粉壟耕作對耕地土壤酶活性、微生物群落結構和功能多樣性的影響
2022-08-03 02:53:14陳彥云夏皖豫
生態學報 2022年12期
關鍵詞:功能
陳彥云,夏皖豫,趙 輝,曾 明
1 寧夏大學生命科學學院,銀川 750021 2 寧夏大學生態環境學院,銀川 750021 3 寧夏科技特派員創業指導服務中心,銀川 750021
土壤微生物是生態系統的重要組成部分,也是土壤生態系統的核心,直接或間接參與調節土壤養分循環、能量流動、有機質轉換、土壤肥力形成、污染物的降解及環境凈化等,特別在維持生物多樣性、改善土壤碳固存、生態系統功能和自然和農業生態系統中的養分循環方面發揮著重要作用[1—3]。土壤微生物組成的質與量的變化是土壤健康狀況的重要敏感指示,土壤微生物多樣性可以定義為生命的豐富度,代表著微生物群落的穩定性,也反映土壤生態機制及土壤脅迫對微生物群落的影響,因此對土壤微生物群落結構和功能的研究一直是土壤與植物營養學的研究熱點[4—6]。研究表明,土壤微生物群落受到各種因素的影響,包括耕作方式、氣候、施肥和pH,其中耕作方式是對土壤微生物的影響尤為關鍵[7—10]。
玉米(ZeamaysL.)是禾本科玉蜀黍屬一年生草本植物,是重要的糧食作物和飼料作物,也是全世界總產量最高的農作物。耕作方式一直是提高土壤肥力和提高作物生產力的最重要的農業實踐之一,大量研究表明不同耕作方式對土壤特性和作物產量的影響[11—14]。寧夏引黃灌區的玉米種植大多采用傳統耕作。然而,傳統耕作導致水土流失加劇,環境污染加劇,土壤退化,犁盤緊湊且封閉,阻礙了深層土層中土壤水、肥和熱量的循環影響生態系統功能[15—18]。粉壟耕作是一種集深松、深耕、旋耕、垂直耕作優點于一體的深耕技術[19]。它由配備六個立式螺旋鉆的新型強力機器進行,可以在不擾亂土壤層和生產新的硬質犁的情況下打碎壓實的犁盤并松動土層[20]。有研究表明,粉壟耕作模式對農田土壤質量、蓄水能力、作物產量、根系活力以及土壤微生物生物量和微生物多樣性等方面的有益作用更強[19,21—24]。近年來,粉壟耕作技術在農業生產方面取得了巨大成功。然而,目前尚不清楚粉壟耕作對耕地土壤微生物群落和土壤生態系統功能的影響。
本研究利用高通量 16S rRNA和ITS基因 Illumina 測序和Biolog-ECO方法,系統地研究了粉壟耕作對土壤中微生物群落結構和土壤微生物功能多樣性的影響。本研究的目的是:(1)探尋不同耕作方式下耕地土壤酶活性、土壤微生物多樣性和群落結構的差異;(2)分析不同耕作方式下土壤微生物群落功能多樣性及功能基團;(3)粉壟耕作如何影響土壤微生物結構、酶活性與微生物功能多樣性間的相關性,使玉米產量得到提升。本研究為粉壟耕作技術在寧夏引黃灌區作物栽培中的應用發展提供了科學理論依據和農業實踐理論支撐。
1 材料與方法
1.1 試驗樣地
試驗于2019年4月—10月在寧夏回族自治區石嘴山市平羅縣頭閘村(106°63′N、38°95′E)進行,屬溫帶大陸性氣候,海拔1091 m,日照充足、晝夜溫差大、干旱少雨、年蒸發量1755 mm,平均風速2—3 m/s,無霜期171 d,年降雨量為184 mm,年均氣溫8.21℃。試驗區土壤為黏土,土壤狀況為全磷0.86 g/kg,全氮1.10 g/kg,堿解氮76.92 mg/kg,速效磷34.69 mg/kg,速效鉀214.03 mg/kg,有機碳11.42 mg/kg,總鹽0.67 g/kg,含水量12.38%,pH值為8.5。
1.2 試驗處理與土樣采集
采用大區試驗,試驗面積為900 m2(60 m×15 m),設置3個處理分別為傳統耕作深度20 cm(CK)、粉壟耕作深度35 cm(FL1)、粉壟耕作深度50 cm(FL2)。玉米出苗后將各處理的樣地平均分為4個67.5 m2(15 m×4.5 m)的小區,即為每個處理的4次重復,每個小區之間設1 m寬走道,重復小區間設走道,寬1 m,四周設置保護行,寬3 m。
供試玉米品種為“迪卡5號”,采用吳忠市伊禾農機作業服務有限公司提供的懸掛式粉壟機作業,全面深松垂直旋磨粉碎土壤,粉壟后鎮壓。傳統耕作采用拖拉機犁耙整地。2019年5月3日整地,5月16日播種,10月18日收獲。播種密度為82500株/hm2,肥料為控釋肥0.12 kg/m2(N-P-K:30- 12- 5),作為底肥一次性施入,各處理田間管理均一致。土壤采集時間為10月18日,以五點取樣法在0—20 cm,20—40 cm耕作層取土樣,每個土壤3個平行樣品。將土壤樣品裝入無菌袋中,一部分鮮土過2 mm篩后4℃保存,用于土壤酶活性及Biology-ECO測定,另一部分鮮土過1 mm篩-80℃保存,用于土壤微生物群落結構的測定。
1.3 土壤酶活性的測定
脲酶用靛酚藍比色法測定,以24 h后1 g土壤NH3-N的毫克數表示;土壤堿性磷酸酶采用磷酸苯二鈉比色法,以1 h后1g土壤中對硝基苯酚的毫克數表示;轉化酶用3,5-二硝基水楊酸比色法,以24 h后1 g土壤葡萄糖的毫克數表示[25]。
1.4 土壤微生物功能多樣性的測定
土壤微生物功能多樣性用Biolog方法進行測定[26,27]:稱取5.0 g土壤樣品加入裝有45 mL 0.85%無菌生理鹽水的三角瓶中,25℃ 150 r/min振蕩30 min,冰浴1 min,后靜置30 min,獲得土壤樣品的微生物懸浮液。將土壤懸液梯度稀釋為10-3g/mL,在超凈工作臺中用移液槍將制備好的土壤懸液接種至Biolog微平板的各孔中,每孔150 μL,蓋好蓋子置于25℃培養箱中暗培養7 d,每隔24 h用酶標儀讀取在590 nm與750 nm波長的數值。
Shannon指數(H)用于評估物種豐富度,計算公式如下:
H=-∑(Pi×lnPi)
(1)
式中,Pi為第i孔的相對吸光值與整個平板相對吸光值總和的比率。
McIntosh指數(U)用于評估群落均一度:
(2)
式中,ni為第i孔的相對吸光值。
Simpson指數(D)用于評估常見種優勢度的指數:
D=1-∑Pi2
(3)
式中,Pi為第i孔的相對吸光值與整個平板相對吸光值總和的比率。
1.5 DNA 抽提和 PCR 擴增
各土層FL1、FL2和CK一共18個土壤樣品進行IlluminaMiSeq測序。根據E.Z.N.A.? soil DNA kit (Omega Bio-tek, Norcross, GA, U.S.)說明書進行微生物總DNA抽提,使用1%的瓊脂糖凝膠電泳檢測DNA的提取質量,使用NanoDrop2000測定DNA濃度和純度;細菌引物采用338F (5′-ACTCCTACGGGAGGCA GCAG- 3′)和806R (5′-GGACTACHVGGGTWTCTAAT- 3′),真菌引物采用ITS1F (5′-CTTGGTCATTTAGAGGAA GTAA- 3′)和ITS2R (5′-GCTGCGTTCTTCATCGATGC- 3′),擴增程序如下:95℃預變性3 min,27個循環(95℃ 變性 30s,55℃ 退火30s, 72℃ 延伸30s),然后 72℃ 穩定延伸10min,最后在4℃進行保存(PCR儀:ABI GeneAmp? 9700型)。PCR反應體系為:5×TransStart FastPfu 緩沖液4 μL,2.5mmol·L-1dNTPs 2μL,上游引物(5μmol·L-1)0.8 μL,下游引物(5μmol·L-1)0.8 μL, TransStart FastPfu DNA聚合酶0.4 μL,模板DNA 10 ng,補足至20 μL。
1.6 Illumina Miseq 測序和測序數據處理
將同一樣本的PCR產物混合后使用2%瓊脂糖凝膠回收PCR產物,利用AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA)進行回收產物純化,2%瓊脂糖凝膠電泳檢測,并用QuantusTMFluorometer (Promega, USA)對回收產物進行檢測定量。使用NEXTFLEX Rapid DNA-Seq Kit進行建庫。利用Illumina公司的Miseq PE300平臺進行測序(上海美吉生物醫藥科技有限公司)。
使用fastp[28]軟件對原始測序序列進行質控,使用FLASH[29]軟件進行拼接。使用UPARSE軟件,根據97%[30,31]的相似度對序列進行OTU聚類并剔除嵌合體。利用RDP classifier[32]對每條序列進行物種分類注釋,細菌比對Silva 16S rRNA數據庫,真菌比對UNITE ITS數據庫,設置比對閾值為70%。功能注釋分別基于Tax4Fun和FunGuild用于細菌和真菌。
1.7 數據分析
方差分析(ANOVA)采用SPSS 18.0分析軟件(SPSS Inc.,美國)和Tukey檢驗,在P=0.05顯著性水平上進行。結構方程模型(SEM)用于評估假設的潛在因素,使用IBM SPSS Amos 24進行分析。使用Microsoft Excel 2016,Origin 2016和R軟件繪圖。
2 結果Results
2.1 粉壟耕作對土壤酶活性及玉米產量的影響
表1列出了FL1、FL2和CK處理下不同土壤深度的土壤脲酶、堿性磷酸酶和轉化酶活性。在不同處理下,0—20 cm深度的3種酶活性高于20—40 cm深度。0—20 cm土層中,FL1處理的土壤脲酶活性顯著高于CK處理、堿性磷酸酶和轉化酶活性顯著高于FL2和CK處理。20—40 cm土層中,FL1和FL2處理的脲酶活性比CK處理提高了28.80%和4.45%,堿性磷酸酶活性比CK處理提高了30.95%和16.67%,FL1處理土壤轉化酶比CK處理提高了25.61%,但各處理間差異均沒達到顯著水平(P>0.05)。FL1、FL2處理產量分別為8.58 t/hm2和 8.38 t/hm2,與對照處理相比提高了37.9%和34.7%(P<0.05)。
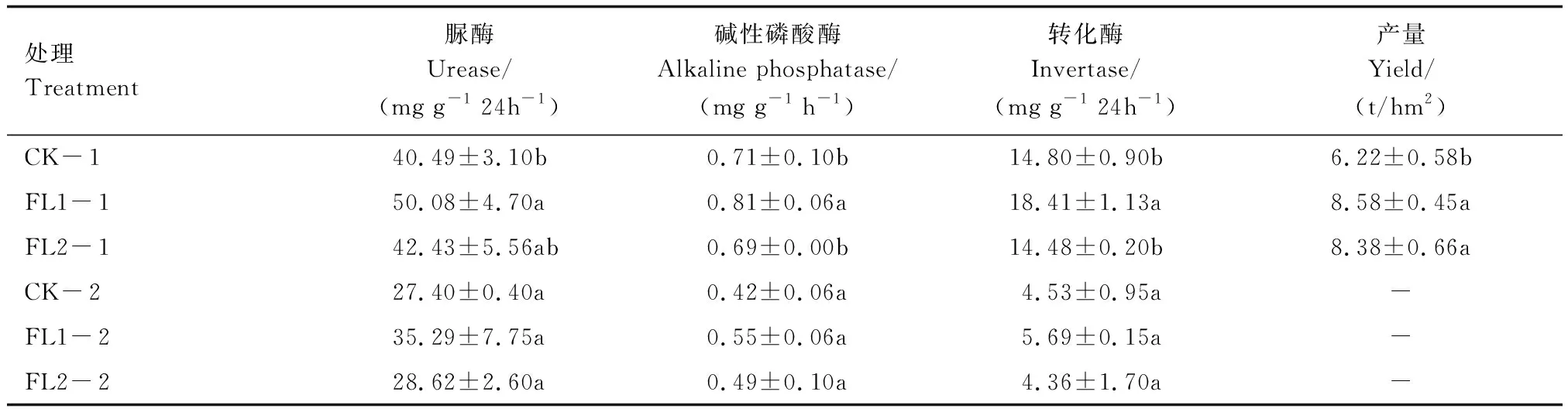
表1 不同土層處理間土壤酶活性Table 1 Soil enzyme activity between treatments in different soil layers
2.2 粉壟耕作對微生物群落組成的影響
本研究中,通過高通量測序技術檢測所有土壤樣品的微生物,獲得了30730個OTUs,包括26668個細菌OTUs和4062個真菌OTUs。對各處理平均豐度大于1的所有土壤細菌和真菌OTUs進行了Flower plot分析。對于土壤細菌OTUs,在0—20 cm和20—40 cm的FL1、FL2和CK處理共有2296個相同OTUs。0—20 cm土層中FL1、FL2和CK處理特有細菌OTUs分別是81、206和72個,20—40 cm土層中FL1、FL2和CK處理特有細菌OTUs分別是135、83和194個(圖1)。對于土壤真菌OTUs,在0—20 cm和20—40 cm的FL1、FL2和CK處理共有211個相同OTUs。0—20 cm土層中FL1、FL2和CK處理特有真菌OTUs分別是129、85和102個,20—40 cm土層中FL1、FL2和CK處理特有真菌OTUs分別是67、129和88個(圖1)。在門水平上分析了樣本中的細菌和真菌分布(圖1)。不同土層各樣本中的細菌在門水平排名前10的物種組成相同(圖1)。不同土層各處理間優勢菌門為變形桿菌、放線菌門、綠彎菌門和酸桿菌門,達78.85%—81.83%。0—20 cm土層中的FL1、FL2和CK處理的細菌門差異不顯著(P>0.05)。
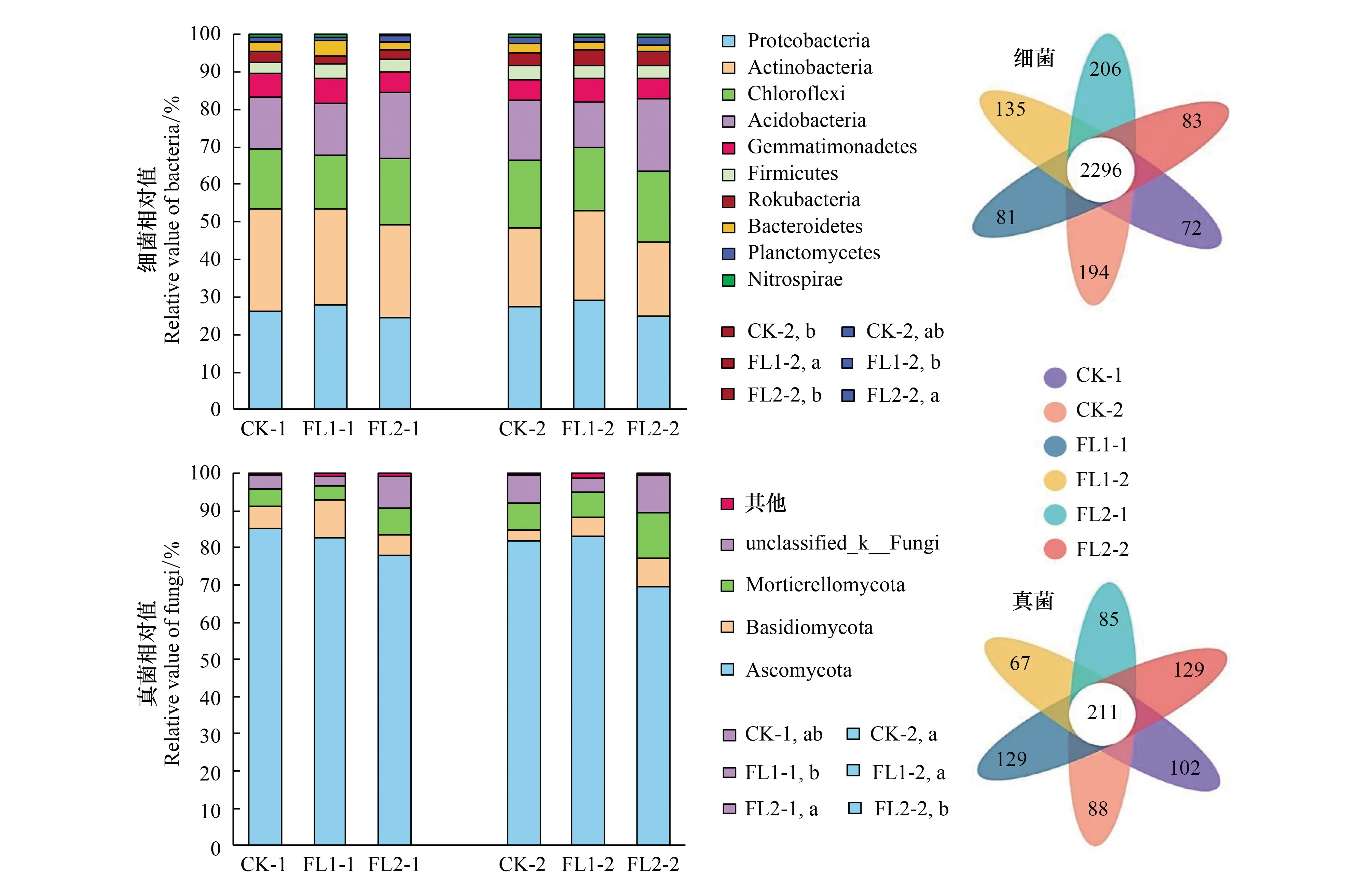
圖1 不同處理下土壤微生物群落組成及花瓣圖Fig.1 Soil microbial community composition and flower plot under different treatments細菌相對值:Proteobacteria 變形桿菌;Actinobacteria放線菌門;Chloroflexi 綠彎菌門;Acidobacteria 酸桿菌門;Gemmatimonadetes 芽單胞菌門;Firmicutes 厚壁菌門;Bacteroidetes 擬桿菌門;Planctomycetes 浮霉菌門;Nitrospirae 硝化螺旋菌門;真菌相對值:Ascomycota 子囊菌門; Basidiomycota 擔子菌門; Mortierellomycota 被孢霉門; unclassified_k__Fungi 未分類_k__真菌
20—40 cm土層中,FL1處理的Rokubacteria與FL2和CK處理差異顯著(P<0.05),FL2的變形桿菌門與FL1處理差異顯著(P<0.05)。在真菌在不同土層各樣本中,對豐度小于1%的物種歸類為others。0—20 cm土層中,FL1處理的unclassified_k__Fungi與FL2處理差異顯著(P<0.05)。20—40 cm土層中,FL1處理的子囊菌門與FL2處理差異顯著(P<0.05)(圖1)。
在這些OTUs的基礎上,進一步分析了物種豐富度指數和多樣性指數(表2)。總的來說,在0—20 cm土層中,FL2與CK處理的細菌Ace、Chao和香農指數差異顯著(P<0.05);FL1與CK處理的真菌Ace指數、Chao指數和辛普森指數差異顯著(P<0.05),FL1與FL2處理間Ace指數、Chao指數、香農指數和辛普森指數差異顯著(P<0.05)。20—40 cm土層中,FL1與FL2處理的細菌香農指數差異顯著(P<0.05),FL2與CK、FL1處理的細菌辛普森指數差異顯著(P<0.05);FL1與CK處理間真菌辛普森指數差異顯著(P<0.05)。
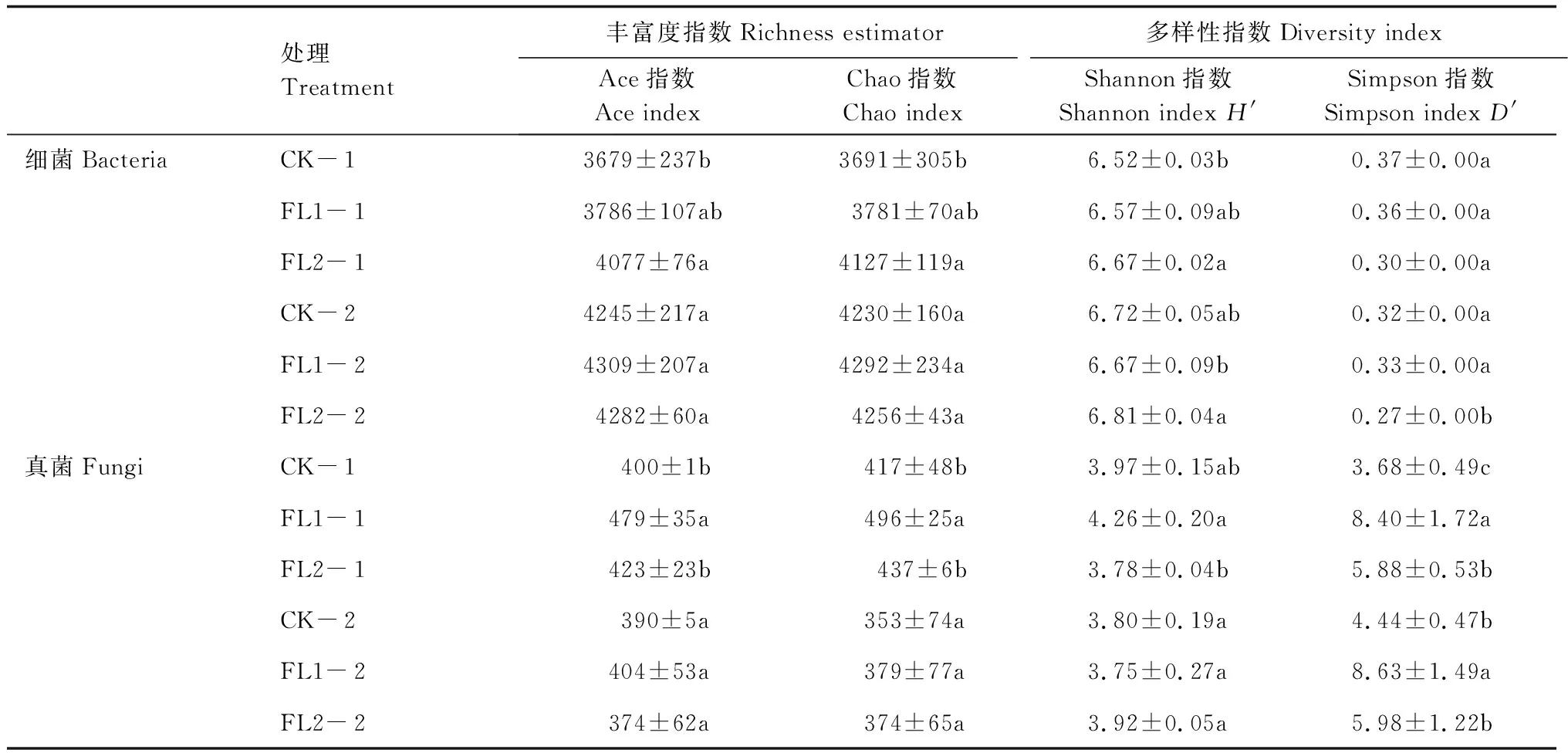
表2 不同土層處理間土壤細菌群落豐富度和群落多樣性指數Table 2 Soil bacterial community richness and community diversity index between treatments in different soil layers
2.3 粉壟耕作下土壤微生物群落功能多樣性
本研究中以培養96 h獲得的數據對土壤微生物功能多樣性指數進行分析(表3)。0—20 cm土層中,FL1處理的香農指數與FL2處理相比差異顯著(P<0.05),麥金托什指數中,FL1處理與FL2、CK處理相比差異顯著(P<0.05)。20—40 cm土層中,FL2處理香農指數與FL1、CK處理相比差異顯著(P<0.05),辛普森指數FL2與CK處理之間差異顯著(P<0.05)。

表3 不同土層處理間土壤微生物功能多樣性Table 3 Soil microbial functional diversity between treatments in different soil layers
對不同土層各處理間土壤微生物群落的碳源利用情況進行分析時發現,從土壤微生物群落利用百分比來看,以利用羧酸類、聚合物類、氨基酸類以及糖類為主,胺類和雙親化合物類的利用較低(圖2)。在0—20 cm土層中,FL1、FL2和CK處理利用羧酸類碳源差異顯著(P<0.05)。在20—40 cm土層中,FL1與FL2處理對胺類碳源的利用差異顯著(P<0.05)(圖2)。
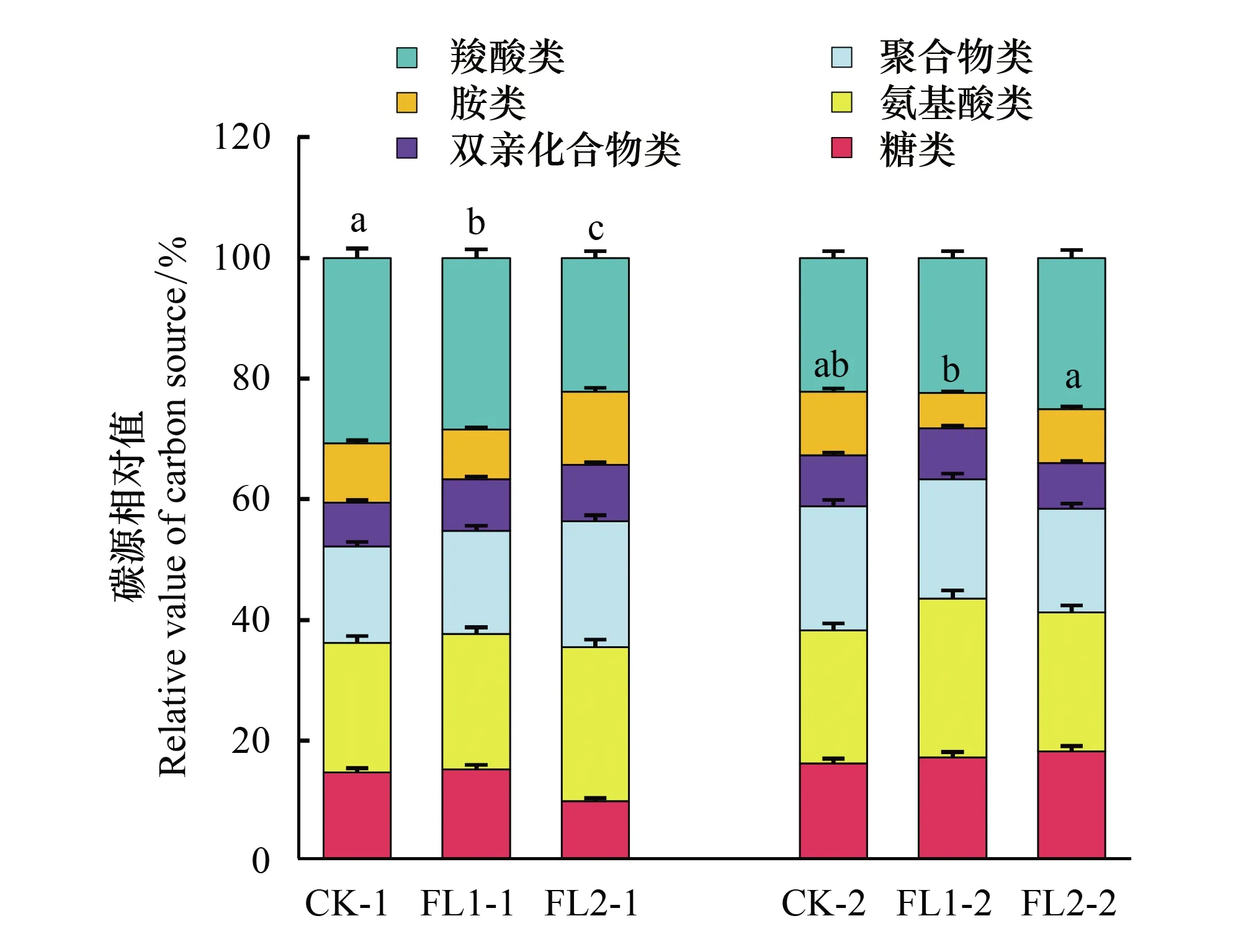
圖2 不同處理下土壤微生物對6種碳源的相對利用 Fig.2 The relative utilization of soil microorganisms to 6 carbon sources under different treatments不同小寫表示不同處理間差異顯著(P<0.05)
主成分分析結果表明,PC1和PC2的貢獻率分別為27.7%和16.6%。影響PC1的碳源主要有糖類、氨基酸類、羧酸類和聚合物類;影響PC2的碳源主要有羧酸類和糖類。0—20 cm土層中,FL1和CK處理位于PC2正端,受PC2上的碳源影響較大,相似度較高,碳源利用差異較小;FL2處理在第三象限,與FL1和CK距離過大,碳源利用差異較大(圖3)。20—40 cm土層中,CK位于第四象限,且FL1、FL2和CK處理相對分離,說明各處理間碳源利用有差異(圖3)。
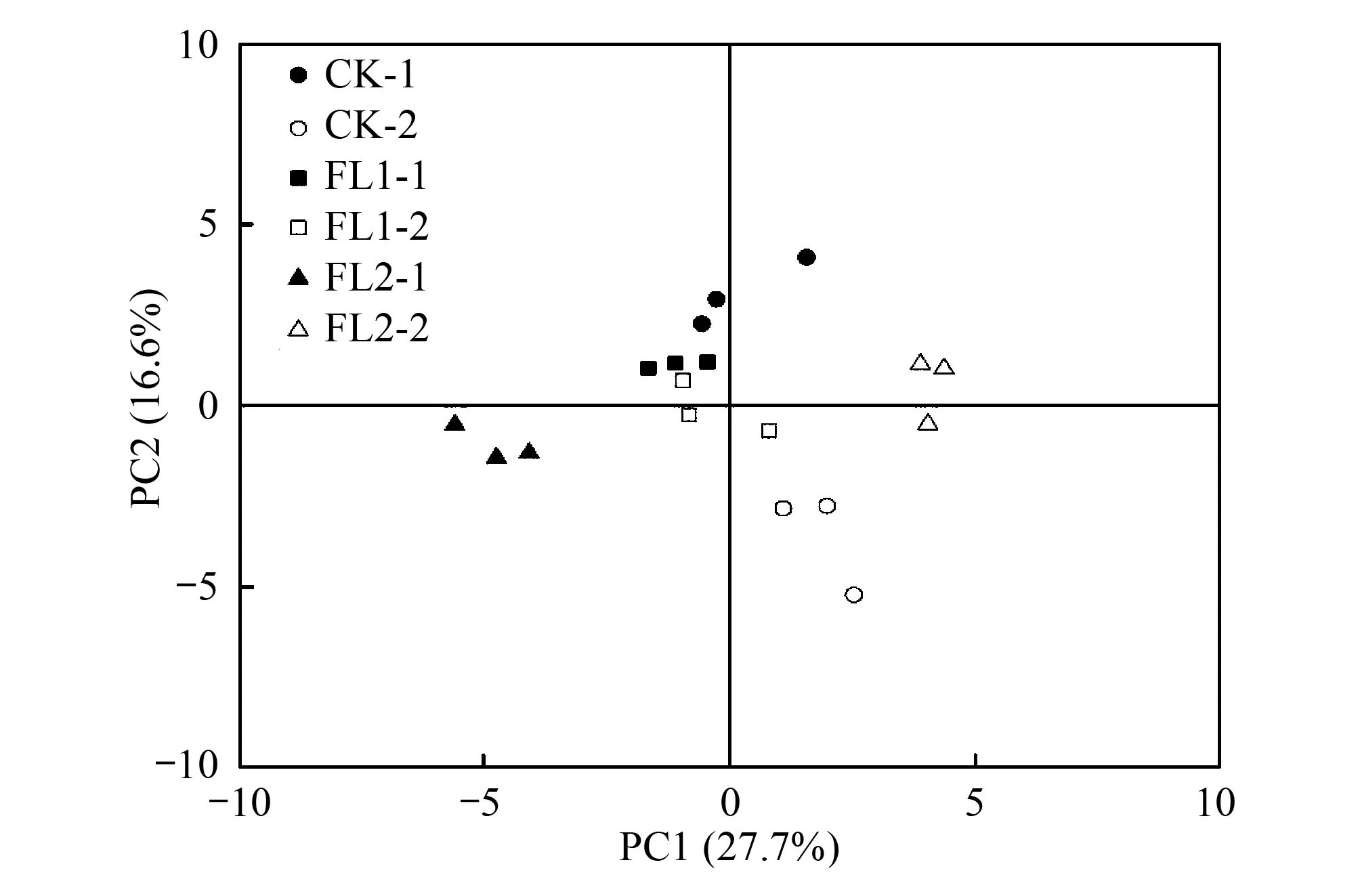
圖3 不同處理下土壤微生物群落碳源利用主成分分析 Fig.3 Principal component analysis of carbon source utilization of soil microbial communities under different treatments
2.4 粉壟耕作下土壤細菌和真菌的功能預測
使用Tax4Fun進行潛在的微生物KEGG功能預測,代謝作用和環境信息處理是主要一級代謝通路,平均占比分別為63.67%和17.29%(圖4)。在二級代謝通路上總共注釋了41個特征,圖5顯示了前15個細菌的相對功能基團豐度。在0—20 cm和20—40 cm土層中,FL1、FL2和CK處理的主要細菌功能基團的功能注釋為:氨基酸代謝、碳水化合物代謝、膜運輸、信號傳遞、萜類化合物和聚酮化合物的代謝、能量代謝(圖4)。FL1、FL2和CK處理在0—20 cm土層的輔因子和維生素的代謝差異顯著(P<0.05)。在20—40 cm土層中,FL1、FL2的碳水化合物代謝顯著高于CK處理(P<0.05),FL1的氨基酸代謝、輔因子和維生素的代謝和膜運輸顯著高于CK處理(P<0.05),FL2的信號傳遞、能量代謝顯著高于CK處理(P<0.05)。

圖4 不同處理下不同土層主要土壤細菌的功能結構(二級代謝通路)Fig.4 Functional structure of main soil bacteria in different soil layers under different treatments (Pathway level 2)
使用FUNGuild對真菌進行功能Guild注釋(圖5)。按真菌的營養類型主要分為9種功能類型:病原-腐生-共生菌、腐生菌、病菌、腐生-共生菌、病原-腐生-共生菌、病菌-腐生菌、共生菌和病菌-共生菌。在0—20 cm土層中,FL1的病菌-腐生菌、病菌-共生菌、腐生菌和腐生-共生菌顯著高于CK(P<0.05),FL2的共生菌和腐生-共生菌顯著高于CK(P<0.05)。在20—40 cm土層中,FL1的病菌-腐生菌-共生菌顯著高于CK(P<0.05),FL2的病菌-腐生菌和共生菌顯著高于CK(P<0.05)(圖5)。
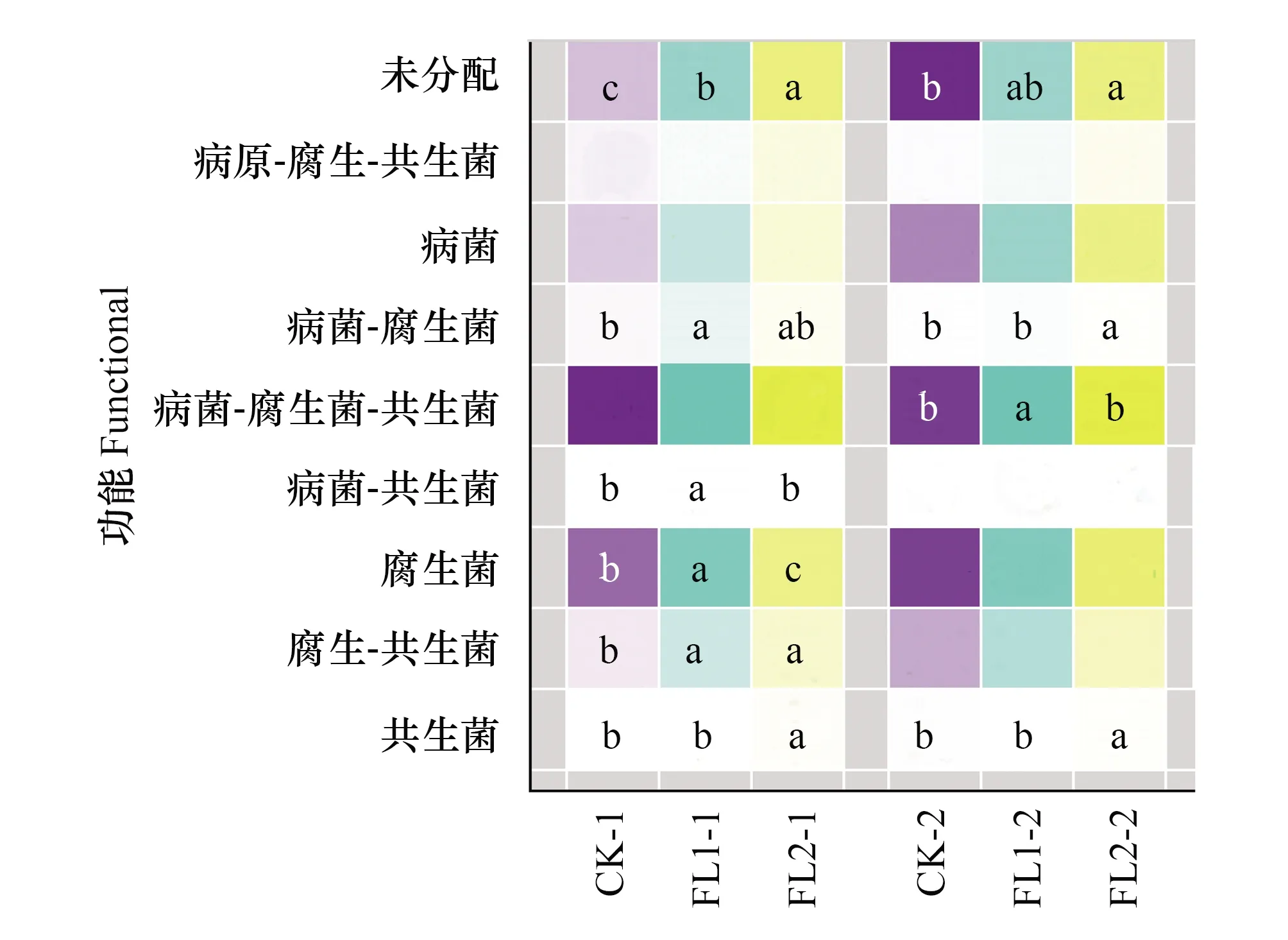
圖5 不同處理下不同土層主要土壤真菌的功能結構(二級代謝通路)Fig.5 Functional structure of main soil fungi in different soil layers under different treatments (Pathway level 2)
2.5 影響微生物功能多樣性的生態因素及相關性分析
結構方程模型(SEM)檢測控制因素對土壤微生物功能多樣性有直接和間接的影響(圖6)。耕作方式的直接效應能解釋土壤細菌群落多樣性總方差的72%(圖6)。土壤細菌群落多樣性能夠解釋土壤真菌群落多樣性總方差的11%(圖6)。耕作方式和土壤微生物(細菌、真菌)群落多樣性能解釋土壤酶活性總方差的96% (圖6)。耕作方式、土壤微生物群落多樣性和土壤酶活性能解釋土壤微生物功能多樣性總方差的69% (圖6)。
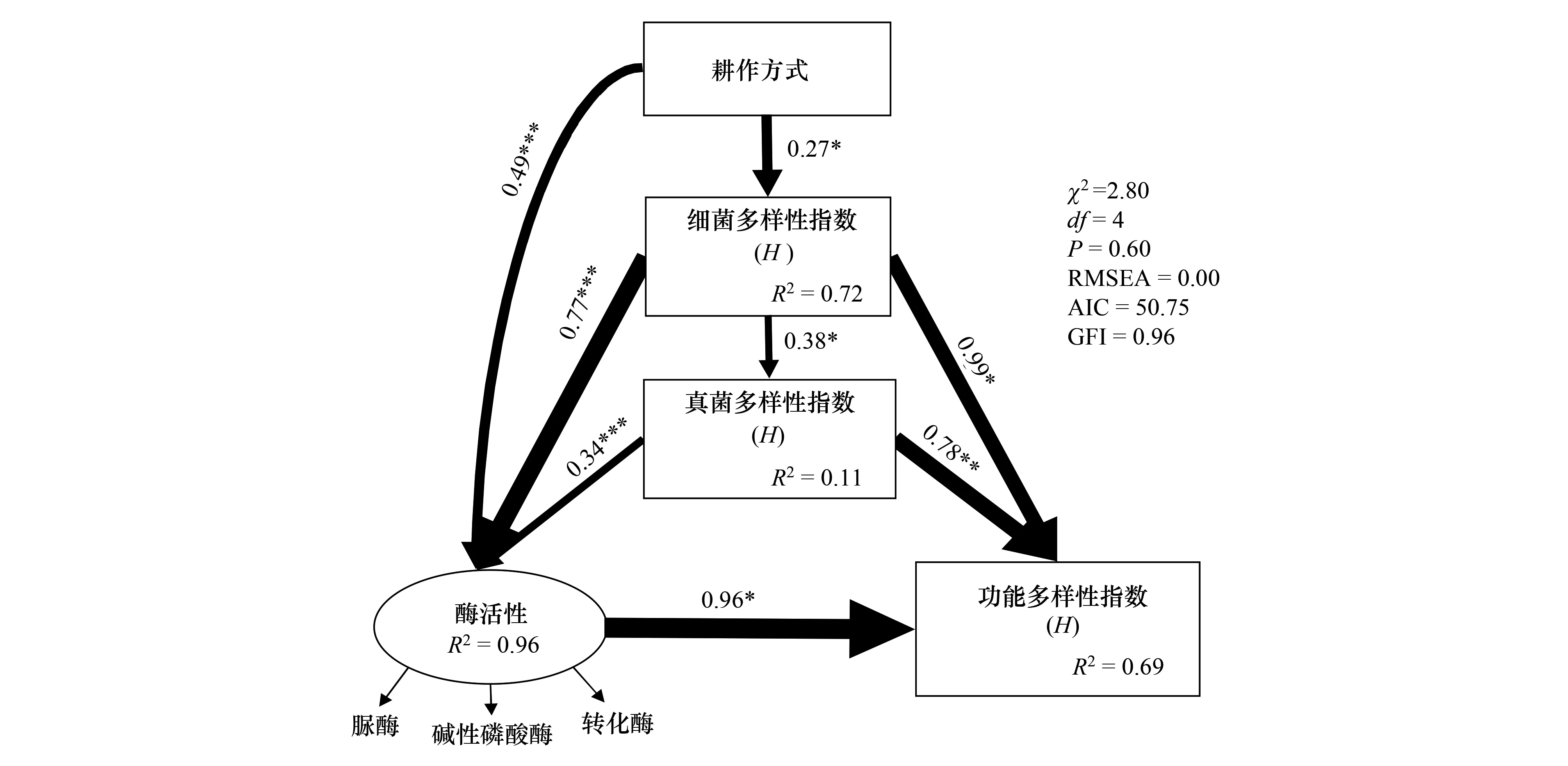
圖6 耕作方法、土壤酶活性、細菌和真菌多樣性指數和土壤微生物功能多樣性的結構方程模型(SEM)Fig.6 Structural equation model (SEM) of farming methods, soil enzyme activity, bacterial and fungal diversity index and soil microbial functional diversity箭頭的粗細表示標準化路徑系數的大小;R2 值代表每個內生變量解釋方差的比例;GFI:擬合優度指數;RMSEA:近似的均方根誤差;*P < 0.05, **P < 0.01, ***P < 0.001
如表4所示,玉米產量與土壤脲酶、轉化酶、土壤微生物香農指數(H)、麥金托什指數(U)和細菌群落香農指數(H′)顯著正相關(P<0.05) ; 三種土壤酶(Ure、Inv、Alp)均與土壤微生物辛普森指數(D)、細菌群落多樣性指數(Ace、Chao、H′、D′)和真菌群落多樣性指數(Ace、Chao)顯著正相關(P<0.05);土壤微生物香農指數(H)與細菌群落多樣性指數(Ace、Chao、H′、D′)和真菌群落多樣性指數(H′、D′)顯著正相關(P<0.05)。
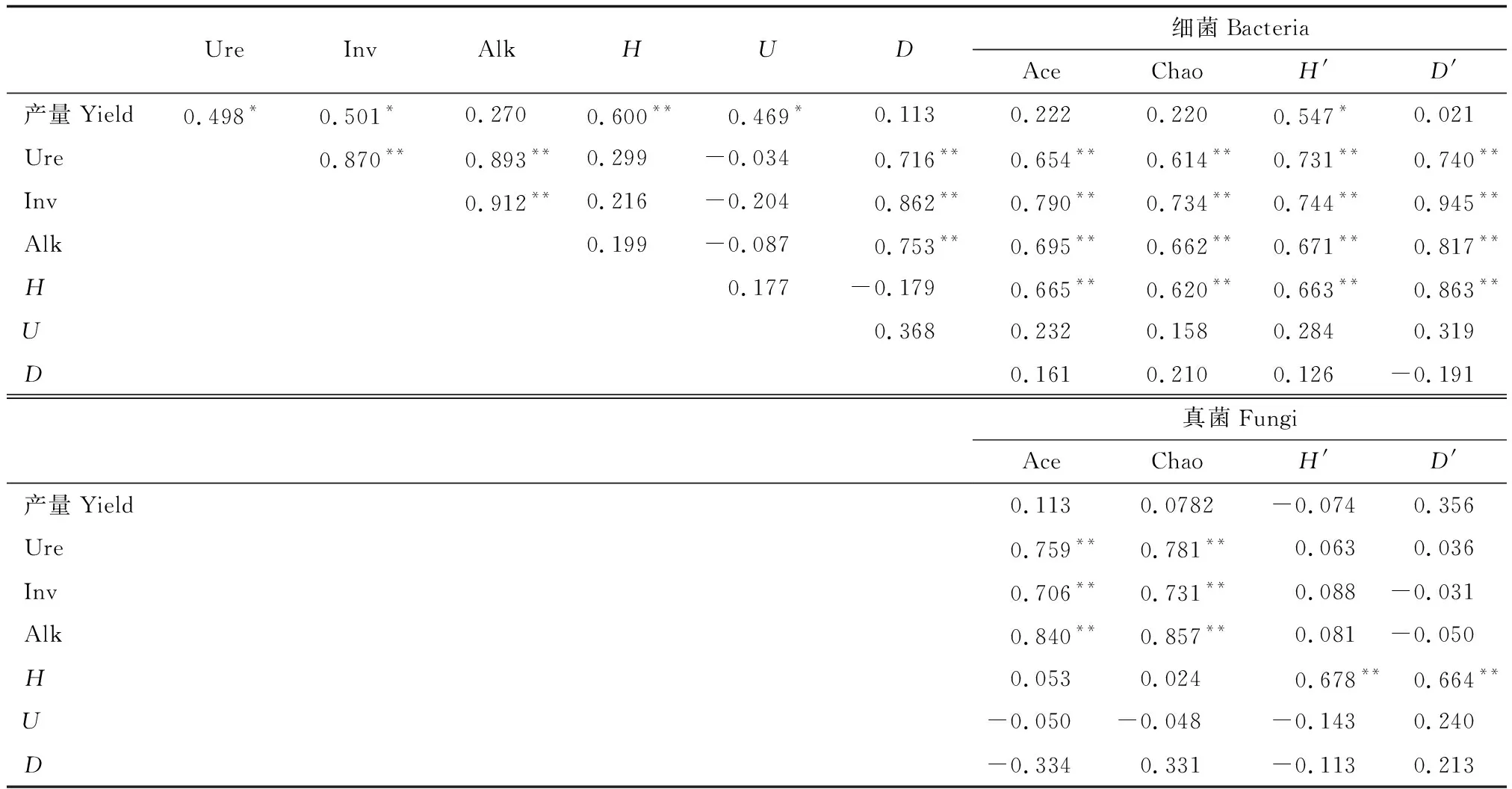
表4 土壤酶活性、微生物群落與微生物功能多樣性以及產量相關性分析Table 4 Correlation analysis of soil enzyme activity, microbial community and microbial functional diversity and yield
3 討論
3.1 粉壟耕作對土壤微生物的影響
土壤微生物群落活動和組成受到農業耕作方式影響[33—35]。粉壟耕作較常規耕作有效提高土壤微生物數量,使土壤中的氨化細菌、固氮菌、無機磷細菌和鉀細菌的數量高于旋耕耕作處理和深翻耕作處理[24]。楊博[36]研究發現,粉壟耕作為深層土壤微生物的生存提供了有利條件,0—20 cm和20—40 cm土層土壤微生物總菌落數較傳統耕作顯著提升。本研究中,細菌的OTUs數量明顯高于真菌的OTUs,表明土壤細菌占土壤微生物總數量的比例較大。本研究發現,粉壟耕作與傳統耕作相比改變了細菌的多樣性和群落組成(圖1,表2)。
造成的原因可能是,粉壟耕作提高了耕地土壤耕層碳存儲,降低了土壤緊實度,促進了土壤大團聚體的形成[37,38]。土壤團聚結構的動態變化又反饋控制土壤微生物活動和土壤有機質組分的分異作用,使土壤有機質組分-土壤團聚結構-微生物群落結構的動態變化之間存在耦合作用[39],從而改變了細菌群落結構和多樣性。此外,由于細菌提供信號分子或將介質化合物轉化為真菌消耗的物質促進了真菌的生長,提高了真菌的定殖率,甚至促進真菌的完整生命周期[40—42]。這些結果表明,真菌群落間接地受到了粉壟耕作方式的影響,導致真菌群落結構、物種豐富度指數和多樣性指數較傳統耕作有所提升(圖1,表2)。粉壟耕作提升了耕地土壤的微生物群落多樣性,使參與土壤養分轉化的功能型微生物增加,為維系耕地土壤微生態環境的穩定發揮了重要作用。
3.2 控制因素對土壤酶活性的影響
研究發現,耕作方式和土壤酶活性顯著正相關(圖6)。土壤中的氮、磷轉化以及碳水化合物的代謝和運輸貯藏與脲酶、轉化酶和堿性磷酸酶密切相關[43,44]。作物根系可以產生酶進行養分吸收[45],粉壟耕作通過物理方式改善作物土壤環境,使作物根系具有活力,向下伸展[46],釋放出更多的酶來加強與土壤微生境養分的互換[24],使粉壟耕作比傳統耕作具有更高的土壤酶活性。土壤中的酶不僅來源于作物根系,還來源于土壤微生物的代謝活動[47]。異養型細菌為了自身發展,會釋放出相應功能的酶,來參與土壤養分的循環[48]。王丹等[49]研究發現,放線菌門(Actinobacteriota)可以降解土壤中各種不溶性有機物質以供細胞代謝所需的各種營養,與水解酶的活性有顯著相關。擬桿菌門(Bacteroidota)是具有溶磷作用的富營養菌,參與養分代謝,與脲酶、轉化酶和堿性磷酸酶呈正相關[50]。真菌群落的發展使養分循環加快,能量代謝有關的真菌和參與養分循環土壤酶高度相關[51],也進一步證明土壤酶活性的高低受土壤真菌的影響。傳統耕作不利于作物表層根系發育和土壤微生物活動[52],導致土壤中酶活性低。粉壟耕作通過改善耕地土壤物理結構,促進了作物根系的生長和土壤微生物自身發展,并釋放出相應的酶,加快了土壤養分的循環,使粉壟耕作下的土壤酶活性高于傳統耕作(表1),并且脲酶和轉化酶活性與產量顯著正相關(表4),粉壟耕作處理的玉米產量得到提升。
3.3 控制因素對功能多樣性的影響
已有研究表明,根系的生長可以釋放酶和碳源[53],從而影響分解碳源的微生物,有利于土壤微生物多樣性的增加[54,55]。粉壟耕作與傳統耕作之間微生物群落結構存在差異,粉壟耕作深度35 cm顯著提升了0—20 cm土層的功能多樣性,粉壟耕作深度50 cm顯著提升了20—40 cm土層微生物功能多樣性。造成的原因可能是,粉壟耕作可以促進根系生長,使根系釋放出更多的碳源,從而提高了微生物功能多樣性。有研究表明,患病根系增加了羧酸類和胺類碳源的釋放,從而提升了土壤微生物對羧酸類和胺類碳源的利用[56,57]。本研究得出,在0—20 cm土層中粉壟耕作對羧酸類碳源的利用率顯著降低;在20—40 cm土層中,粉壟耕作深度35 cm和粉壟耕作深度50 cm相比顯著降低了對胺類碳源的利用比例。說明粉壟耕作可以減少作物根系患病的幾率,患病根系向周圍分泌羧酸類和胺類碳源減弱,使微生物利用碳源的能力發生改變。
土壤微生物細菌和真菌群落多樣性可以直接影響微生物功能多樣性(圖6)。已有研究表明,土壤中微生物群落結構和碳源代謝緊密相關[58]。土壤中擁有代謝、遺傳信息處理、有機系統3類功能基因的共養生物(copiotrophs、R-strategists)和寡養生物(oligotrophs、K-strategists)相互作用,促進了細菌群落從土壤中吸收營養物質和溶解鐵和小分子等來加速自身生長,使土壤細菌代謝旺盛,提高了細菌功能群落多樣性[59—61]。真菌功能預測的腐生營養型為最主要的營養型,這可能與子囊菌門為優勢菌門有關。子囊菌門大多為腐生菌,是土壤中重要的分解者[62],可以分解難降解的有機質,在養分循環方面起著重要作用[63]。另外,含有共生營養型的叢枝菌根真菌(AMF)與宿主植物建立共生關系后, 不僅顯著影響植物生長, 還能引起根系分泌物的變化[64,65]。AMF與植物共生可以影響土壤細菌群落結構和功能多樣性[66]。本研究得出,粉壟耕作下細菌功能預測的一級代謝通路代謝作用和環境信息處理比傳統耕作顯著增加(圖4),真菌的功能預測出的營養類型也比傳統耕作顯著提高(圖5)。原因可能是,粉壟耕作可以增加了細菌的功能基團,細菌增強了養分的攝取能力,功能代謝得到提升。真菌不同營養類型的功能基團增加,提升了自己參與碳源代謝的能力。真菌不僅可以分解出細菌利用的碳源,而且間接促進根系向土壤釋放碳源[67]。細菌真菌相互作用,使土壤中代謝底物變得豐富多樣,土壤微生物功能多樣性提升。此外,粉壟耕作下的病菌-腐生菌、病菌-共生菌、腐生菌、腐生-共生菌和病菌-腐生菌-共生菌等功能基團不同程度下均高于傳統耕作處理,可能的原因是,粉壟耕作處理顯著改變了真菌群落結構,真菌數量增多,導致相應功能的功能基團也增多。
土壤酶活性作為微生物活性的指示物,與微生物功能多樣性顯著相關[68—71]。段益莉等[72]研究表明,土壤酶活性的高低不僅可以反映出有機物質的水解程度,還能反映出對碳源利用能力的高低。本研究也發現,土壤酶活性顯著影響了微生物功能多樣性(圖6),表明粉壟耕作可以通過提升酶活性間接的使土壤微生物功能多樣性提高,使土壤微生物群落利用養分的能力加強,群落功能結構完善,增加了土壤生態系統的穩定性,提升了玉米的產量。
4 結論
本文研究了粉壟耕作對玉米耕地土壤酶活性、微生物群落結構和功能代謝的影響,研究表明:粉壟耕作可以顯著改善土壤細菌、真菌群落結構,增加參與養分代謝的功能基團,提高土壤微生物群落多樣性,并增加了玉米產量;粉壟耕作提升了參與養分循環的土壤脲酶、轉化酶和堿性磷酸酶的活性;粉壟耕作技術促進了土壤微生物養分代謝的能力,使土壤微生物總體的功能多樣性得到提升;從土壤微生物群落多樣性、功能多樣性、酶活性和玉米產量可以看出,粉壟耕作深度35 cm處理優于粉壟耕作深度50 cm。總的來說,從土壤微生態的角度可以初步得出粉壟耕作技術可以促進了養分循環,使土壤耕地質量得到提升,增加了農作物的產量。本研究為粉壟耕作技術在中國西部干旱半干旱地區作物栽培的應用發展提供了科學依據。
耕作方式對耕地質量的影響是一個周期較長的過程。因此,隨著長期粉壟耕作的使用,耕地物理結構和土壤微生態也會發生著變化。本團隊僅研究了一年的粉壟耕作不同深度對耕地土壤酶活性、微生物群落結構和功能多樣性的影響,未來會在此試驗田進行長期工作,今后會對土壤微生態的變化進行長期檢測和跟蹤研究,并進一步增加關于土壤團聚體、土壤碳含量以及土壤病害等研究內容。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
