馬鈴薯長期連作對土壤真菌群落特征的影響
2022-07-28 03:19:30胡新元譚雪蓮孫小花謝奎忠柳永強
干旱地區農業研究 2022年4期
胡新元,譚雪蓮,孫小花,謝奎忠,柳永強
(1.甘肅省農業科學院,甘肅省馬鈴薯種質資源創新工程實驗室,甘肅 蘭州 730070; 2.甘肅省農業科學院旱地農業研究所,甘肅省水資源高效利用重點實驗室,甘肅 蘭州 730070; 3.甘肅省農業科學院馬鈴薯研究所,甘肅 蘭州 730070)
馬鈴薯(SolanumtuberosumL.)是世界第四大糧食作物,我國是馬鈴薯種植面積最大的國家,約占世界種植面積的22%[1-2]。由于人口不斷增長,預計到2050年全球糧食需求將翻一番[3-4]。中國作為世界第二大經濟體,一直在采取措施提高糧食產量,2015年中國農業部啟動馬鈴薯主糧化戰略,到2020年,馬鈴薯面積由5.3×106hm2增加到1.0×107hm2以上[5]。近年來,馬鈴薯連年種植現象非常普遍,連作會造成環境因子的過度消耗,使土壤微生態環境惡化,植物病原體富集,最終導致作物生長障礙[6]。連作導致馬鈴薯土傳病害逐年加劇,嚴重影響了馬鈴薯產業的健康發展。據報道,馬鈴薯連作2 a,產量沒有顯著變化,但連作第3年,產量下降17.07%以上[7-8]。有研究表明,微生態系統的失衡是造成連作障礙發生的主要原因之一,土壤微生物類群的變化又會影響土壤微生態功能[6]。馬鈴薯連作導致的土壤微生物群落結構演替一直以來備受關注。因此,研究不同連作年限下土壤微生物多樣性的變化,對于促進馬鈴薯種植業的可持續發展具有重要意義。
土壤真菌與土傳病害的發生密切相關[9]。孟品品等[10]采用PCR-DGGE分子指紋圖譜技術研究了馬鈴薯連作對根際土壤真菌種群結構的影響,研究表明連作使根際土壤中病原真菌成為優勢種群,根際微生態環境惡化。李繼平等[11]采用稀釋平板法和殘渣分離法研究了馬鈴薯連作對土壤病原真菌種群動態變化的影響,發現0~10 cm土層中土傳病原真菌菌落數隨連作年限的增加而增加。龔靜等[12]研究表明,馬鈴薯連作1季和3季土壤真菌結構相似。研究發現,大豆長期連作,真菌群落結構和功能組成向利于植物健康方向發展[13]。大豆長期連作(6~38 a),土壤中抑制大豆胞囊線蟲的假單胞菌屬(Psudomonas)、紫霉屬(Purpureocillium)和pochonia豐度逐年增加[14]。與連作3 a相比,大豆連作20 a根際土壤中根腐病病原菌鐮孢菌(Fusariumoxysporum)種群密度顯著降低[15]。長期種植馬鈴薯是否有利于抑病土的產生,從而抑制病原菌的生長,改善土壤環境和植株生長,馬鈴薯是否可以進行適度連作,目前仍罕見報道。因此,本研究采用高通量測序技術,通過研究馬鈴薯土壤真菌多樣性和群落結構在連作條件下的演變規律,以期為解決馬鈴薯適度連作和土壤微生態修復問題提供理論依據。
1 材料與方法
1.1 研究區概況和試驗設計
試驗布設在甘肅省定西市安定區寧遠鎮紅土村中川社(35°26.714′N, 104°50.886′E),海拔1 991 m,試驗地為平坦川地,屬中溫帶偏旱區,日照時數2 476.6 h,年均氣溫6.4℃,≥10℃積溫2 239.1℃,無霜期140 d。多年平均降水量390.9 mm,年蒸發量1 531 mm,干燥度2.53,為典型的黃土高原半干旱雨養農業區,土壤為典型的黃綿土。
2010年3月開始進行定位試驗,供試品種為:春小麥(定西35號)、豌豆(隴豌1號)、馬鈴薯(隴薯10號)。試驗采用隨機區組設計,設6個處理。(1)輪作(R):馬鈴薯/小麥/豌豆序列輪作,2019年種植馬鈴薯;(2)馬鈴薯連作1 a(C1):2018—2019 年連續2 a種植馬鈴薯;(3)馬鈴薯連作3 a(C3):2016—2019 年連續4 a種植馬鈴薯;(4)馬鈴薯連作5 a(C5):2014—2019 年連續6 a種植馬鈴薯;(5)馬鈴薯連作7 a(C7):2012—2019年連續8 a種植馬鈴薯;(6)馬鈴薯連作9 a(C9):2010—2019年連續10 a種植馬鈴薯。C5~C9處理的前茬作物均為春小麥。每個處理3次重復,共18個小區,每個小區面積為23.8 m2(5.67 m×4.2 m)。
1.2 播種、施肥量及田間管理
小麥播種量為187.5 kg·hm-2, 3月下旬播種,7月下旬收獲。豌豆播種量為180 kg·hm-2,4月初播種,7月中旬收獲。馬鈴薯種植密度為67 500株·hm-2,4月下旬種植,9月下旬收獲。小麥施肥量為純N和P2O5各105 kg·hm-2,豌豆施肥量為純N 135.0 kg·hm-2、P2O5108.0 kg·hm-2,馬鈴薯施肥量為純N135 kg·hm-2、P2O5108 kg·hm-2。在作物生長季節,雜草采用人工除草,此外無其他田間管理措施。
1.3 取樣
馬鈴薯塊莖膨大期采用 “S”形方法取樣,從0~20 cm土層采集距離馬鈴薯根部周圍0~2 cm土樣,樣品混勻后過2 mm篩,裝入無菌塑料離心管中,放入液氮中帶回實驗室,并儲存于-80℃冰箱用于微生物分析。
1.4 土壤微生物高通量測序
使用MN NucleoSpin soil Kit (Machery-nagel, Dueren, Germany)從0.75 g土壤中提取總DNA。根據真菌ITS1保守區使用引物ITS1F (5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2 (5′ -GCTGCGTTCTTCATCGATGC-3′)。每10 μl PCR混合物中含有5 μl KOD FX Neo Buffer、0.3 μl 10 μM引物、2 μl dNTP(每個2 mmol·L-1)、0.2 μl KOD FX Neo、50 ng DNA,并加入蒸餾水定容至10 μl。每個樣品進行重復擴增、合并,然后使用Cycle Pure Kit (Omega, Norcross, GA, USA)進行純化,形成測序文庫,使用Illumina HiSeq 2500技術進行測序分析。
1.5 數據質控和數據處理
每個種植方式對應選取3個獨立土壤樣品進行ITS測序,鑒定樣品中的真菌;測序序列的Q3控制90%以上;從樣本中隨機抽取一定數量的序列,統計這些序列所代表的物種數目,并以序列數與物種數來構建曲線,控制序列數取值在曲線平緩段,保證測序數據量飽和,足以反映樣品中的物種多樣性。
用QIIME軟件挑選出屬分類學水平上豐度最高的OTU序列作為代表序列,進行多重序列比對并構建系統進化樹,然后通過Python語言工具繪制圖形。通過Beta多樣性分析來比較不同樣品在物種多樣性方面(群落組成及結構)存在的差異大小,根據距離矩陣獲得相應距離下的樣品層次聚類樹,并進行非度量多維標定(Non-metric multi-dimensional scaling, NMDS)分析。
2 結果與分析
2.1 土壤豐富度和多樣性比較
采用高通量測序技術對馬鈴薯土壤真菌群落的α-多樣性指數進行了分析。馬鈴薯連作年限對土壤真菌群落豐富度有顯著影響,相似性水平為97%(圖1)。隨著馬鈴薯連作年限的增加,土壤真菌菌群豐度和Chao1顯著增加。連作7 a和9 a的馬鈴薯土壤真菌菌群豐度最高,馬鈴薯/春小麥/碗豆輪作的馬鈴薯土壤真菌菌群豐度最低。C7與C9間無顯著差異,而C7、C9與R、C1 C3間差異顯著。表明連作年限的增加使土壤真菌群落的豐富度和多樣性提高。
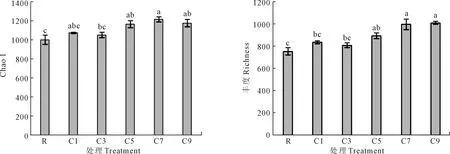
注:不同字母表示處理間差異顯著(P<0.05)。Note:Different letters indicate significant differences among different treatments (P<0.05).圖1 馬鈴薯連作年限對土壤真菌群落Chao1和菌群豐度的影響Fig.1 Effect of potato continuous cropping years on Chao1 and community richness of soil fungal communities
2.2 土壤真菌群落組成
對馬鈴薯土壤真菌群落的分布進行了分析(圖2),置換檢驗(Permutation test)表明,種植年限對馬鈴薯土壤真菌群落的分布有顯著影響(R2=0.56,P<0.001)。從圖2可以看出,C7和C9處理比較接近,說明這兩個處理的真菌群落相似度較高,他們和R處理的距離較遠,C1、C3和C5處理位于R處理和C7、C9處理之間,證明C7和C9處理與R處理的真菌群落相似度低,真菌群落差異顯著。
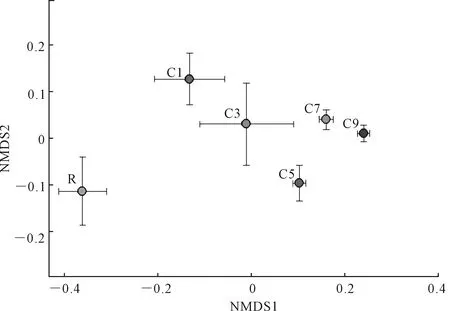
注:NMDS是基于真菌群落組成的布雷-柯蒂斯距離。誤差條表示標準誤差。Note:NMDS are based on Bray-Curtis distances according to fungal community composition. Error bars indicate standard errors.圖2 不同馬鈴薯連作年限下土壤真菌群落的非度量多維尺度(NMDS)圖Fig.2 NMDS plot showing the shift of fungal communities in relation to potato continuous cropping years
2.3 土壤綱水平真菌群落組成
對檢測到的>90%的真菌群落在綱水平的分類進行分析(圖3,見189頁),可以看出,試驗供試樣本主要富集的13個真菌綱:錘舌菌綱(Leotiomycetes)、傘菌綱(Agaricomycetes)、壺菌綱(Chytridiomycetes)、座囊菌綱(Dothideomycetes)、子囊菌綱(Ascomycota_unidentified)、糞殼菌綱(Sordariomycetes)、銀耳綱(Tremellomycetes)、盤菌綱(Pezizomycetes)、散囊菌綱(Eurotiomycetes)、酵母綱(Saccharomycete)、Fungi_unidentified、Archaeorhizomycetes、傘型束梗孢菌綱(Agaricostilbomycetes)。土壤真菌群落的相似度總體分為兩類,C7和C9處理為一組,其他處理分為一組。R、C1、C3、C5間差異較小,長期連作C7和C9間差異較小,而R、C1、C3、C5與C7、C9間差異較大,說明連作7 a后馬鈴薯田土壤真菌群落結構發生較大改變。再細分,R處理和C1、C3、C5處理分開,可以分為3組,這和圖2的真菌群落相似度分類相似,說明馬鈴薯輪作和連作也會引起土壤微生物群落一定的變化。
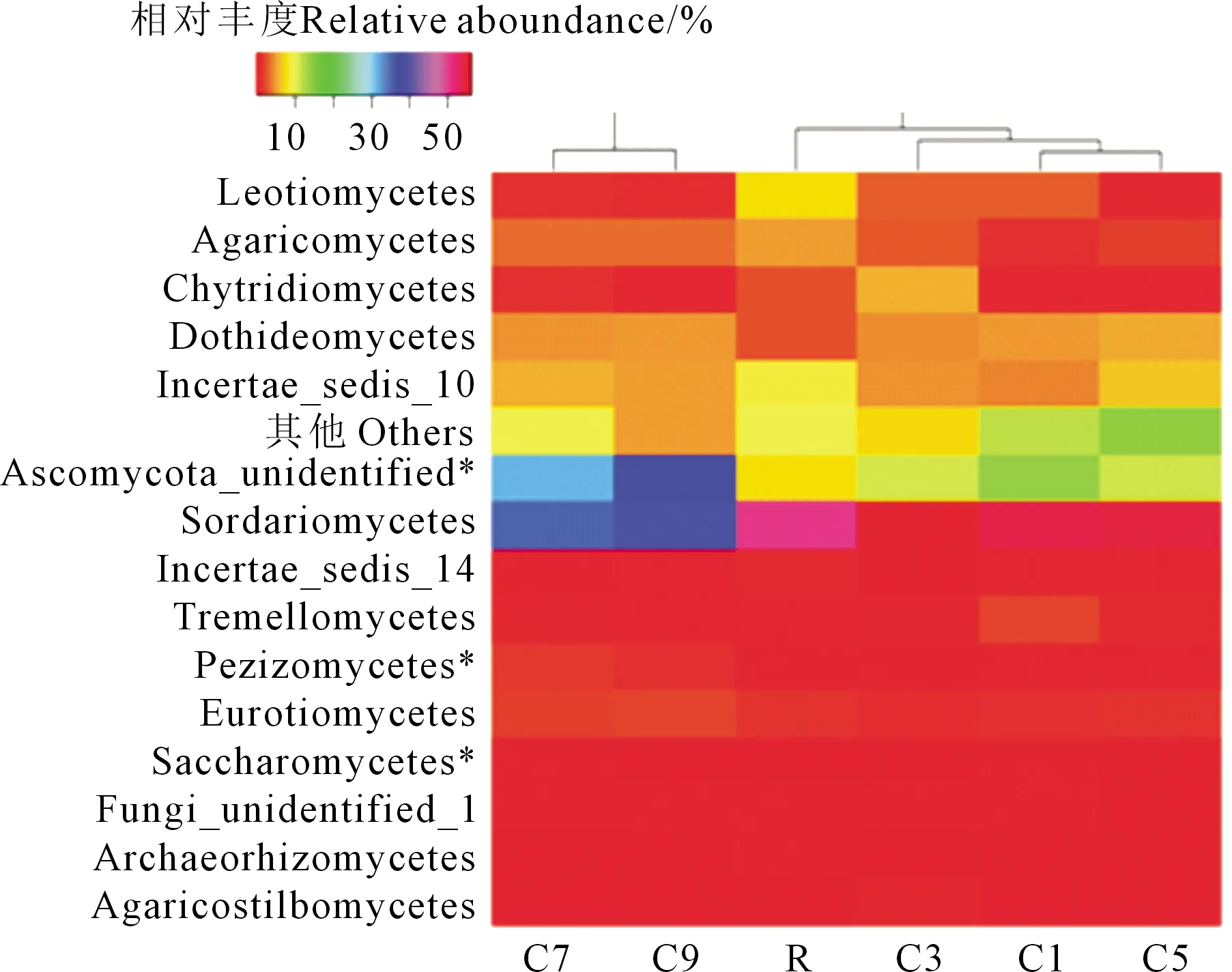
注:*表示處理間差異顯著(P<0.05),下同。Note: * shows significant differences among treatments (P<0.05),the same below.圖3 連作年限對馬鈴薯田土壤綱水平真菌群落影響的熱圖Fig.3 Heat map shows the effects of continuous cropping years on potato soil fungal community at class level
2.4 土壤有害真菌屬水平的分類與分布
在不同種植年限的馬鈴薯農田土壤中,共檢測到273個真菌屬,其中平均相對豐度>0.15%的真菌屬有44個,對檢測到的豐度排名前44的真菌屬進行差異分析,其中,13個真菌屬在不同處理的馬鈴薯農田土壤中差異顯著(圖4),分別為:鐮刀菌屬(Fusarium)、Nectriaceae、靈芝屬(Ganoderma)、Metarhizium、Lecanicillium、木生紅曲霉屬(Xylogone)、叢赤殼屬(Nectria)、Ascomycota_unidentified、曲霉屬(Aspergillus)、青霉屬(Penicillum)、頭梗霉(Cephaliophora)、隱球菌屬(Cryptococcus)、剛毛四枝孢菌屬(Tetracladium)。青霉菌在連作5、7 a和9 a的土壤中數量顯著高于輪作。潛在的有害病菌為鐮刀菌屬(Fusarium)、Nectriaceae、靈芝屬(Ganoderma)、綠僵菌屬(Metarhizium)、Lecanicillium、木生紅曲霉屬(Xylogone)、叢赤殼屬(Nectria),其中,鐮刀菌屬是引起馬鈴薯枯萎病的土傳病原真菌。鐮刀菌屬種類很多,本研究土壤中主要是尖孢鐮刀菌(F.oxysporum),而綠僵菌屬在馬鈴薯連作土壤中屬于拮抗菌,沒有致病性。
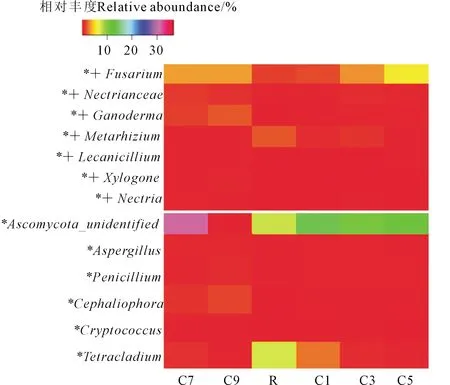
注:+表示該真菌屬為潛在的有害病菌。Note:+ indicates that the fungus is a potentially harmful pathogen.圖4 連作年限對馬鈴薯田土壤屬水平真菌群落影響的熱圖Fig.4 Heat map shows the effects of continuous cropping years on potato soil fungal community at genus level
土壤潛在有害病原真菌屬水平豐度聚類分析顯示(圖5),不同連作年限潛在病原菌群落可以分為兩組,C5、C7、C9處理為一組,R、C1、C3為一組,同一組內真菌群落組成類型相似。結果表明馬鈴薯連作5、7 a和9 a群落組成在屬水平上相似性較高,而馬鈴薯輪作、連作1 a、連作3 a群落組成在屬水平上相似性較高。由此可見,當馬鈴薯連作5 a以后,土壤潛在病原真菌種群豐度開始發生較大變化。病原菌鐮刀菌屬(Fusaruim)在C5中相對豐度達最高,顯著高于R、C1、C3以及C7、C9。拮抗菌綠僵菌屬(Metarhizium)在C5、C7、C9中相對豐度顯著高于R、C1、C3。
3 討 論
連作障礙的產生與土壤微生物的多樣性密不可分,土壤真菌多樣性和數量與土壤健康狀況呈負相關[16],長期連作使作物土壤微環境發生改變,土壤真菌多樣性和豐富度有所提高,破壞了原有土壤微生物群落的結構,影響作物的健康生長[17-18]。目前不同作物不同連作年限的研究結果不盡相同。趙存鵬等[6]研究得出,輪作后棉花根際土壤中真菌Chao1、ACE和Shannon指數顯著高于連作棉田。張子龍等[19]研究表明,小麥/三七輪作土壤真菌群落多樣性和豐富度顯著高于三七連作。劉珊廷等[20]研究表明木薯輪作根際土壤中Shannon指數和Chao1指數均高于連作。而本研究發現,隨著連作年限的增加,真菌多樣性指數呈上升趨勢。馬鈴薯連作5 a Richness顯著高于小麥/馬鈴薯輪作,馬鈴薯連作7 a和9 a菌群豐度顯著高于連作1、3 a以及小麥/馬鈴薯輪作,馬鈴薯連作3 a的Chao1顯著高于小麥/馬鈴薯輪作,馬鈴薯連作5、7 a和9 a與連作3 a和輪作處理間差異顯著。說明隨著馬鈴薯種植年限的延長,馬鈴薯田真菌多樣性呈上升趨勢。這與大豆連作20 a的根際土壤真菌種群的多樣性顯著高于3 a短期連作的研究結論一致[15]。
連作通常會使得土壤中真菌致病菌數量增加,植株根系釋放的分泌物也會促進病原菌的繁殖和生長,同時抑制有益細菌的生長,使病原菌成為優勢種群而引發土傳病害,使作物植株生長發育不良而引起產量和品質下降[21-22]。本研究中,從真菌屬分類水平進行分析,馬鈴薯土壤中優勢真菌屬包括鐮刀菌屬(Fusarium)、Nectriaceae、靈芝屬(Ganoderma)、綠僵菌屬(Metarhizium)、Lecanicillium、木生紅曲霉屬(Xylogone)、叢赤殼屬(Nectria)等致病病原菌;在我國,馬鈴薯土傳病害主要有枯萎病(Fusariumsp)、干腐病(Fusariumsp)、青枯病(Ralstoniasolanacearum)、黃萎病(Verticilliumdahliae)、黑痣病(Rhizoctoniasolani)、瘡痂病(Streptomycesscabies)、粉痂病(Spongosporasubterranean)等。已有研究顯示,鐮刀菌屬(Fusaruimspp.)是引起馬鈴薯枯萎病和干腐病的土傳病害致病菌[11]。本研究中,鐮刀菌屬在連作土壤中的豐度較高,尤其是連作第5年的土壤,這與劉星等[23]研究顯示,Fusariumsp.、Fusariumsolani在馬鈴薯連作土壤中數量均增加的結論一致。
馬鈴薯枯萎病病原菌種類隨地域不同而有差異,其致病力也不一致。引起枯萎病的病原菌鐮刀菌(Fusariumspp.)主要有茄病鐮刀菌(F.solani)、尖孢鐮刀菌(F.oxysporum)、接骨木鐮刀菌(F.sambucinum)、雪腐鐮刀菌(F.nivale)、串珠鐮刀菌(F.moniliforme)、三線鐮刀菌,銳頂鐮刀菌(F.acuminatum)和燕麥鐮刀菌(F.avenaceum)等[24]。河北馬鈴薯枯萎病病原菌為茄病鐮刀菌、串珠鐮刀菌和尖孢鐮刀菌[25]。內蒙古和甘肅馬鈴薯枯萎病病原菌有尖孢鐮刀菌、茄病鐮刀菌和三線鐮刀菌[25],甘肅馬鈴薯枯萎病病原菌還有燕麥鐮刀菌[26]。本研究中,馬鈴薯連作土壤中尖孢鐮刀菌(F.oxysporum)在連作5 a的土壤中大量富集,說明在甘肅定西馬鈴薯種植區內,尖孢鐮刀菌是導致馬鈴薯發病的主要致病菌。同時,尖孢鐮刀菌也是河北、內蒙、甘肅馬鈴薯枯萎病的主要致病菌。
然而,長期連作的效果不一定是負面的,有研究發現,作物連作可能引起拮抗菌逐年富集[27],某些有益菌數量也會增加[28],這些有益菌群通常產生一些激素類化合物,通過促進作物養分吸收、抑制病原菌生長等方式來促進植物生長。一些作物在一定連作年限后土壤逐步得到自我恢復,并且連作障礙程度趨于減輕甚至消失[29]。本研究中,Ascomycota_unidentified、曲霉屬(Aspergillus)、青霉屬(Penicillum)、頭梗霉(Cephaliophora)、隱球菌屬(Cryptococcus)、剛毛四枝孢菌屬(Tetracladium)等有益菌真菌屬在不同處理的馬鈴薯根部土壤中差異顯著。其中,青霉菌在連作5、7 a和9 a的土壤中數量顯著高于輪作。青霉菌(Penicillium)參與有機物的分解,促進土壤碳和氮的循環,降解土壤中的有害物質[30],其產生的代謝活性物質對部分鐮刀菌屬(Fusarium)病原菌產生明顯抑制作用[31]。這有可能是鐮刀菌在連作5 a的土壤中數量達到最高,在連作7 a和9 a的土壤中數量又開始顯著降低的原因,與大豆連作20 a的土壤中,鐮孢菌數量顯著低于3 a連作的結論一致[15]。本研究中,綠僵菌屬(Metarhizium)在馬鈴薯連作5、7 a和9 a的土壤中相對豐度顯著增加。綠僵菌屬(Metarhizium)可以對植物病害產生拮抗作用[32],同時促進植物對氮素營養的吸收,增強植物抗病性[33],還可以分泌各種有機酸溶解土壤中固態磷,提高作物養分利用率[34]。
4 結 論
1)與馬鈴薯輪作農田相比,馬鈴薯連作7 a和9 a的農田土壤真菌菌群豐度和Chao1顯著提高,說明馬鈴薯連作7 a后,農田土壤真菌群落的豐富度和多樣性增加。
2)對馬鈴薯土壤真菌群落的分布進行分析,表明C7和C9真菌群落結構相似度較高,C1、C3、C5真菌群落結構相似度較高,C7和C9與R的真菌群落結構差異較大,C1、C3、C5的真菌種群群落結構介于C7、C9與R之間。說明馬鈴薯長期連作會引起土壤微生物群落的變化,尤其是連作7 a真菌群落結構發生較大改變。
3)引起馬鈴薯枯萎病的土傳致病菌鐮刀菌屬(Fusaruim)在C5中相對豐度達最高,顯著高于R、C1、C3以及C7、C9。有益菌青霉菌(Penicillum)在C5、C7和C9的土壤中數量顯著高于R。拮抗菌綠僵菌屬(Metarhizium)在C5、C7、C9中相對豐度顯著高于R、C1、C3。馬鈴薯連作5 a和7 a的農田土壤中拮抗菌和有益菌大量富集,同時,土傳病原菌在連作5 a土壤中也大量富集,但是在連作7 a和9 a土壤中數量逐漸下降。
