中華菊頭蝠的棲息生態特征及種群保護
2022-08-03 02:53:18伍麗艷任伯凇黃太福龔小燕劉志霄
生態學報 2022年12期
伍麗艷,任伯凇,吳 濤,黃太福,龔小燕,劉志霄
吉首大學生物資源與環境科學學院,吉首 416000
巖溶洞穴簡稱溶洞或洞穴,是地球表層可溶性巖石在地表水和地下水的溶蝕及侵蝕作用下所形成的地下空間,我國喀斯特地貌分布十分廣泛,溶洞資源非常豐富,為洞棲性蝙蝠的棲息和繁衍提供了優越的生態條件[1—4]。
棲息地選擇是種群生態學研究的重要內容,已逐漸成為蝙蝠生態研究的熱點[5—9]。洞棲性蝙蝠既是翼手目動物的主要生態類群之一,也是洞穴生態系統中最重要的組成部分,其在溶洞中的棲息生態特征與其自身的生存繁衍及洞穴生態系統的物質循環、能量流動與信息傳遞密切相關,具有重要的生態生物學與保護生物學研究意義[10—14]。
隨著城市的發展,蝙蝠的棲息地遭到了嚴重的破壞,可供蝙蝠選擇的棲息場所日趨減少,使得蝙蝠的數量也急劇減少,目前約有51%的蝙蝠物種正遭受著威脅[15]。
湘西土家族苗族自治州(以下簡稱湘西州)境內氣候溫暖濕潤,喀斯特地貌發育良好,溶洞眾多,已知有大蹄蝠(Hipposiderosarmiger)、西南鼠耳蝠(Myotisaltarium)和菊頭蝠(Rhinolophusspp.)等洞棲食蟲性蝙蝠20多種,它們在自然生態平衡中發揮著非常重要的作用[4, 14]。可是,已有的文獻[16—17]及近年的調查表明,不同的洞穴中所棲息的菊頭蝠的種類及數量大不相同,而更多的洞穴內根本沒有菊頭蝠棲息,并且菊頭蝠棲息的洞穴環境不斷地遭受著人類活動的干擾與破壞,許多洞穴中菊頭蝠的種群數量在急劇下降,甚至已經絕跡。因此,菊頭蝠的棲息生態研究與物種就地保護具有十分重要的意義。
中華菊頭蝠是典型的洞棲性蝠種[18],廣泛分布于我國境內,主要分布于長江以南的省區,也見于老撾、馬來西亞、越南等周邊國家[19—20]。當前,對中華菊頭蝠的研究包括形態、核型、回聲定位、系統進化、基因組學及其病源性等方面[21—31],但有關其在溶洞中的棲息生態特征還缺乏專門的研究報道。
鑒于中華菊頭蝠分布的廣泛性及其在洞穴生態系統中的典型性與代表性,近年我們結合湘西州溶洞資源豐富的生態環境特點[4,32],對其棲息生態特征進行了較為系統的野外觀察,旨在豐富該物種的生態生物學資料,并為洞棲性蝙蝠的種群保護提供參考依據。
1 研究區域與方法
1.1 區域簡況
湘西州(109°10′—110°22.5′E,27°44.5′—29°38′N)地處我國第二階梯向第三階梯的過渡地帶,位于武陵山脈東部的湖南省西北部,西接云貴高原、北鄰鄂西山地,大體上處于我國東洋界華中區、華南區、西南區三大動物地理區的交匯區域,總面積約1.5萬km2。境內屬亞熱帶季風濕潤性氣候,降雨量豐沛,加之地貌結構多樣,山勢陡峻,溝壑縱橫交錯,因此河流眾多,溶洞發育良好,洞穴資源較為豐富[32],為洞棲性蝙蝠的棲息及種群繁衍提供了良好的自然生態條件。
1.2 研究方法
1.2.1物種識別
在湘西州域內,中華菊頭蝠較為常見[4],常與皮氏菊頭蝠(Rhinolophuspearsonii)、菲(小)菊頭蝠(Rhinolophuspusillus)、大菊頭蝠(Rhinolophusluctus)、西南鼠耳蝠(Myotisaltarium)、大蹄蝠(Hipposiderosarmiger)、中華鼠耳蝠(Myotischinensis)等蝠種棲居于同一洞穴。但中華菊頭蝠體型中等偏小,前臂長約44.6—51.5 mm,其鼻葉較寬,馬蹄葉較大,頂葉呈三角形且下緣凹陷,連接葉寬而平滑,下唇有三個缺口,毛色黃棕色或棕紅色,易與其他蝠種區分。
1.2.2數據采集與分析
按照項目組前期的研究方法[33—35]進行野外數據采集與室內數據分析。主要利用T391+激光測距儀、標智GM700手持式激光測溫儀、尼康D7200相機以及高性能手機等工具測量及拍攝記錄蝙蝠的棲點位置、體溫、棲點溫度,以及棲息姿勢等棲息生態學數據,同時對于洞內其他生物類群的分布情況與生態關系也進行一些觀察與記錄。此外,我們還在該蝠集中分布的區域布放紅外相機或便攜式主動紅外錄像裝置以監拍其活動情況[14,33—35]。
本文結合實際,主要根據主洞道的長度及整個洞穴的復雜程度將溶洞分為兩類:Ⅰ類:主洞道長度在500 m之內(包括500 m),洞道結構相對簡單;Ⅱ類:主洞道長度大于500 m,洞內結構較為復雜,岔道較多或洞道分層。
根據實地調查,結合對洞穴附近村民的訪問調查,我們將洞穴的受干擾程度分為高、中、低三個等級:“高”,溶洞被強度開發過,洞內及洞口附近的人工設施及廢棄物較多,經常有人進洞游玩或砸取鐘乳石;“中”,溶洞在路邊或村莊附近,洞口周圍的植被遭到明顯破壞,偶有人進洞活動;“低”,溶洞位置較為偏僻,洞口周圍的植被相對呈自然狀態,多年來幾乎無人進洞活動。
在綜合分析洞道長度和蝙蝠棲點分布密度數據的基礎上,考慮到該蝠種在非冬眠期的分布位置較為多樣,為便于數據分析,我們將每個洞穴的洞道劃分成間距為50 m的區段,共劃分為10個區段:A(0—50 m)、B(50—100 m)、C(100—150 m)、D(150—200 m)、E(200—250 m)、F(250—300 m)、G(300—350 m)、H(350—400 m)、I(400—450 m)、J(450—500 m)。由于普通人伸手及踮腳時的一般高度為2 m左右,所以在蝙蝠棲點高度分布的分析中,我們以2m為高度間隔,將蝙蝠棲點的離地高度劃分為8個區段:①(0—2 m)、②(2—4 m)、③(4—6 m)、④(6—8 m)、⑤(8—10 m)、⑥(10—12 m)、⑦(12—14 m)、⑧(14—16 m)。根據棲點的隱蔽性及人為因素,將棲點的安全性分為 3 級:“高”,棲點位置高,人類難于直接或借助石頭,或手持拐杖、短木棍等工具傷害蝙蝠,或者位置雖較低,但藏得很隱蔽,不易被發現;“中”,棲點位置較高,利用上述工具可以較易打擊到蝙蝠;“低”,蝙蝠的棲點位置低,隱蔽性差,人類極易接觸到它和對其構成傷害。
采用Excel、SPSS 22.0、GraphPad 8.0軟件進行數據分析,利用獨立樣本t檢驗和單因素方差分析對數據進行顯著性檢驗。
2 結果與分析
2.1 種群數量與分布
調查期間,共記錄到中華菊頭蝠12942只次,它們分布于16個溶洞,海拔介于248—1083 m,其中I類洞穴的數量約占洞穴總數的68.75%,II類洞穴占31.25%。中華菊頭蝠主要分布于海拔330—380 m的溶洞中,其他海拔的溶洞中也有分布但相對較少(表1)。
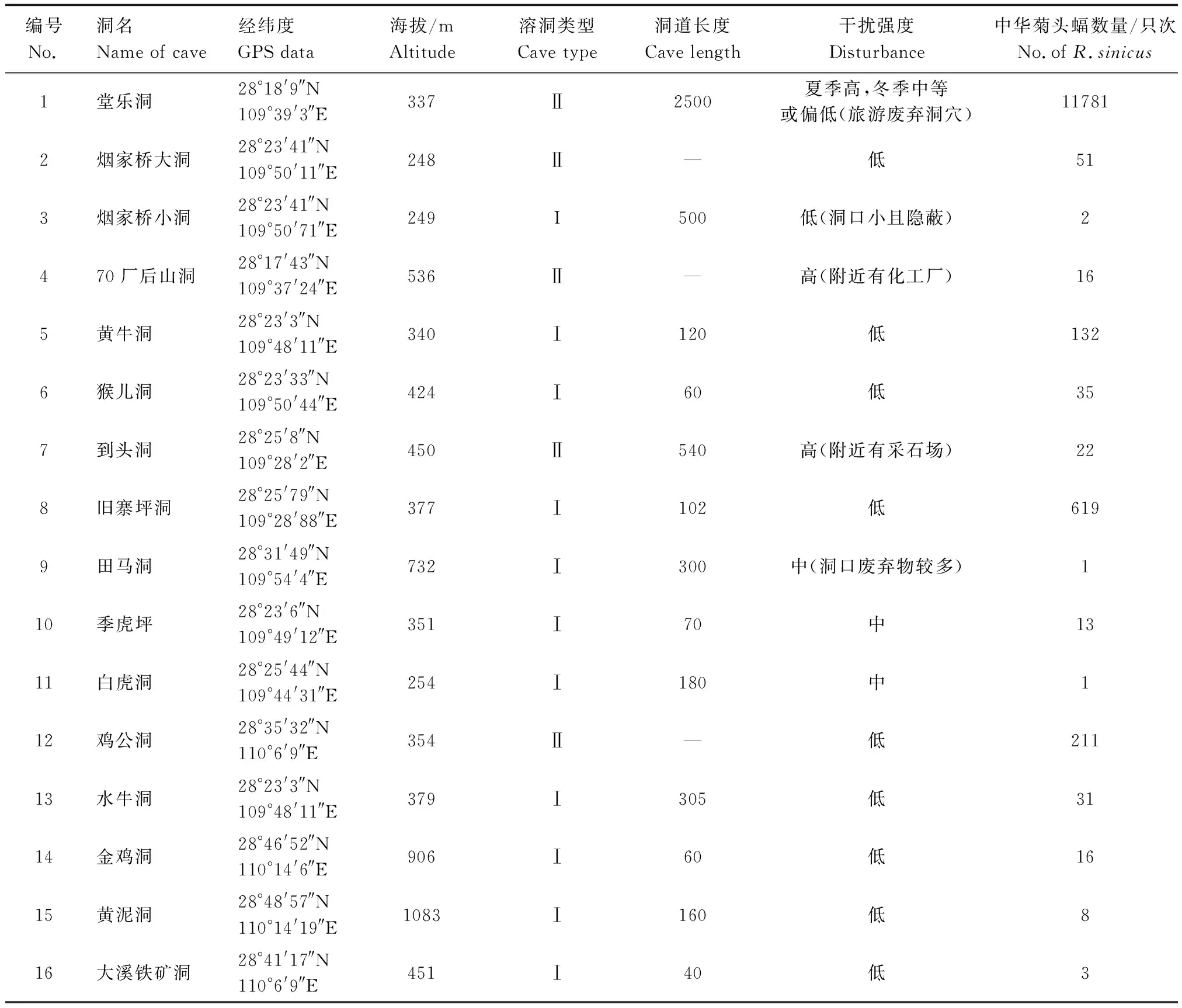
表1 調查期間發現有中華菊頭蝠(Rhinolophus sinicus)棲息的16個溶洞的基本情況Table 1 Basic information of 16 Karst caves inhabited by Rhinolophus sinicus during investigation
從中華菊頭蝠的種群數量分布來看,II類洞穴約占93.71%,并且大都集中分布于II類洞穴中的堂樂洞。堂樂洞的洞道結構復雜而寬闊,洞道分層且有多條分支,可供蝙蝠選擇的洞道、棲所及棲點極為多樣,洞內的水源及洞穴周邊的植被等環境條件也非常有利于蝙蝠的棲息,因此,只要人們不故意對其構成傷害,它們通常都會習慣性地在熟悉的棲點棲掛,并呈現某種程度上的棲點依戀性。堂樂洞雖屬旅游廢棄洞穴,但仍存在一定程度上的人為干擾,夏季高,冬季中等或偏低。
2.2 中華菊頭蝠的棲點特征
中華菊頭蝠主要棲息在洞道的D段(占91.60%),即離洞口150—200 m處(圖1),并且主要在離地高度6—10 m處棲息(占91.75%)(圖2)。其棲掛位點(棲點roostpoint)因季節而異(圖3),春、夏兩季喜棲于洞道的深處,秋季喜棲于洞口段,冬季多在洞道的中間段聚集。季節不同,其棲掛的高度也不同,春、夏、秋三季的棲高多在6 m以下,冬季則多在6—10 m的高處集群冬眠(圖4)。
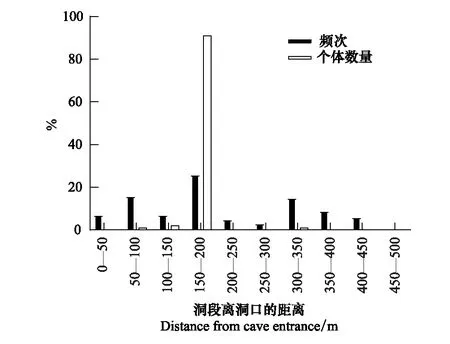
圖1 中華菊頭蝠(Rhinolophus sinicus)棲點距離洞口的個體數量(只次)及頻次分布Fig.1 The individual amount (individual-by-times) and frequency (times) distribution of Rhinolophus sinicus′ roostpoint distance from cave entrance頻次是指觀察到該蝠在每一洞段棲掛的次數(有時一次僅觀察到一只,有時可能是一群),個體數量是指該蝠種在每一洞段棲掛的只次(每次1只,即為1只次);個體數量在有些洞段占比過低,在圖上沒有顯示出來
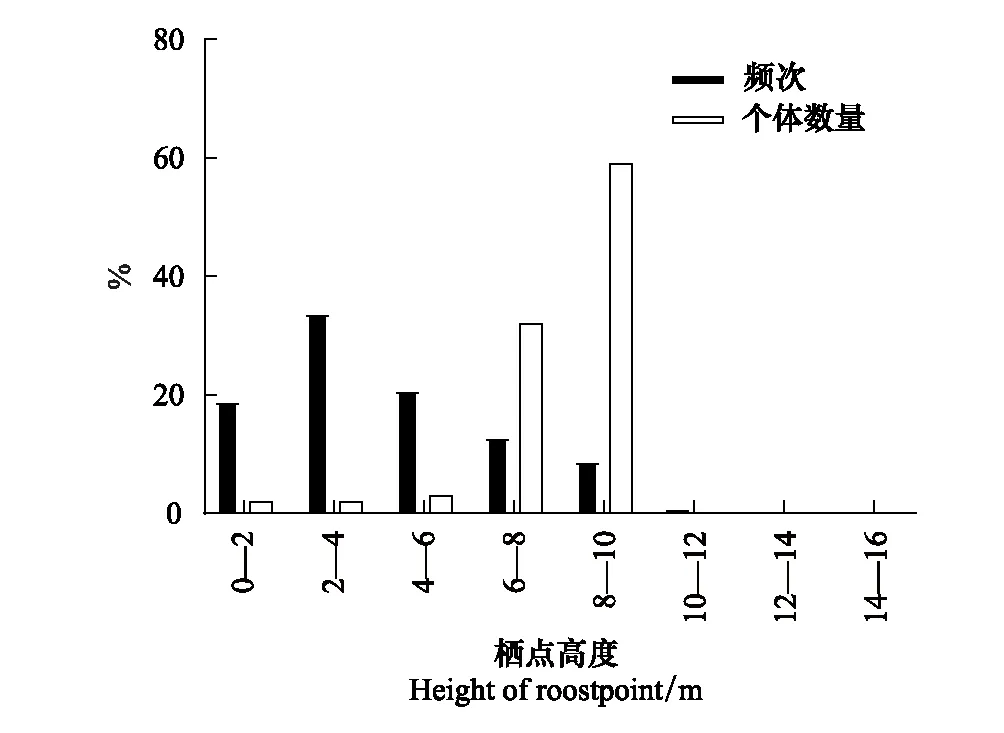
圖2 中華菊頭蝠(Rhinolophus sinicus)棲點距離地面高度的個體數量(只次)及頻次分布Fig.2 The individual amount and frequency (times) distribution of Rhinolophus sinicus′ roostpoint height from cave ground
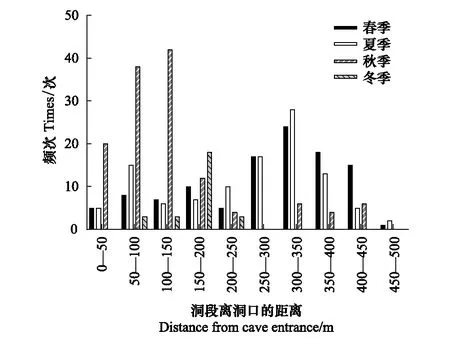
圖3 中華菊頭蝠(Rhinolophus sinicus)棲點離洞口距離的季變化Fig.3 Seasonal changes of Rhinolophus sinicus′ roostpoint distance from cave entrance
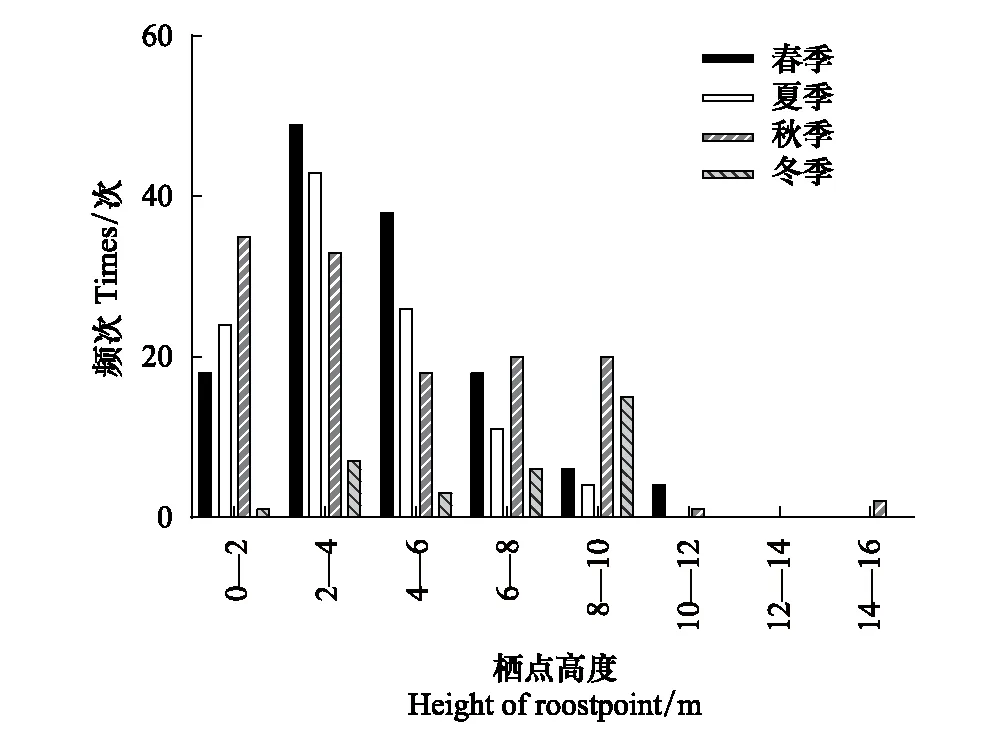
圖4 中華菊頭蝠(Rhinolophus sinicus)棲點距離地面高度的季變化Fig.4 Seasonal changes of Rhinolophus sinicus′ roostpoint height from cave ground
主要以“雙足倒掛”的方式棲于洞道的頂壁面(56.86%)、側壁面(25.44%)、凹窩內(15.49%)或狹縫間(2.21%)(圖5)。棲點安全性評價表明,安全性中等及以上的個體占69.32%,安全性低的個體占30.68%(圖6),總體上秋季的安全性偏低(圖7)。
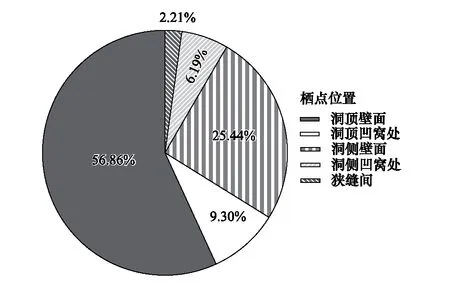
圖5 中華菊頭蝠(Rhinolophus sinicus)的棲點位置分布Fig.5 Distribution of Rhinolophus sinicus′ roostpoint location
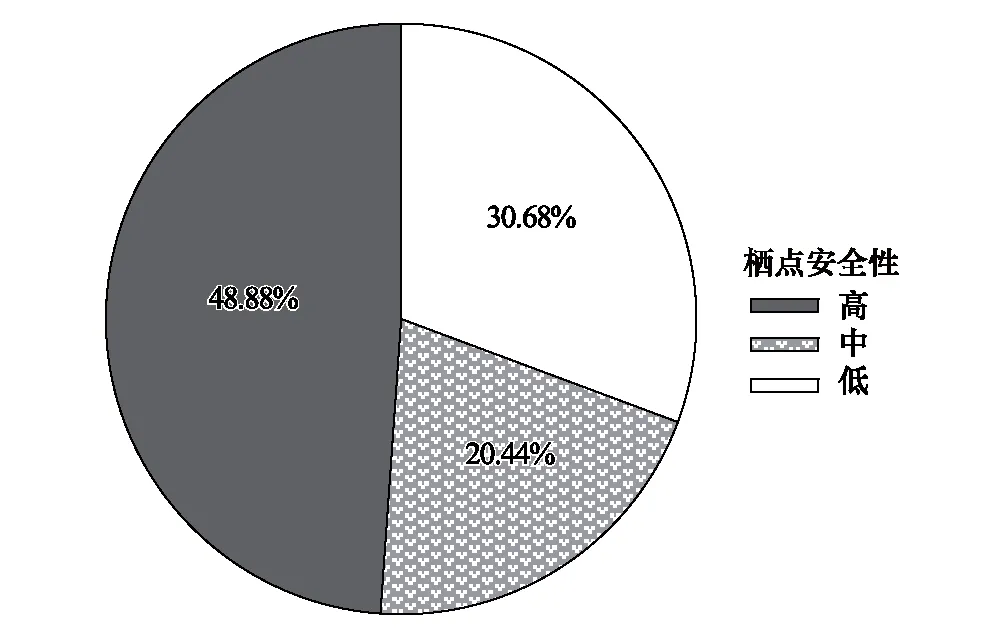
圖6 中華菊頭蝠(Rhinolophus sinicus)的棲點安全性分布Fig.6 Distribution of Rhinolophus sinicus′ roostpoint safety
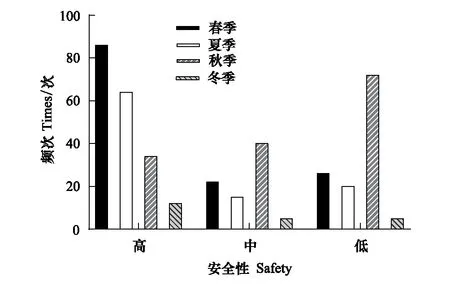
圖7 中華菊頭蝠(Rhinolophus sinicus)棲點安全性的季變化Fig.7 Seasonal changes in the roostpoint safety of Rhinolophus sinicus 安全性高、中、低的含義參見研究方法
2.3 中華菊頭蝠的體溫與棲點溫度
通過測量翼膜、背毛、耳和鼻四個部位的體表溫度,并進行統計分析,結果顯示:4個部位的體表溫度(體溫)之間不存在顯著性差異(P﹥0.05),并且變化也無規律性,介于10.4—27.8 ℃之間,平均值為(16.86±3.62)℃(n=80)。棲點溫度的變化范圍是10.3—24.3 ℃,平均值為(15.60±2.70)℃(n=80)。體溫和棲點溫度之間無顯著性差異(P>0.05),且兩者之間的變化呈現明顯的線性正相關關系,關系式為:Y=0.9860X+1.554(Y為體溫,X為棲點溫度,X∈[10.3, 24.3],R2= 0.8886,圖8)。
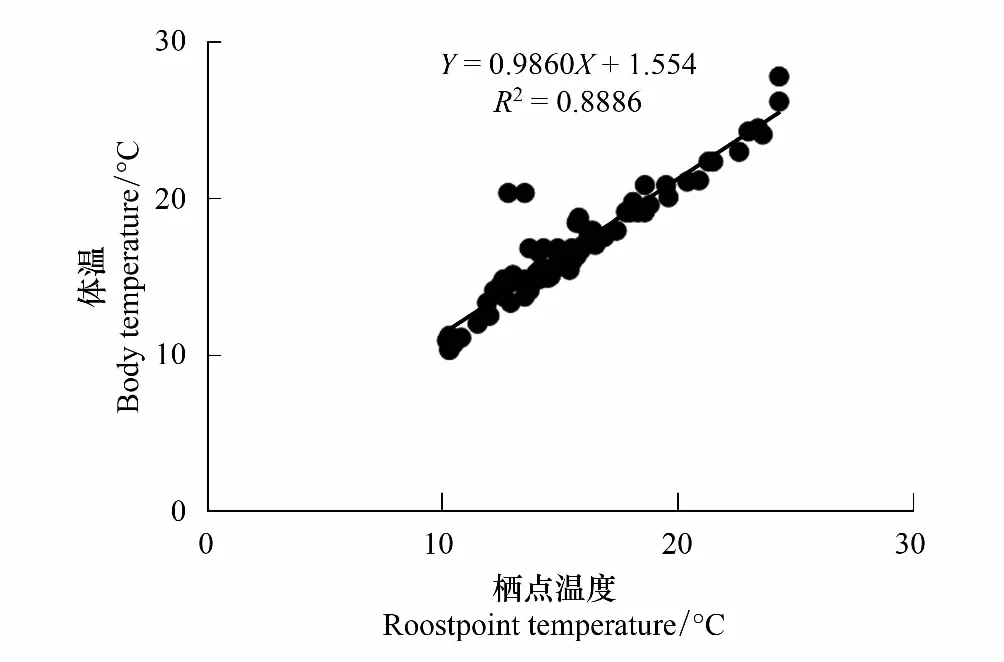
圖8 中華菊頭蝠(Rhinolophus sinicus)的體溫與棲點溫度之間的線性關系Fig.8 Linear relationship between body and roostpoint temperature of Rhinolophus sinicus
3 討論
3.1 中華菊頭蝠的棲息生態特征
3.1.1中華菊頭蝠的棲點選擇、棲掛姿勢、休眠與集群習性
在湘西州境內的溶洞中,中華菊頭蝠的數量相對較多,較為常見[4],全年都可見其棲息于洞道的頂壁或側壁,通常在洞穴的較深處呈“雙足倒掛”的棲姿,即以雙足爪抓扣住棲點而使身體懸空垂直向下倒掛的姿勢棲息。這明顯不同于西南鼠耳蝠的“腹部貼壁式”掛姿或臥姿[34],而皮氏菊頭蝠“單足倒掛”的棲姿較為常見[35]。
雖然中華菊頭蝠所棲息的溶洞的海拔高度介于248—1083 m之間,但主要棲息在海拔為330—380 m的溶洞中,喜棲于離洞口150—200 m的洞段和離地高度為6—10 m的洞頂壁及側壁。在同一冬眠洞穴中,西南鼠耳蝠多散棲于洞口段,皮氏菊頭蝠常集小群或獨棲于洞道的深處,而中華菊頭蝠的獨棲及集群冬眠處多介于兩者之間。
日眠期間的個體淺睡易醒,人通常不易接近。育幼場所非常隱蔽,人跡罕至,安全性高,有的洞道人體不能進入。雖然獨棲個體并不少見,但大多集群活動,通常能見到數只至數十只緊靠(圖9)或非緊靠(圖10)在一起棲息的情況。冬眠期間,則可聚集成數百只的大群。集群是一個動態的過程,一般在每年的9月中下旬開始集群,數量逐漸增多,原來分散在不同洞道中棲息的個體都聚集起來,但更多的個體可能是從其他的洞穴中飛來集群的,在堂樂洞數量最多時可聚集達到500只以上(圖11)。至次年3月底,這些聚集的個體陸續從冬眠中醒來,之后逐漸散開,飛往其他洞穴或散棲于該洞穴的不同洞道。

圖9 中華菊頭蝠個體的緊靠式棲姿 Fig.9 Snuggling roosting of Rhinolophus sinicus individuals

圖10 中華菊頭蝠小群的非緊靠式棲姿 Fig.10 Non-snuggling roosting of Rhinolophus sinicus grouplet

圖11 中華菊頭蝠的大型冬眠聚群Fig.11 Large hibernating group of Rhinolophus sinicus
冬眠期間,中華菊頭蝠的個體數量雖相對穩定,但也是動態的。一方面,蝙蝠可能受到人為或其他因素的干擾,醒來后會飛走。另一方面,即使沒有人為干擾或其他因素的影響,蝙蝠也會自然醒來。因為冬眠型動物在整個冬眠季節通常會出現幾個冬眠期,其間會有周期性的短暫覺醒[36]。而處于冬眠狀態的蝙蝠對周圍環境的變化仍保持一定的警覺性。在調查過程中,通過紅外相機拍攝發現,大多數中華菊頭蝠在受到外界刺激醒來后改變了位置,只有少數個體回到了原來的棲點。如果洞道較短,可供選擇和利用的棲點較少,蝙蝠在冬眠過程中長期受到強烈干擾后,也將被迫飛出洞外,尋找其他洞穴冬眠,從而增加能量消耗,影響越冬存活率。
野外觀察還發現,中華菊頭蝠數只、數十只乃至數百只個體緊靠或鄰近在一起冬眠,但并非所有的個體都“睡得很死”,有的個體睡得很淺,有的個體則處于清醒狀態,一旦發現周圍異常,有燈光照射或有人靠近,就會很快飛走,并在周圍穿梭或盤旋,在展翅飛離和盤旋的過程中會將周圍“淺睡”的個體驚醒,隨后處于不同深度冬眠狀態的個體會被逐個“喚醒”而飛走,但也有少數個體一直處于“沉睡”狀態而“喚不醒”。于是,就有“著急的”的“早醒者”直接飛過去,撞擊它們,試圖將它們撞醒,稱這種現象被稱為“撞醒行為”(bumping for awakening companions)[14]。經過多次撞擊后,“沉睡者”也會很快地蘇醒過來,并迅速飛走。推測“清醒者”或“淺睡者”可能就是“哨兵”,而“哨兵”是輪換值班的,即不同的個體進入深睡狀態的時間可能不同,并且同一個體在整個冬眠期間可能存在多個沉睡期和覺醒期,而不同個體的沉睡期與覺醒期可能相互交錯[14],從而使群體安全得到保障。
3.1.2中華菊頭蝠的體溫
中華菊頭蝠的體溫介于10.4—27.8 ℃之間、棲點溫度范圍為10.3—24.3 ℃,兩者皆明顯高于大菊頭蝠[33]和西南鼠耳蝠[34],而與皮氏菊頭蝠[35]相近(表2)。其主要原因可能是,大菊頭蝠和西南鼠耳蝠通常在洞口段棲息,而皮氏菊頭蝠和中華菊頭蝠通常在洞穴的較深處棲息。洞穴深處的空氣溫度與濕度相對穩定,日變化與季節性變化的幅度很小,基本上是在當地多年平均氣溫的附近波動[14]。可是,就體溫及棲點溫度變化范圍而言,中華菊頭蝠和皮氏菊頭蝠的幅度較大,而大菊頭蝠和西南鼠耳蝠較小,這可能是后兩者主要是在湘西州域內越冬,所測數據幾乎都是冬眠期數據的原因所致。
蝙蝠等小型哺乳類動物在冬眠期主要依靠體內儲存的脂肪維持最低的能耗[36—39]。近年的調查數據[33—35,40]顯示(表2),洞棲性蝙蝠的體溫是變化的,在洞道中日眠和冬眠期間,其體溫會降至比棲點溫度略高的狀態,這可能有利于節省能量。中華菊頭蝠常集大群冬眠,且個體之間通常緊靠在一起,在自然深睡的狀態下翼膜緊緊地包裹著身體,顯然這也有助于減少能耗。更值得注意的是,集大群冬眠不僅有利于體溫調節,還可如上述那樣,以“輪留值班放哨”的方式有利于群體自身的保護。
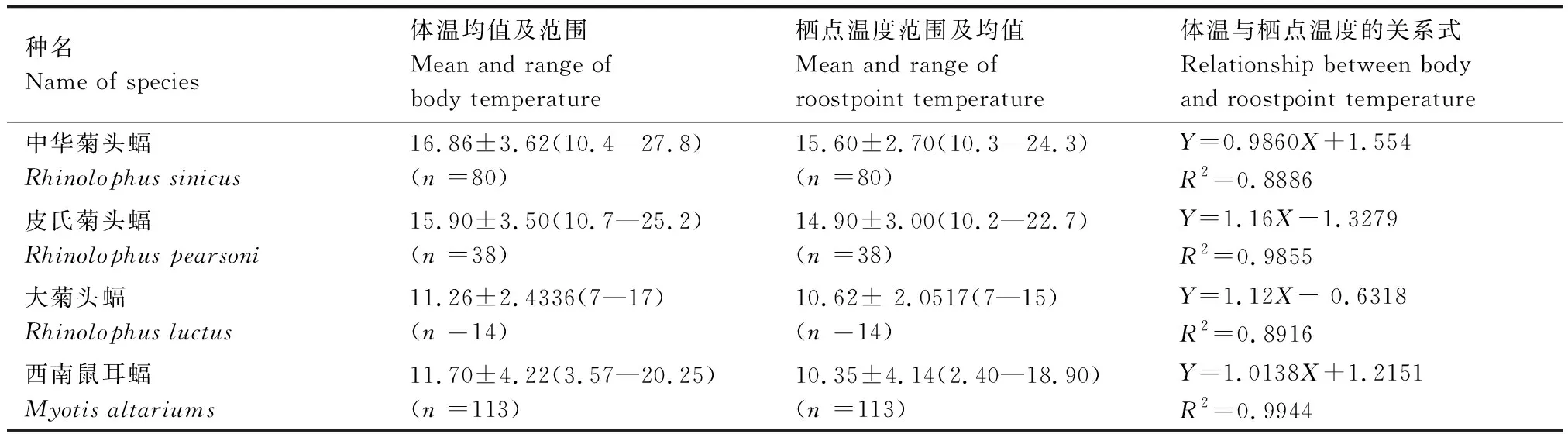
表2 四種洞棲性蝙蝠的體溫與棲點溫度/℃Table 2 Body and roostpoint temperature of four cave-dwelling bat species
3.2 種群保護
翼手目是哺乳動物綱中的第二大類,棲息方式多種多樣,主要有樹棲性、宅棲性、洞棲性和兼棲性4種類型[14,41]。中華菊頭蝠是典型的洞棲性蝙蝠,白天隱藏在洞內“日眠”,傍晚才開始出飛,到洞外覓食,主要捕食夜間活動的昆蟲等無脊椎動物,飽食后飛回洞內,食物經消化后,大量的糞便被排到棲掛處下方的地面上,日積月累,可堆積成為非常厚實寬大的糞場,為洞內許多動物的生存提供營養與能量來源[12—14]。因此,不僅對于洞外自然生態系統及農林生態系統的穩定發揮著重要功能,而且對于許多洞穴動物類群的生存和繁衍也起著關鍵性的作用。
中華菊頭蝠等洞棲性蝙蝠的活體、尸體和糞便等排泄物都可成為許多洞穴生物生活的營養條件及藏身之處。小泡巨鼠(Leopoldamysedwardsi)是湘西州境內溶洞及廢棄礦洞中較為常見的大型鼠類,其攀爬及跳躍能力很強,有時可竄到蝙蝠棲掛之處捕食蝙蝠,對蝙蝠種群存在一定的威脅;灰林鸮(Strixaluco)等鸮形目鳥類傍晚時分經常在洞口附近守候出飛的蝠群,伺機捕食,有時還可見其飛進洞內搜捕蝙蝠[14]。蝙蝠的活體還是洞內蜱、螨、蠕蟲、線蟲、原蟲等內、外寄生生物,以及病毒、病菌等微生物的主要宿主[12—14,36,42—45]。蝙蝠的尸體及糞便更是洞穴甲蟲、蠅、蚊、潮蟲、馬陸、軟體動物、蚯蚓、扁蟲、原蟲等無脊椎動物及菌類等微生物的主要營養、能量來源,或依托、藏匿場所。因此,洞棲性蝙蝠在維持洞穴生物多樣性與洞穴生態系統穩定方面起著不可或缺的作用,它們猶如一把巨“傘”傘護著許多洞穴生物的生存和繁衍,因此加強對洞穴生態系統中蝙蝠傘護性的研究有助于洞穴生態保護教育及對蝙蝠種群的保護實踐[14]。
鑒于蝙蝠廣泛的生態作用與巨大的經濟價值[46—50],倡議將“五月五日”設立為“國際蝙蝠日”,因為這段時間蝙蝠遷徙頻繁,開始繁育,也有“五五有福”或“五福臨門”的喻意,如此既能夠廣泛地宣揚中國傳統的“蝠文化”,又有助于推進全球性的生態文化建設與蝙蝠資源保護。
致謝:張佑祥、黃興龍、荀二娜老師,彭樂、張佩玲、蔣能等同學參與了部分野外調查和數據處理工作,特此致謝。
