應用脂肪酸組成研究熱帶東太平洋同域中上層鯊魚營養生態位分化
2022-07-30 11:09:28李云凱
生態學報 2022年13期
李云凱,徐 敏,貢 藝,4,*
1 上海海洋大學海洋科學學院, 上海 201306
2 農業農村部遠洋與極地漁業創新重點實驗室, 上海 200009
3 青島海洋科學與技術試點國家實驗室海洋漁業科學與食物產出過程功能實驗室, 青島 200071
4 大洋漁業資源可持續開發省部共建教育部重點實驗室, 上海 201306
探究有限資源環境中物種多樣性維持及物種共存機制是群落生態學的核心問題[1]。各物種特征的權衡及組合的不同決定了其生活史對策的不同,由此決定了各物種在群落中生態位的差異化[2]。其中,基于物種對食物資源利用差異的營養生態位分化是物種共存的必要條件之一[3]。
大洋中上層鯊魚多處于食物網的頂端或近頂端,其可通過下行效應(top-down control)對食物網結構和功能起調控作用[4],但對同域分布中上層鯊魚的物種多樣性維持及生態共存機制仍不明晰,尤其在資源相對貧瘠的開闊大洋。熱帶東太平洋海域主要棲息有8種中大型中上層鯊魚物種,包括大青鯊(Prionaceglauca)、鐮狀真鯊(Carcharhinusfalciformis)、長鰭真鯊(C.longimanus)、大眼長尾鯊(Alopiassuperciliosus)、淺海長尾鯊(A.pelagicus)、尖吻鯖鯊(Isurusoxyrinchus)、路氏雙髻鯊(Sphyrnalewini)和錘頭雙髻鯊(S.zygaena)[5],為東太平洋的頂級捕食者。對其營養生態位的比較研究有助于揭示大洋同域分布頂級捕食者的共存機制。
脂肪酸組成分析是近年來在生態學領域興起的一項新方法,在海洋生物研究領域,該方法已開始應用于頭足類[2]和硬骨魚類[6]的營養生態位研究,而對鯊魚的研究較少。作為頂級捕食者,鯊魚自身合成的脂肪酸極少,且主要為短鏈和去飽和度較低的種類[7],而其攝入的外源性脂肪酸通常以保守形態沉積于組織中,可整合一段時間內的攝食信息[8]。其中某些脂肪酸在生物的食性和營養關系研究中具有指示作用,如捕食者機體內Σ22+Σ24的含量可追溯其陸源植物食性,DHA/EPA(C22:6n3/C20:5n3)含量比可用于指示其營養級[9]。因此,脂肪酸組成可用于反映鯊魚與其食物間的動態關系和營養流動[10],進而用于探究鯊魚的營養生態位。Bierwagen等通過肌肉和血漿脂肪酸組成分析研究了澳大利亞東北海域兩種同域礁鯊——灰三齒鯊(Triaenodonobesus)和鈍吻真鯊(Carcharhinusamblyrhynchos)的資源分配及利用,結果表明,兩種鯊魚存在激烈的資源競爭;灰三齒鯊營養生態位較小,表現為其食性的特化[11]。本研究通過測定熱帶東太平洋8種中上層鯊魚肌肉的脂肪酸組成,分析其食性差異、營養關系及營養生態位分化,探討其資源分配方式及同域共存機制。
1 材料與方法
1.1 樣品采集

圖1 熱帶東太平洋鯊魚樣品捕獲地點 Fig.1 Sampling location of pelagic sharks in the tropical Eastern Pacific
鯊魚樣本采集自2014年6月—11月我國金槍魚延繩釣兼捕漁獲。采樣范圍為8°—10°N,115°—125°W的東太平洋海域(圖1)。共采集到鯊魚72尾,具體生物學信息見表1。取魚體臀鰭附近白肌樣本,真空包裝低溫運至實驗室,于-20 ℃冰箱冷凍保藏。
1.2 脂肪酸分析與數據處理
肌肉樣品去除表皮,使用超純水漂洗后放入冷凍干燥機(Christ Alpha 1- 4)-55 ℃干燥24 h,用冷凍混合球磨儀(Mixer mill MM440)磨碎成粉末(直徑5 μm)。稱取樣品粉末200 mg于具塞離心管中,加入12 mL二氯甲烷-甲醇溶液(2/1,體積分數),室溫下浸泡24 h。離心后取上清液,加入4 mL 0.9 %的氯化鈉溶液洗滌,靜置至溶液明顯分層。收集下層二氯甲烷層至圓底燒瓶中,用氮氣(氮吹儀)吹掃除去有機試劑后,加入4 mL氫氧化鈉—甲醇溶液(0.5 moL/L)復溶,獲得肌肉總脂提取液。
將盛有總脂提取液的圓底燒瓶連接水浴回流裝置加熱20 min后,加入4 mL三氟化硼-甲醇溶液(14%,w/v),在60 ℃水浴中進行甲酯化反應20 min,反應結束冷卻至室溫,再加入4 mL正己烷,震蕩30 s后,加入飽和氯化鈉溶液定容。正己烷層移入配有聚四氟乙烯瓶蓋的后壁玻璃管。脂肪酸采用氣相色譜/質譜聯用儀(7890B/5977A,Agilent)進行分析。色譜條件:毛細管柱型號為AgilentHP- 88(60 m×0.25 nm×0.20 μm),載氣為高純氦氣,分流比為10:1,進樣口溫度為250 ℃。升溫程序:初始溫度為125 ℃,以8 ℃/min升溫至145 ℃并保持26 min,然后以2 ℃/min升溫至220 ℃并保持1 min,最后以1 ℃/min升溫至227 ℃并保持1 min。
以37種脂肪酸混標及內標十九烷酸甲酯標品作為標準,通過比對保留時間對脂肪酸進行定性分析,采用內標法(GLC 37,Nu-Chek Prep Inc,Elysian)對脂肪酸進行定量分析
其中,Xi是待測樣品中組分i的含量(mg/g);Ai是組分i的峰面積;m是樣品質量;m19是內標物十九烷酸甲酯的質量;A19是待測樣品中內標物的峰面積;Fi是組分i與內標物的校正因子之比,稱為相對校正因子[12—13]。
計算各脂肪酸占總脂肪酸含量的百分比,整體分析時去除小于0.5%的數據[14]。

表1 鯊魚采樣信息及基礎生物學參數
1.3 數據分析
鯊魚脂肪酸組成測定結果以“平均值±標準偏差”(mean±SD)表示,采用單因素方差分析(analysis of variance, ANOVA)檢驗各脂肪酸含量是否存在顯著種間差異。聚類分析(cluster analysis)用以分析數據之間的相似性,明確劃分成組。主成分分析(principal component analysis,PCA)可用于探究多元事物的主要影響因素,通過降維將多個相互關聯的數值轉化成少數互不相關的指標。因此本研究進一步應用PCA將脂肪酸測定結果降至二維,并利用R語言中的SIAR軟件包計算標準橢圓面積(SEAc),以表征各鯊魚物種的營養生態位[15]。繪圖使用Origin 2021 pro和R軟件4.0.2操作。
2 結果與分析
2.1 脂肪酸組成
8種鯊魚肌肉中共檢測得脂肪酸35種,含量大于0.5%的共21種(表2)。飽和脂肪酸含量范圍為39.29%—48.64%,種間差異顯著(P<0.05),主要種類為C14:0、C16:0、C17:0、C18:0、C20:0、C22:0、C23:0和C24:0。其中,C16:0和C18:0含量最高,分別為13.22%—25.18%和15.22%—28.52%。兩種飽和脂肪酸含量存在顯著種間差異(P<0.05)。

表2 鯊魚肌肉脂肪酸組成/%
單不飽和脂肪酸含量范圍為8.75%—15.08%,種間差異顯著(P<0.05),主要種類為C16:1n7、C18:1n9和C24:1n9。其中,C18:1n9含量較高,為3.11%—5.93%。長鰭真鯊肌肉中C18:1n9含量最高,錘頭雙髻鯊最低,種間差異顯著(P<0.05)。
多不飽和脂肪酸含量范圍為39.46%—49.38%,種間差異顯著(P<0.05),主要種類為C18:2n6、C20:2、C20:3n3、C20:3n6、C20:4n6、C22:2n6、C20:5n3(EPA)和C22:6n3(DHA)。其中,C20:3n3、C20:4n6、C20:5n3和C22:6n3含量較高,分別為3.95%—9.13%、3.96%—9.20%、0.64%—3.34%和22.32%—33.89%,錘頭雙髻鯊肌肉中C20:3n3和C20:4n6含量最高,大青鯊肌肉中C20:5n3含量最高,淺海長尾鯊肌肉中C22:6n3含量最高,此4種脂肪酸含量種間差異顯著(P<0.05)。此外,∑22+∑24含量范圍為27.35%—38.38%,種間差異顯著(P<0.05),大青鯊低于其余7種鯊魚。鯊魚肌肉中DHA/EPA含量的比值,尖吻鯖鯊(50.68)>淺海長尾鯊(33.72)>大眼長尾鯊(33.12)>鐮狀真鯊(21.34)>錘頭雙髻鯊(15.21)>路氏雙髻鯊(14.05)>長鰭真鯊(9.76)>大青鯊(8.38)。
2.2 聚類分析
為進一步探究8種鯊魚的脂肪酸組成特性關系,依據含量大于0.5%的21個脂肪酸指標進行的聚類分析結果顯示(圖2),樹狀圖在距離8—10內分成兩組,第一組為大眼長尾鯊、淺海長尾鯊和尖吻鯖鯊,屬鼠鯊目;第二組為大青鯊、鐮狀真鯊、長鰭真鯊、路氏雙髻鯊和錘頭雙髻鯊,屬真鯊目,其中,大青鯊、鐮狀真鯊和長鰭真鯊屬真鯊科,路氏雙髻鯊和錘頭雙髻鯊屬雙髻鯊科。同一組鯊魚在聚類樹狀圖上的歐氏距離相距較短,表明其肌肉脂肪酸組成較為相似。

圖2 聚類分析圖Fig.2 Cluster analysis of fatty acid profilesBSH:大青鯊 blue shark;SPL:路氏雙髻鯊 scalloped hammerhead shark;SPZ:錘頭雙髻鯊 smooth hammerhead shark;FAL:鐮狀真鯊 silky shark;OCS:長鰭真鯊 oceanic whitetip shark;BTH:大眼長尾鯊 bigeye thresher shark;PTH:淺海長尾鯊 pelagic thresher shark;SMA:尖吻鯖鯊 shortfin mako shark
2.3 主成分分析
由主成分散點圖可知(圖3),3種鼠鯊散布在相同區域,表明其脂肪酸組成相似。但3種鼠鯊與5種真鯊基本散布在不同區域,表明其脂肪酸組成存在差異。負荷圖可反映造成差異的主要脂肪酸種類。依據負荷圖,3種鼠鯊肌肉中C16:0、C16:1n7和C22:6n3含量更高,而5種真鯊C18:0、C20:4n6、C24:1n9和C20:5n3含量更高。主成分分析結果與聚類分析結果相似。營養生態位寬度及重疊見表3,大青鯊和鐮狀真鯊占據較大的營養生態位寬度,尖吻鯖鯊和路氏雙髻鯊占據的營養生態位寬度較小。3種鼠鯊與5種真鯊營養生態位存在明顯分離或有較小重疊。大眼長尾鯊和淺海長尾鯊營養生態位重疊程度較高。
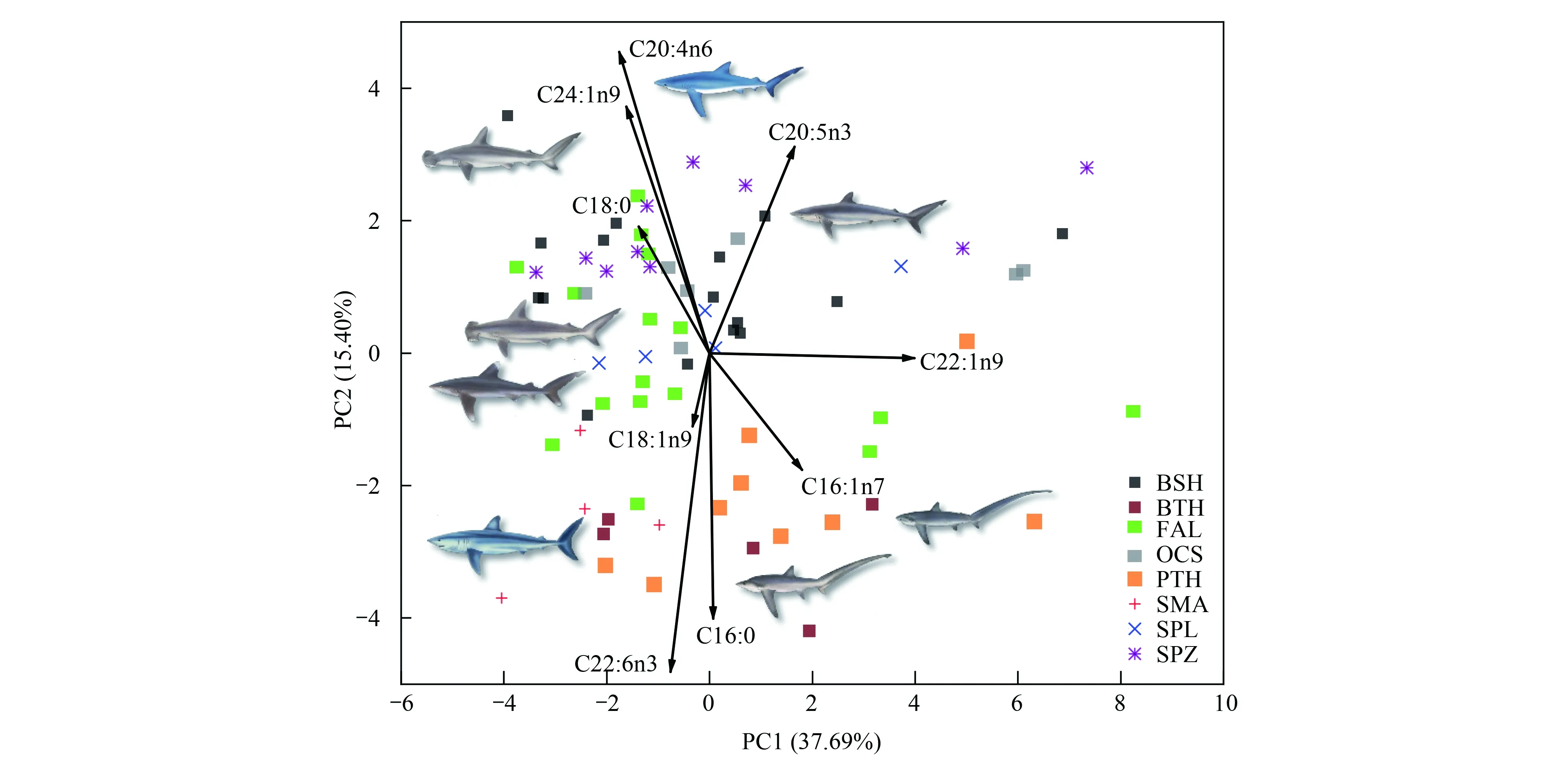
圖3 主成分分析圖Fig.3 Principal component analysis of fatty acid profiles
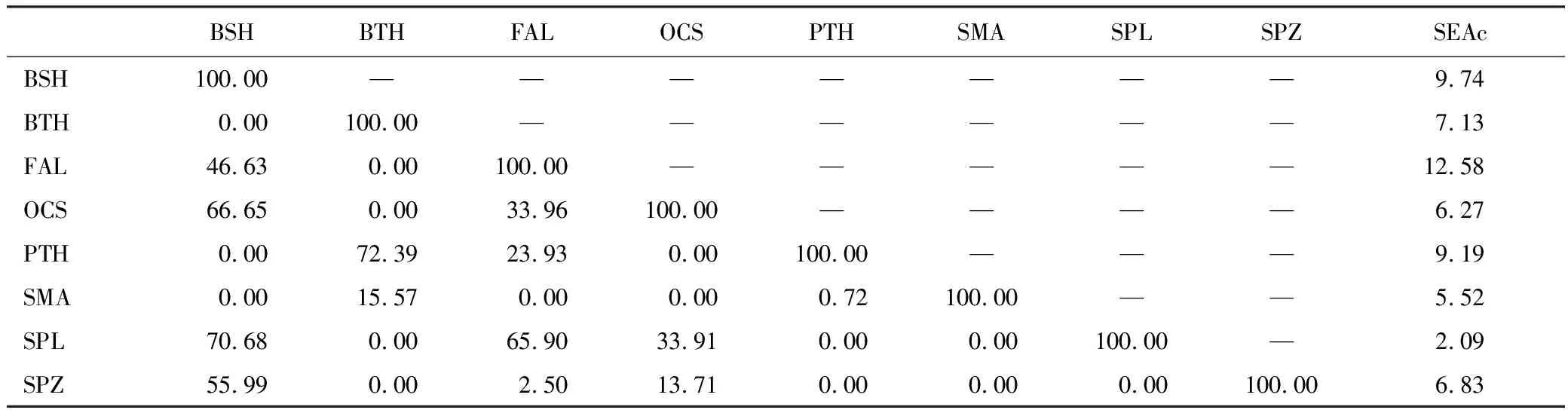
表3 東太平洋8種鯊魚肌肉營養生態位面積(SEAc)及重疊面積百分比/%
3 討論
本研究以東太平洋海域8種中上層鯊魚為研究對象,通過分析肌肉組織脂肪酸組成探討其營養生態位分化。8種鯊魚肌肉中飽和脂肪酸和多不飽和脂肪酸占比較大,而單不飽和脂肪酸占比較低。這與Cárdenas-Palomo等對加勒比海鯨鯊(Rhincodontypus)及Davidson等對南非東海岸9種大型鯊魚的研究結果一致[16—17]。鯊魚多為頂級捕食者,其自身合成的脂肪酸種類較少,主要有C14:0、C16:0和C18:0以及它們在Δ9去飽和酶作用下產生的C14:1n5、C16:1n7和C18:1n9脂肪酸[7]。必需脂肪酸(essential fatty acid)如C20:5n3、C22:6n3和C20:4n6等僅能由初級生產者或細菌合成,并沿食物鏈保守性儲存于捕食者體內,可用于指示鯊魚的攝食信息[10]。其中,C20:5n3主要由硅藻自身合成,且在頭足類體內含量較高[18—19],本研究中大青鯊和鐮狀真鯊肌肉中C20:5n3含量較高,表明這2種鯊魚所處食物鏈的初級生產者可能為硅藻,且偏好捕食頭足類,這與2種鯊魚的胃含物分析結果一致[5]。C22:6n3是甲藻的特征脂肪酸,也可指示肉食性程度[20—21],路尖吻鯖鯊,淺海長尾鯊和大眼長尾鯊肌肉中較高含量的C22:6n3表明其可能使用由甲藻主導的營養途徑且肉食性程度較高。路氏雙髻鯊肌肉中20:4n6含量較高,而C20:4n6多由雙殼類和甲殼類選擇性的保留在體內,可用于指示捕食者對甲殼類的攝食[22]。Flores-Martínez等研究發現,路氏雙髻鯊存在垂直運動模式,捕食底棲硬骨魚類和甲殼類[23]。Σ22+Σ24含量可一定程度上反映陸源植物食性[24],大青鯊肌肉中Σ22+Σ24含量遠低于其余7種鯊魚,說明其可能因終生的大洋性生活史特征,受陸源因素影響較小。研究發現,魚類肌肉中DHA/EPA比值與營養級成正比[20]。本研究中尖吻鯖鯊比值最高,為50.68,而大青鯊僅為8.38。Silveira等利用穩定同位素技術分析了加利福利亞西南海域5種鯊魚的營養級,發現尖吻鯖鯊營養級最高,而大青鯊則相對較低[25]。
營養生態位重疊反映了物種間對資源利用的相似程度及物種間潛在的競爭關系,而營養生態位分化可表征種間食性差異及棲息地分化[10]。淺海長尾鯊和大眼長尾鯊營養生態位高度重疊,反映了淺海長尾鯊和大眼長尾鯊可能存在較為激烈的資源競爭關系[26]。種間攝食空間隔離是緩解食物資源競爭的有效方法[3],大眼長尾鯊奇網(rete mirabile)發達,可維持顱內溫度[27],較淺海長尾鯊更適應在深海捕食。尖吻鯖鯊、大眼長尾鯊和淺海長尾鯊與其他5種鯊魚營養生態位存在明顯分離或僅有較小重疊,表明其種間食物資源及棲息地利用的競爭程度較弱。三種鼠鯊目鯊魚中,尖吻鯖鯊屬溫血性(endothermic)鯊魚,體溫可較環境溫度高達7℃,捕食的環境適應性更強,區域更廣[28]。胃含物分析也同樣發現,尖吻鯖鯊和大眼長尾鯊可在深層捕食硬骨魚類,鐮狀真鯊和大青鯊則主要捕食中上層頭足類[29]。
生態位寬度反映了生物對食物資源和棲息地利用的競爭能力[10]。大青鯊和鐮狀真鯊占據較大的營養生態位寬度,表征大青鯊和鐮狀真鯊對食物資源和棲息地利用的競爭能力較強,食性相對泛化[26]。Estupián-Montao等發現,1—2齡的大青鯊游向近岸覓食,而3—6齡大青鯊更傾向于遠洋覓食,對食物資源的利用范圍相對較廣[30]。胃含物分析表明,大青鯊主要以頭足類、硬骨魚類和甲殼類為食[5],這也一定程度上解釋了大青鯊營養級相對較低的原因。Flores-Martínez等研究顯示,鐮狀真鯊的攝食存在性別間差異并在生長發育過程中存在食性轉化,雌性鐮狀真鯊幼鯊喜好捕食密斑刺鲀(Diodonhystrix),雄性喜好捕食莖柔魚(Dosidicusgigas),而成年雄性捕食巴鰹(Euthynnusaffinis)[23],本研究中的鐮狀真鯊同時存在幼體和成體樣本,可能對結果有一定的影響。尖吻鯖鯊營養生態位寬度較窄,是由于本研究中的尖吻鯖鯊個體較其他鯊魚相對較小且均未達到性成熟,可能對食物資源的需求較為單一,胃含物分析也顯示,尖吻鯖鯊幼鯊喜好捕食硬骨魚類,如劍魚(Xiphiasgladius)和頜針魚(Belonebelone)等[5,31]。
綜上所述,本研究量化分析了東太平洋8種中上層鯊魚的資源分配方式,探討了其同域共存機制。其中,3種鼠鯊與5種真鯊存在食性差異或棲息地隔離。大青鯊與鐮狀真鯊生態位寬度較大,表征其對環境的可塑性較強,而尖吻鯖鯊和路氏雙髻鯊生態位寬度較小,表現為其食性的特化。盡管脂肪酸組成分析為研究種間資源利用模式,探究同域物種共存機制提供了新的方法。但魚類生理差異(個體大小、游動速度及體內關鍵代謝酶活性等)可影響外源性脂肪酸在其體內的同化程度,進而影響脂肪酸組成[32—33]。目前在攝食生態學中,脂肪酸組成分析仍主要起定性或半定量的輔助功能,未來應優化脂肪酸組成定量分析方法,結合穩定同位素和胃含物DNA條形碼技術從多維度完善大洋性鯊魚攝食生態研究內容,揭示這一特殊捕食群體的共存機制。
猜你喜歡
今日農業(2021年11期)2021-08-13 08:53:34
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
作文周刊·小學一年級版(2017年17期)2017-06-27 19:32:55
廣東第二課堂·小學(2017年2期)2017-02-20 15:10:08
