黏蟲HSC70互作蛋白基因HIP的分子特征與時空表達分析
2022-07-30 08:08:08李鴻波符俊洪
江蘇農業科學 2022年13期
李鴻波, 符俊洪, 馮 磊
(1.貴州省農業科學院植物保護研究所,貴州貴陽 550006; 2.貴州大學昆蟲研究所,貴州貴陽 550025)
黏蟲別稱行軍蟲,屬鱗翅目夜蛾科,是我國和其他亞洲國家及澳洲的重大遷飛性害蟲。該蟲的寄主十分廣泛,可危害玉米、水稻、小麥等100多種作物,主要通過幼蟲取食寄主植物的葉片,影響作物的光合作用,從而造成農作物減產。上世紀50—80年代期間,黏蟲多次在全國范圍內暴發成災,造成嚴重的經濟損失。90年代后,隨著全國小麥種植面積銳減,加之測報與防治技術的完善,黏蟲得到有效控制。近年來,受耕作制度和全球氣候變化等因素的影響,黏蟲在我國局部地區暴發,其暴發頻率、危害程度及危害面積為近20年之最,對我國的糧食安全生產已構成嚴重威脅。黏蟲的暴發與其遷飛為害和暴食能力相關,其強大的生殖能力也是其暴發的重要原因。黏蟲無滯育特性,在適宜條件下1年可以繁殖數代,雌成蟲可多次交配,單雌產卵量達500~1 000粒。昆蟲的產卵主要受生殖相關基因的調控,因此開展黏蟲生殖相關基因的研究可望為該害蟲的防治提供新途徑。
卵黃發生是昆蟲生殖調控的關鍵,與昆蟲種群數量的消長密切相關。目前,關于黏蟲生殖的研究主要集中在卵黃原蛋白及其受體方面。例如,在黏蟲中已經報道了2個卵黃原蛋白(vitellogenin,簡稱Vg)及其受體(vitellogenin receptor,簡稱VgR)基因,發現這2個基因主要在黏蟲雌成蟲脂肪體和卵巢高表達;進一步研究發現,沉默相關基因后黏蟲卵的形態畸形,產卵量明顯下降,從而驗證了這2個基因在黏蟲生殖中的功能。昆蟲的生殖是一個復雜的過程,受多個基因的調控。熱激蛋白(HSPs)作為一種分子伴侶在昆蟲抵御環境脅迫中發揮重要作用,然而近年來的研究發現HSPs參與多種昆蟲的生殖。以飛蝗為例,研究發現和在飛蝗的脂肪體中高表達;沉默這2個基因后導致飛蝗卵黃蛋白表達水平顯著降低,同時卵母細胞發育遲緩;敲除和后飛蝗Vg的合成和卵母細胞的成熟嚴重受阻。這些結果表明,HSPs通過調控昆蟲Vg的合成而控制昆蟲的生殖。然而,HSPs是否參與黏蟲的生殖還未見報道。
HSC70互作蛋白(HSC70 interacting protein,簡稱HIP)是一種高度保守的協同分子伴侶。該蛋白主要通過由34個氨基酸組成的保守結構域TPR(tetratricopeptide repeats)與HSC70 C-端的EEVD結合,從而調控HSC70的功能。目前,HIP已在人類、擬南芥、果蠅、番茄等生物中被相繼報道,并已證實其在生物抵御各種生物脅迫中發揮重要作用。然而,HIP在參與動植物生殖的研究還未見報道。最近,筆者所在課題組在分析黏蟲4日齡雌蟲卵巢轉錄組數據時發現,在卵巢中高表達,暗示其可能在黏蟲的生殖中具有重要作用。為此,本研究采用RT-PCR從黏蟲克隆了的基因,并采用實時熒光定量PCR分析了其在黏蟲不同發育時期,4日齡雌成蟲不同組織中的表達模式,以期為進一步研究在黏蟲生殖中的功能奠定基礎。
1 材料與方法
1.1 試驗時間
試驗于2021年1—3月在貴州省植物保護研究所完成。
1.2 供試蟲源及試劑
供試蟲源:本試驗所用黏蟲為長期飼養在貴州省植物保護研究所的室內種群,至今飼養30代以上,期間未接觸任何殺蟲劑。飼養條件為光—暗周期 14 h—10 h、溫度為24~25 ℃、濕度為70%~80%,具體飼養過程參照李鴻波等的方法進行。
供試試劑:EastepR Super Total RNA isolation Kit購自Promega公司;cDNA第1鏈反轉錄試劑盒購自Thermo 公司;iScriptcDNA Synthesis Kit和SsoAdvancedUniversal SYBR Green Supermix購自Bio-RAD公司;LA酶、DL2000 marker等購自TaKaRa公司;其他試劑為國產分析純。
1.3 總RNA提取,cDNA第1鏈合成和RT-PCR
按照Eastep? Super Total RNA isolation Kit提供的方法提取樣品總RNA,利用DNAaseI 去除基因組DNA,并采用oligo(dT)引物(Thermo Scientific,USA) 合成 cDNA第1鏈,存于-20 ℃下備用。基于筆者所在實驗室前期的轉錄組數據分析,鑒定得到1個長1 218 bp的基因序列,初步分析發現該基因具有完整的開放閱讀框。據此序列設計1對特異性引物,以4日齡雌成蟲卵巢反轉錄而來的cDNA為模板,采用RT-PCR進行擴增(表1),擴增參數為:94 ℃ 預變性3 min;94 ℃ 變性30 s,54 ℃ 退火 1 min,72延伸30 s,共30個循環;最后在72 ℃下再延伸10 min。 反應結束后取PCR產物進行瓊脂糖凝膠電泳,將符合目標條帶大小的PCR產物送上海生物工程有限公司測序。
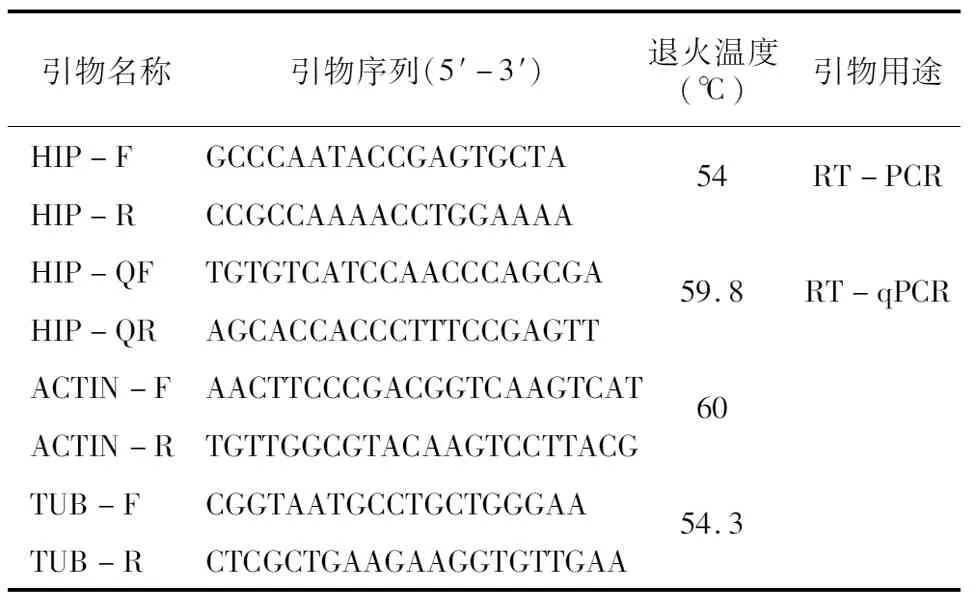
表1 試驗中的引物信息
1.4 基因序列分析和進化樹構建
采用ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)鑒定的開放閱讀框和氨基酸序列;采用Compute pI/Mw(https://web.expasy.org/compute_pi/)鑒定分子量和等電點;利用NCBI CDD(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)鑒定HIP的保守結構域;使用MEGA 5.1中的臨接法(NJ)構建系統發育樹,Bootstrap取樣值設置為1 000次。
1.5 MsHIP的時空表達分析
1.5.1 樣品收集 收集黏蟲不同發育階段(包括卵、1~6齡幼蟲、蛹和4日齡雌成蟲)和4日齡雌成蟲不同組織(頭、胸部、觸角、翅、中腸、脂肪體和卵巢)樣品,立刻用液氮速凍,存于-80 ℃下,用于總RNA的提取。每個樣品重復4次。
1.5.2 Real-time PCR分析 基于基因序列設計特異性引物,以和為內參基因,并以不同發育階段和不同組織樣品的cDNA為模板,在CFX-96 PCR儀(Bio-Rad,USA)進行qRT-PCR。PCR反應體系為20 μL,包括SsoAdvancedUniversal SYBR Green Supermix(Bio-Rad) 10 μL,上下游引物各1 μL(表1),模板cDNA 1 μL,ddHO 7 μL。qRT-PCR程序為95 ℃預變性3 min;95 ℃變性15 s,退火溫度下 30 s,共35個循環,反應結束后確認擴增曲線和熔解曲線。采用2-ΔΔ法計算黏蟲在不同發育階段和不同組織中的相對表達量。每個樣品含4個生物學重復,每個生物學重復含3個技術重復。
1.6 數據分析
所有表達量數據表示為平均值±標準誤。采用DPS 17.0中的單因素方差分析法(One-Way ANOVA)分別分析不同發育時期和不同組織處理表達量間的差異顯著性(=0.05)。
2 結果與分析
2.1 MsHIP的序列特征分析
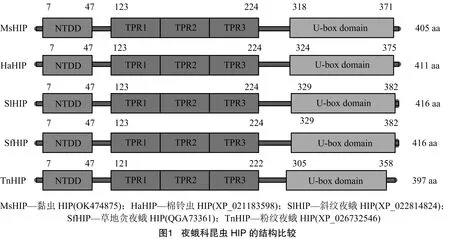
通過RT-PCR獲得的序列全長為1 215 bp,命名為,GenBank登錄號為OK474875。編碼405個氨基酸(aa),預測的分子量為44.04 ku,等電點為4.93。采用NCBI中的保守結構域搜索發現,中存在昆蟲HIP的3個保守結構域,分別是1個位于N-端的二聚體結構域(7~47 aa),1個TPR結構域(TPR1,123~156 aa,TPR2,157~190 aa,TPR3,191~224 aa)和1個位于C-端的U-box結構域(圖1)。多序列比對分析發現,夜蛾科昆蟲的HIP結構比較保守,均具有定義HIP蛋白的3個保守結構域(圖1)。
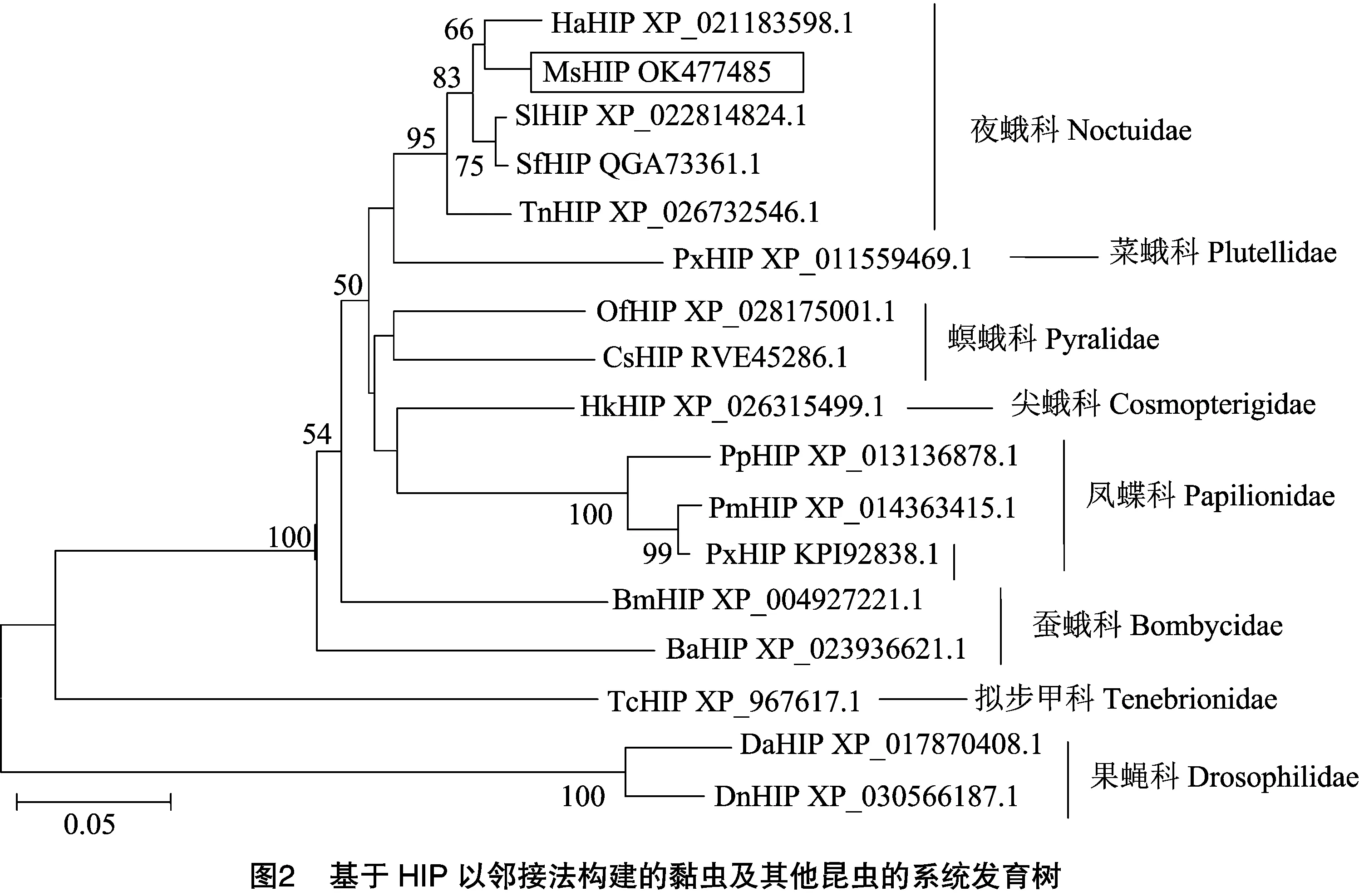
2.2 鱗翅目昆蟲HIP系統發育分析
通過NCBI搜索比對,發現黏蟲的HIP氨基酸序列與棉鈴蟲()的 HIP 氨基酸序列相似度最高,為86.62%,其次為草地貪夜蛾()(86.54%)和斜紋夜蛾()(85.34%),與粉紋夜蛾()的相似度為82.57%。系統發育樹分析顯示,該系統樹被明顯分為2支,即鱗翅目和雙翅目。在鱗翅目夜蛾科中,黏蟲HIP與棉鈴蟲HIP的親緣關系最近,以較高的支持率(66.00%)聚在一起(圖2)。
2.3 MsHIP時空表達分析
在黏蟲各發育階段的表達差異顯著(=5260,=8,35,<0.001)。其中,以4日齡成蟲的表達量最高,是對照(卵)的45.28倍;其次為6齡幼蟲,表達量是對照的36.90倍;在其余發育階段的表達量較低,均與對照相比差異不顯著(圖3-A)。
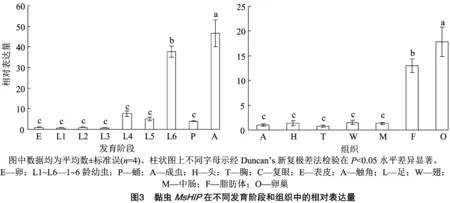
在4日齡成蟲不同組織中表達差異顯著(=3052,=6,27,<0.001)。其中,在卵巢中的表達水平最高,是對照(頭)的18.55倍;其次為脂肪體,其表達水平是對照的13.58倍;在胸、觸角、翅和中腸中表達水平較低,與對照相比差異不顯著。
3 結論與討論
HSPs作為分子伴侶在調控昆蟲生殖中的作用已得到證實。本研究基于前期的轉錄組數據分析,結合RT-PCR獲得的全長序列,是GenBank中登錄的第24個昆蟲的HIP序列。同已報道的HIP一樣,MsHIP含有3大保守結構域,即二聚體結構域、TPR結構域和U-box結構域。N-端的二聚體結構域的主要功能是作為一種調控因子調控HSP70的功能;TPR是由34個氨基酸組成的多肽,其功能主要是直接與HSP/HSC70和HSP90 中ATPase 結構域結合,減緩ADP的釋放速度;U-box具有E3泛素化抑制劑連接酶和多聚泛素化鏈延伸活力,有助于加速依賴泛素化的分子伴侶基質的降解。基于鱗翅目昆蟲HIP的系統發育分析顯示,黏蟲主要與斜紋夜蛾、草地貪夜蛾、粉紋夜蛾和棉鈴蟲聚在一起,且與棉鈴蟲的親緣關系最近,支持率高達66.00%,同屬夜蛾科。這幾種昆蟲HIP的分子結構、氨基酸相似度(82.57%~86.62%)及田間多食性特點都佐證了以上結論,表明他們可能來源于同一祖先,因自然選擇壓力而發生分化。
HSPs的表達具有發育階段和組織特性。本研究發現,在所有發育時期均表達,但其在不同時期的表達模式存在差異,這與已報道的其他昆蟲的HSPs的表達規律不一致。例如,赤擬谷盜在蛹早期和成蟲早期的表達量最高;褐飛虱、、、和在成蟲中的表達水平最高,則在5齡幼蟲中表達量最高,而和在所測試的發育階段表達量都很低;對三葉斑潛蠅()而言,在所有發育階段的表達十分恒定,在蛹和成蟲中高表達,而則只在雌成蟲中高表達。在黏蟲中,在2齡和6齡幼蟲中高表達,則在黏蟲1 齡幼蟲中高表達,而在6齡幼蟲和4日齡雌成蟲高表達。這些結果說明,不同的HSPs在不同物種的生長發育過程中發揮不同的作用。此外,研究發現在4日齡雌成蟲中高表達的現象與黏蟲卵黃原蛋白受體基因()和脂蛋白受體基因()(另文發表)的表達規律基本一致,表明可能參與黏蟲的生殖。
為進一步明確該基因是否在黏蟲的生殖中發揮作用,進一步檢測了其在成蟲不同組織中的表達情況。結果顯示,在卵巢和脂肪體中高表達,這與棉鈴蟲、大螟和草地貪夜蛾在不同組織中的表達規律不一致,但與褐飛虱中、、、的表達規律類似。和在昆蟲的生殖中發揮重要作用,其中脂肪體是卵黃原蛋白合成的主要場所,而卵巢則是卵母細胞發育的主要場所。李杰研究發現,在黏蟲脂肪體和卵巢中高表達,采用RNAi沉默該基因后,其在脂肪體和卵巢中的表達水平顯著降低,同時卵的發育畸形,產卵量顯著減少,從而證明了在黏蟲的生殖中發揮重要作用。基于與在黏蟲雌蟲生殖器官中表達規律的一致性,因此推測在黏蟲的生殖中也發揮重要作用。
綜上,本研究首次從黏蟲中克隆了基因,發現具有昆蟲定義的3個保守結構域,并且在黏蟲的雌成蟲及脂肪體和卵巢中高表達,表明其在黏蟲生殖中具有重要作用。
