6.1 糖耐試驗
糖耐試驗,也稱葡萄糖耐量試驗,是檢測動物對葡萄糖利用的一種實驗室方法,主要有注射和口服兩種。在人類疾病診斷中,該試驗有較好的統一標準,并且對患者的藥物服用、飲食攝入、生理活動等都做了一定的約束。糖耐量最早用于診斷人類的糖尿病,其測定方法是在空腹狀態下口服一定劑量(一般為1 670 mg/kg體質量)葡萄糖后,每隔一段時間測定血糖水平,對比血糖恢復到空腹的基礎水平所需要的時間來確診糖尿病。在魚類研究中,糖耐量試驗的給糖方式主要包括口服、靜脈注射、腹腔注射和肌肉注射[72]。不同的方式具有各自優劣性,口服方式對試驗動物的刺激小,能比較準確地反映魚類對葡萄糖的耐受能力。注射方式敏感性高、重復性好、變異性小,但注射易使試驗動物產生一定的應激而影響最后的血糖數值。不同于人類,魚類糖耐量試驗中的給糖量差異較大,為200~2 000 mg/kg不等。從表2可知,不同的給糖量顯著影響糖耐量試驗中的血糖濃度和高血糖持續時間,從而影響不同試驗之間的相互比較與參考。在此建議,今后應確定一套規范的方法,規定糖耐量試驗中的口服以及注射的葡萄糖量,并要詳細規定對象魚類的生理狀態條件,以便比較不同種魚類的糖利用率。
不同食性的魚類對葡萄糖的耐受能力不同,一般來說,肉食性魚類比雜食性和草食性魚類對葡萄糖的耐受能力要低。有研究給青魚和草魚分別飼喂等劑量(1 670 mg/kg或420 mg/kg)葡萄糖后,兩種魚的血糖均有不同程度的升高,但相同時間內草魚的血糖含量比青魚低,血糖恢復到空腹水平的時間也比青魚短[73]。另一個試驗中,被注射等量葡萄糖(300 mg/kg)后,羅非魚的葡萄糖耐受能力最強,卵形鯧鲹次之,軍曹魚最弱[74]。此外,魚的營養狀態也可能影響最后的試驗效果。用糊精含量分別為5%、25%、50%的3種飼料飼養異育銀鯽,禁食24 h后,再口服1 670 mg/kg的葡萄糖,各組魚血糖迅速升高,均在3 h后出現血糖峰值,但5%糊精組和25%糊精組在5 h后血糖即恢復到空腹水平,50%糊精組7 h后才恢復到空腹水平[75]。對羅非魚(Oreochromisniloticus)的研究結果也表明,給養殖魚類投喂不同的飼料糖水平,以及給糖量不同都將影響最后的血糖數值,詳見表2。
6.2 投喂試驗
此外,可以通過配制含有不同淀粉水平的飼料,投喂養殖魚類數周,觀察其生長、飼料利用效率、血糖以及其他指標來評價魚類對糖的利用能力。試驗飼料應符合如下要求:①除被試驗的成分(淀粉)外,試驗飼料的所有其他組分盡可能完全一致,且營養全面;②試驗飼料的化學物理性狀應符合試驗動物的攝食習性要求;③具有良好的水中穩定性,以減少營養物質的溶失,提高營養物質攝入量的估計精度。
由于魚類不能利用大量糖作為能量(糖酵解受限),因此過高的淀粉攝入往往以糖原和脂肪形式儲存在體內。但肝臟是主要代謝器官,過量的糖原和脂肪積累將導致肝臟功能減退。因此,探明不同魚類的肝臟對高糖飼料的反應特征具有十分重要的意義。幾乎所有魚類的肝臟體積都會隨著飼料淀粉水平的提高增大,但形式有所不同。例如,對草魚的研究表明,其肝臟質量和脂肪水平隨著飼料糖水平提高而增加,但糖原含量變化不明顯[76]。還有一些魚類,如虹鱒的脂肪和糖原都能夠在肝臟中有效積累。但很多肉食性魚類傾向于將過量葡萄糖合成糖原而非脂肪,且糖原主要儲存于肝臟中。對石斑魚的研究表明,肝臟體積和肝糖原含量隨著飼料淀粉水平提高而增加,且肝臟脂肪呈現下降趨勢。對加州鱸的研究也表明,其肝臟糖原的合成速率遠高于脂肪的合成速率[40]。研究發現,肉食性魚類的肝臟往往含有較高的糖原,如歐洲海鱸肝臟糖原含量為105 mg/kg[27],金鯧魚(Trachinotusovatus)為287 mg/kg[77],石斑魚(Epinephelusakaara)為227 mg/kg[78]。由此可見,肝臟糖原過度積累是許多養殖肉食性魚類后期生長受限和健康問題頻發的主要原因。值得注意的是,不同魚類的肝臟成分差異巨大,這導致很難形成一個專門的診斷標準來確定魚類肝臟代謝疾病。因此,未來應根據不同魚類自身的生理特點來建立不同標準診斷依據。此外,不同養殖試驗的血糖結果差異較大,這是因為大部分試驗主要以空腹血糖(饑餓24 h后)為指標。因此建議,養殖試驗和糖耐試驗應相互配合佐證,以得到更明確的試驗結果。
7 結論和展望
飼料可消化糖對魚類生長和代謝影響的研究已較全面,但在許多其他營養物質的研究中,卻一直被當作能量填充物,其對魚類生理和代謝的影響一直被低估。如在對一種肉食性魚類的蛋白質需求量定量試驗中,研究者配制了7組等能但不等氮的飼料,飼料蛋白質水平由25%上升至55%,同時降低飼料淀粉水平,由55%降至25%[79]。盡管該試驗未評價健康指標,但低蛋白組魚的攝食量只有中蛋白組的60%左右。類似的研究也發現,幼魚的肝臟大小、肝糖原含量和攝食量受飼料淀粉水平影響較大[80]。因此,很多試驗可能受淀粉水平的干擾,導致結果誤差較大。在未來的研究和生產中,應該更加注重淀粉水平的合理設置。此外,由于水產養殖對象的多樣性,不同種類的葡萄糖代謝模式存在較大差異,如肉食性魚類往往傾向于將過量葡萄糖以糖原形式儲存在肝臟中,而有些魚類則既能合成糖原也能合成脂肪,在研究時應根據種類制定相應的研究方法。此外,雖然糖耐量試驗是了解魚類對糖類利用的重要研究手段,但目前還沒有統一的試驗標準,導致結果參考意義不強。建議將來在統一方法標準的基礎上,重點研究不同種類魚類的葡萄糖代謝特征及調控方法。
猜你喜歡
保健醫苑(2022年6期)2022-07-08 01:26:34
家庭科學·新健康(2022年3期)2022-05-10 00:32:13
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
家庭醫學(下半月)(2020年1期)2020-05-11 02:05:44
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
媽媽寶寶(2017年3期)2017-02-21 01:22:30
飼料與畜牧(規模養豬)(2016年5期)2016-12-01 03:48:40
人人健康(2016年13期)2016-07-22 10:34:06
學苑創造·A版(2015年11期)2016-01-14 09:03:27
