灤河干流中上游浮游細菌群落多樣性及其影響因素
2022-08-03 01:55:10田雨露董滿宇關夢茜
生態學報 2022年12期
劉 洋,劉 琦,田雨露,董滿宇,徐 霞,關夢茜,江 源,*
1 中藥資源保護與利用北京市重點實驗室, 北京 100875 2 北京師范大學地理科學學部, 北京 100875
城市工業和生活廢水的排放,使得大量含氮、磷物質進入河流,導致水體富營養化,造成河流的嚴重污染和水生生態系統的退化[1]。浮游細菌作為水體環境中元素循環和能量流動的主要承擔者,在生物地球化學循環過程中起著關鍵作用[2—5]。河流中的浮游細菌與水環境具有較為復雜的相互作用,其群落多樣性的變化能敏感地反映出水體環境的變化[6—7]。有研究表明,人類活動產生的外源性營養輸入會對水體浮游微生物群落產生顯著影響[8]。由于居民的生活用水等通常來自于周邊河流,水體中致病微生物的種類和來源也與公共衛生問題密切相關[9]。研究水體浮游細菌群落特征隨水環境的變化將有助于深入揭示人類活動對水生態的影響特征。
然而,以往有關的研究多集中于季節、水體理化指標、營養鹽指標等對浮游細菌群落的影響方面,通常并未考慮河岸帶土地利用因素的影響作用。河岸帶的土地利用結構和組成,對水體環境中的營養鹽濃度以及水文過程等均會產生深刻影響,進而影響到河流浮游微生物的群落結構[10—11]。河岸帶范圍內較高精度的土地利用空間數據可以借助遙感技術獲得,通過將之整合進水環境因素對浮游細菌群落影響的分析之中,有助于辨明河岸帶土地利用組成結構與水環境以及浮游細菌群落間的關系,進而為解決水污染及其導致的環境問題,例如水華等提供新的研究視角和研究方法。
本文中的案例研究區位于灤河中、上游一帶,其匯入的潘家口水庫和大黑汀水庫是“引灤入津”工程的重要水源地。水體浮游細菌群落與水質的關系,將直接反映“引灤入津”工程中的水資源質量,也關聯著天津市民的飲水安全。通過研究河岸帶土地利用和水環境因子以及與河流浮游微生物群落多樣性間的關系,明確水環境對浮游微生物群落的影響,揭示河岸帶土地利用變化的水生態效應,將有助于加深對河流生態系統的了解,并為灤河流域的土地利用結構優化和河流生態健康保護提供案例參考和科學支撐。
1 材料與方法
1.1 研究區域概況
研究區位于華北平原東北部的灤河流域中上游區域(115°32′—118°53′E, 40°11′—42°45′N),總面積3.6×104km2。灤河發源于內蒙古和河北的壩上高原,干流迂回曲折,全長約620 km,水系發達,支流眾多,自西北向東南流經內蒙古自治區和河北省的多個縣、旗、區,其中上游諸支流匯入灤河干流后,于河北省遷西縣流入潘家口水庫和大黑汀水庫,年平均徑流量13.3×108m3[12—13]。研究區地處典型的溫帶大陸性季風氣候區,多年平均氣溫6.1℃;月平均氣溫7月最高(21.9℃),1月最低(-12.0℃)。年平均降水量444.1mm(http://data.cma.cn)。研究區內林地、農業用地、建設用地、草地、水域和其他用地占比分別為56.9%、18.3%、3.9%、17.9%、1.6%和1.5%,其中草地主要分布在西北部的上游地區,農業用地主要分布在河流兩岸的沿河平坦階地一帶,建設用地主要分布在南部各個支流與干流的匯流處(圖1)。
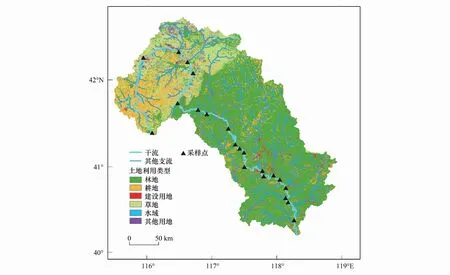
圖1 灤河流域及采樣點分布示意圖Fig.1 Distribution of sampling sites in the Luanhe River basin
1.2 樣品采集
依據分層采樣原則,樣點覆蓋灤河干流中上游河岸帶各類土地利用類型,同時也盡可能考慮空間分布的均勻性、交通可達性等因素,共布設樣點21個,于2018 年4月—5月(春季)進行水樣采集,流程如下:
(1)用于測定水質指標的水樣:使用1L有機玻璃采水器在采樣點所在位置河心或距離河岸5 m 處進行水樣采集(深度約為0.5m),采集3次并在容器中充分混合。將混合后的水樣分為2份,一份為不做任何處理的原始水樣;另一份水樣經孔徑為0.45 μm的Whatman GF/C玻璃纖維膜(Whatman,UK)過濾,過濾后的濾膜避光冷藏保存,用于測定葉綠素a(Chla)和總懸浮固體(SS)。兩份水樣均加入少量濃硫酸(H2SO4)酸化保存。盛裝水樣的聚乙烯樣品瓶預先用1∶10鹽酸溶液浸泡24h,用蒸餾水洗凈并晾干;玻璃纖維素濾膜預先在450℃溫度下烘烤6h,并用萬分之一天平稱取其質量,以供后續懸浮性固體的測定。水樣保存在低溫保存箱中,并在短時間內運回實驗室,在-20℃的條件下冷凍儲存。
(2)用于16S rDNA測序的水樣:使用預先消毒過的聚乙烯桶收集,采集原則同上,每個樣點收集三份平行樣本(每個0.9L)。水樣先后通過2種孔徑的濾膜(2.0μm, 0.8μm,Whatman,UK)進行過濾,以去除砂礫和雜質,增加后續富集效率,并使得絕大多數浮游微生物通過[14—16]。最后使用孔徑為0.22μm的混合纖維素濾膜(Whatman,UK)對水樣進行過濾,將浮游細菌富集在濾膜上。濾膜用鋁箔包裹后保存在液氮罐中(-196℃)。
1.3 水質指標測定
水質指標測定分為現場測定和實驗分析兩部分,共測定12個指標。(1)現場測定:溶解氧(DO)使用便攜式溶解氧儀(YSI Incorporated Company,USA)測定;流速(Vel)使用流速儀(FLOWATCH,JDC Electronic SA, Switzerland)測定;電導率(EC)、pH和水溫(Tem)使用電導率儀(METTLER-SG DUO,Mettler Toledo international LTD,USA)測定[17]。(2)實驗分析:高錳酸鹽指數(CODMn)、總氮(TN)和總磷(TP)使用原始水樣測定;CODMn采用高錳酸鹽滴定法;TN 采用紫外分光消解法;TP 采用鉬銻抗分光光度法;氨氮(NH3-N)和硝酸鹽氮(NO3-N)使用過濾水樣測定;NH3-N 采用納氏試劑比色法;NO3-N 采用紫外分光光度法;懸浮性固體(SS)和葉綠素a(Chla)使用濾膜上的殘余物進行測定;SS 采用稱重法測定;Chla 采用丙酮分光光度法進行測定[18]。
1.4 細菌16S rDNA的提取、處理和測序
將附有浮游細菌的0.22μm混合纖維素薄膜溶解于2 mL丙酮(Beijing Chemical Works,A.R.)中,使用E.Z.N.A. Water DNA Kit 試劑盒(Omega,USA)提取DNA。使用簡并PCR引物799F[19]和1193R[20]對細菌的16S rDNA基因可變區V5—V7進行PCR擴增,每份三組平行。反應條件如下:98℃預變性30 s;98℃變性10 s,55℃復性15s,72℃延伸60 s,循環25次;72℃延伸5 min。反應產物作為第二輪PCR的擴增模板,除引物換為含有Illumina接頭和連接區序列的引物外,其他反應條件不變。對第二輪PCR的產物進行純化并混合,得到最終的測序文庫。文庫送交華大基因,依托HiSeq 2500測序平臺(Illumina Inc.,USA)進行高通量測序。
使用QIIME1.9.1、USEARCH 10.0處理16S rDNA序列,丟棄錯誤率超1%的序列,最終得到3414503條高質量序列(平均每個樣本65664條,最小值為27829條,最大值為100572條)。以97%的閾值對序列進行聚類,得到18360個操作分類單元(Operational Taxomic Unit,OTU)。使用PyNAST方法去除非細菌的16S rDNA序列后,得到15856個OTU,使用Usearch 10生成OTU表。OTU表中的每一個樣本都被重抽樣至30000條序列的樣本大小,以每個OTU的豐度除以30000的樣本大小,計算OTU的相對豐度。使用RDP數據庫對OTU代表序列進行注釋,從而得到含有物種信息的OTU表。
1.5 空間數據提取
土地利用數據來源于中國科學院計算機網絡信息中心國際科學數據服務平臺,數據基于2016年30m分辨率的Landsat TM/ETM 影像(http://www.cnic.cas.cn/)。土地利用解譯為6大類:(1)林地,包括喬木林和灌木林;(2)農業用地,包括菜地、果園和其他農用地;(3)建設用地,包括居民、交通、商業和工業用地;(4)草地;(5)水域,包括河流、濕地和水庫;(6)其他用地。根據美國地質調查局的30m分辨率數字高程模型(DEM)獲取河道平均坡度和地貌形態數據(http://glovis.usgs.gov/),基于國家基礎地理信息中心的1∶250000 矢量水系圖獲取水系數據(http://ngcc.sbsm.gov.cn/),使用軟件ArcGIS 10.1中的水文分析(Spatial Analyst Tools-Hydrology)模塊提取每個樣點的上游集水區。對于每個樣點的集水區,先根據河流水系圖的雙側250m圓角矩形緩沖區進行剪切,得到距河流垂直距離在250m以內的河岸帶緩沖區;再使用以樣點為圓心的3000m圓形緩沖區對上一步的河岸帶緩沖區進行剪切,獲得從樣點起至上游3000m范圍內距河流垂直距離在250m以內的樣點河岸帶緩沖區(圖2)。統計各樣點的緩沖區內草地、林地、農業用地、建設用地的面積占該四種土地利用類型面積總和的比例,用于后續分析。
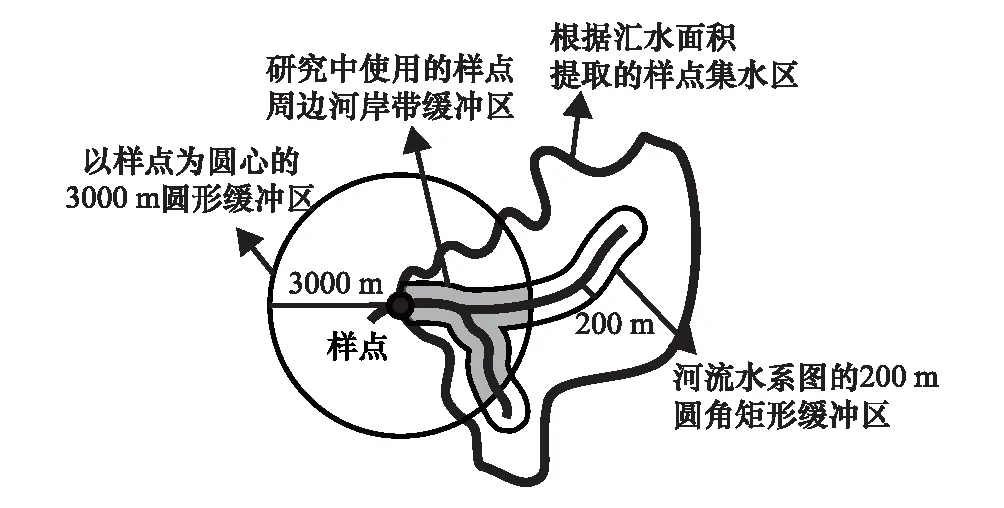
圖2 河岸帶緩沖區的截取方式Fig.2 Method of extraction of riparian buffer
1.6 數據處理與分析
使用Shapiro-Wilks檢驗水環境因子是否符合正態分布,對不符合的數據進行ln(x+1)變換。將每個樣點平行樣本的OTUs豐度取均值,選取其中出現頻率 ≥ 90%的OTU作為核心群落[21]。根據淡水細菌數據庫對OTUs進行分類學注釋,將不能與數據庫匹配的,即未在傳統的靜水水體(例如湖泊)細菌學研究中出現過的OTU記為“非典型淡水分類群細菌”[22]。多樣性指數的計算在R語言環境中使用軟件包“Vegan”完成。選取Chao1豐富度指數(Chao1 richness estimator,Chao1)和Shannon′s多樣性指數(Shannon′s Diversity Index,SHDI)作為群落α-多樣性的表征。選擇計算各樣本OTU水平上的Bray-Curtis距離(Bray-Curtis Dissimilarity)矩陣,作為對β-多樣性的表征。使用Spearman 相關分析計算核心群落占比、非典型淡水細菌類群、多樣性指數和距源頭距離之間的相關性,和α-多樣性指數與水環境因子、土地利用因子之間的相關性。使用Mantel檢驗和偏Mantel檢驗,計算細菌群落的Bray-Curtis距離矩陣和地理空間因子(距源頭距離)、水質、土地利用因子的歐式距離矩陣之間的相關性。
2 結果與分析
2.1 灤河各水質指標特征
灤河各水質指標顯示(表1):河流理化指標中,pH平均為8.54,呈弱堿性;水溫(Tem)平均為13.82 ℃;流速(Vel)平均為1.35 km/h;電導率(EC)平均為417.78 μS/cm;懸浮性固體(SS)平均為30.36 mg/L;溶解氧(DO)平均為8.91 mg/L,達到Ⅰ類水質;高錳酸鹽指數(CODMn,)平均為5.22 mg/L,達到Ⅲ類水質。河流營養鹽指標中,總磷(TP)平均為0.11 mg/L,達到Ⅲ類水質;總氮(TN)平均為2.63 mg/L,為劣Ⅴ類水質;氨氮(NH3-N)平均為0.30 mg/L,達到Ⅱ類水質;硝酸鹽氮(NO3-N)平均為1.82 mg/L。生物指標葉綠素a(Chla)平均12.71 μg/L。
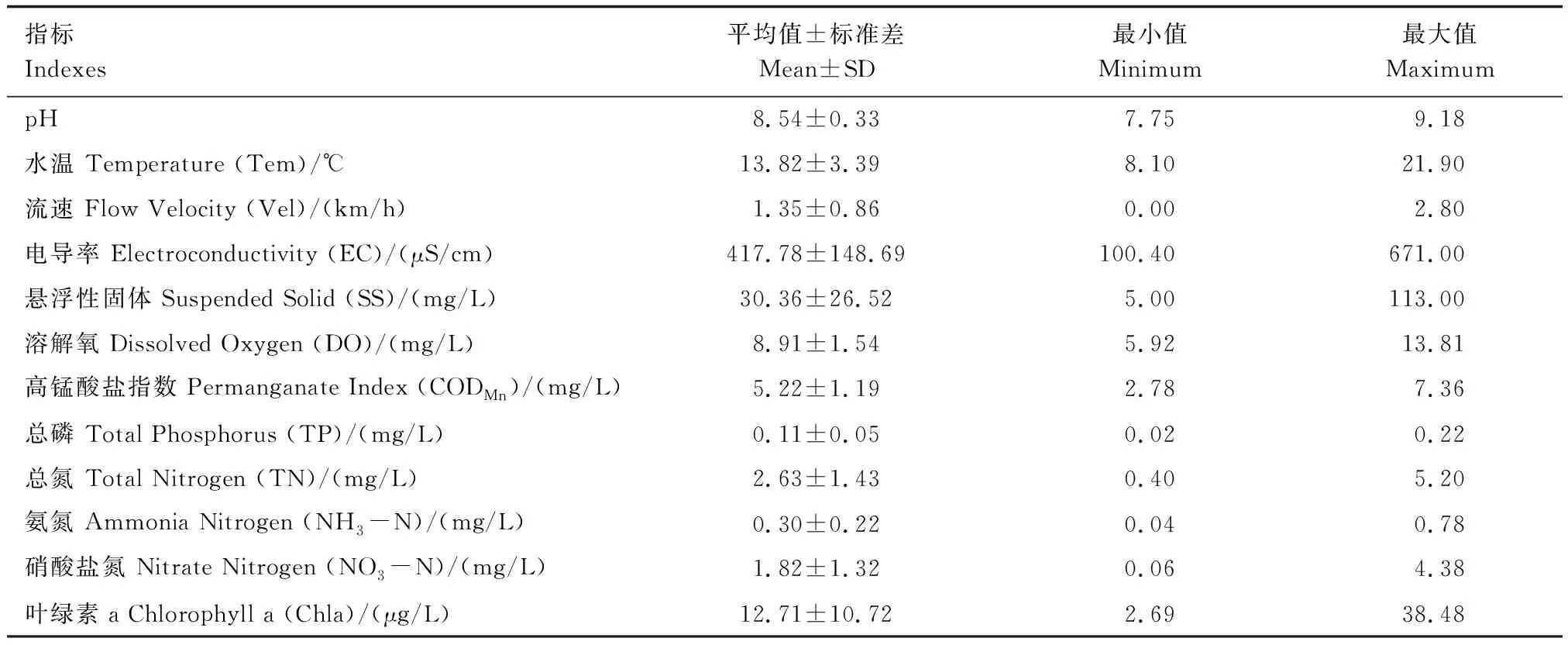
表1 灤河各水質指標特征Table 1 Characteristics of water quality parameters in Luanhe River
2.2 灤河中上游浮游細菌群落的組成特征
使用RDP數據庫對每個OTU的代表序列進行注釋后,對其進行從門到屬水平的分類。在檢出的15856個OTU中,共檢測到分類地位明確的細菌界28門(Phylum)、72綱(Class)、113目(Order)、265科(Family)和936屬(Genus)。其中變形菌門(Proteobacteria)由于種類眾多,根據核糖體rRNA序列的差異,將其拆分為5個綱,分別冠以α、β、γ、δ和ε前綴,以便于研究。
研究共確定出12個的優勢細菌門(綱),它們在至少一個樣本中相對豐度大于1%。依據相對豐度的大小排序,分別為:放線菌門(Actinobacteria,平均28.95%)、β-變形菌綱(β-Proteobacteria,平均28.90%)、α-變形菌綱(α-Proteobacteria,平均10.62%)、擬桿菌門(Bacteroidetes,平均6.14%)、γ-變形菌綱(γ-Proteobacteria,平均5.57%)、δ-變形菌綱(δ-Proteobacteria,平均4.51%)、厚壁菌門(Firmicutes,平均3.93%)、酸桿菌門(Acidobacteria,平均3.87%)、綠彎菌門(Chloroflexi,平均2.24%)、疣微菌門(Verrucomicrobia,平均1.06%)、浮霉菌門(Planctomycetes,平均1.01%)、硝化螺桿菌門(Nitrospirae,平均0.75%),其余20種細菌門(綱)組成了低豐度類群,平均為1.46%。變形菌門和放線菌門的平均相對豐度高于50%,是灤河干流中占據優勢地位的門類。
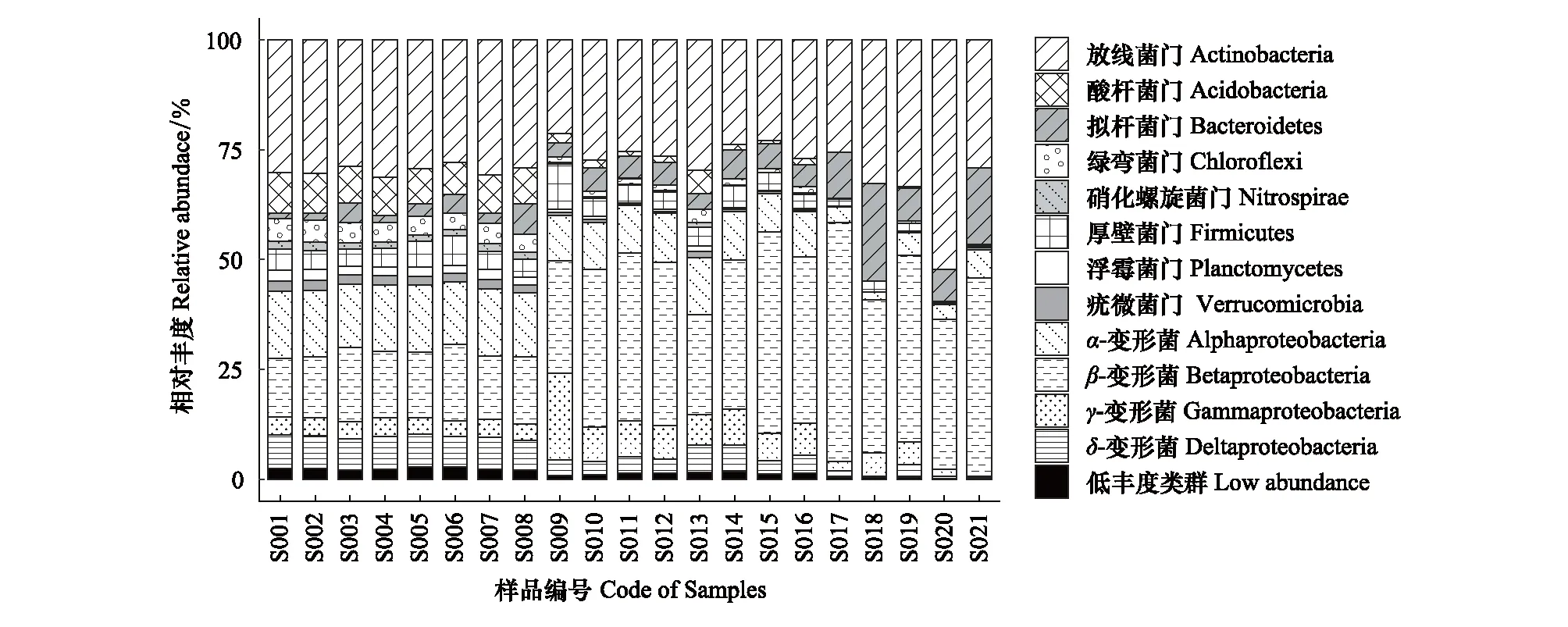
圖3 灤河干流中上游浮游細菌群落組成Fig.3 Composition of bacterioplankton community in upper-middle reach of Luanhe river
灤河中上游浮游細菌的核心群落共有209個OTU,占OTU總數(15 856個)的1.29%,序列數占總序列數的43.04%。核心群落的相對豐度占比與距源頭距離存在顯著正相關,Spearman相關系數為 0.909(P<0.01)。非典型淡水分類群細菌的相對豐度與距源頭距離存在顯著負相關,Spearman相關系數為-0.935(P<0.01)。
2.3 灤河中上游浮游細菌群落的α-多樣性特征
Chao1豐富度指數和Shannon′s多樣性指數計算描述的統計特征如下(表2)。
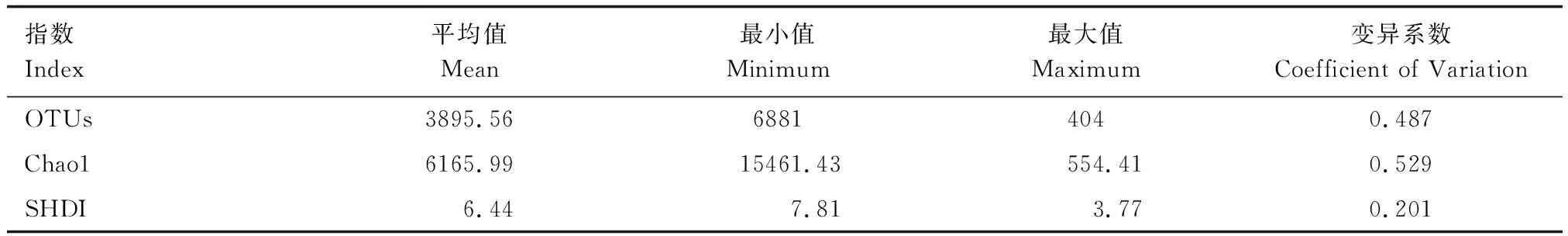
表2 浮游細菌群落α-多樣性指數的描述統計Table 2 Descriptive statistics of α-diversity Index of bacterioplankton community
OTU數和Shannon′s多樣性指數均成階梯狀下降趨勢,而Chao1豐富度指數則在河流中段首先增大,隨后迅速降低(圖4)。
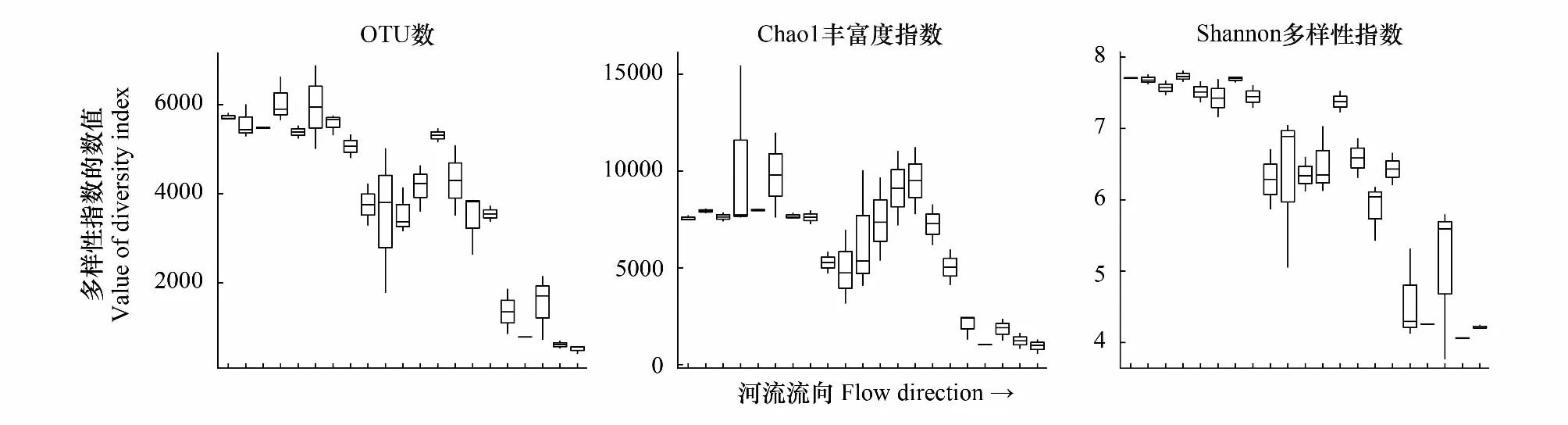
圖4 α-多樣性指數隨流向的變化Fig.4 Variation of α-diversity indexes along the flow direction
浮游細菌群落的α-多樣性指數與各項環境因子的Spearman相關性分析如表3所示。
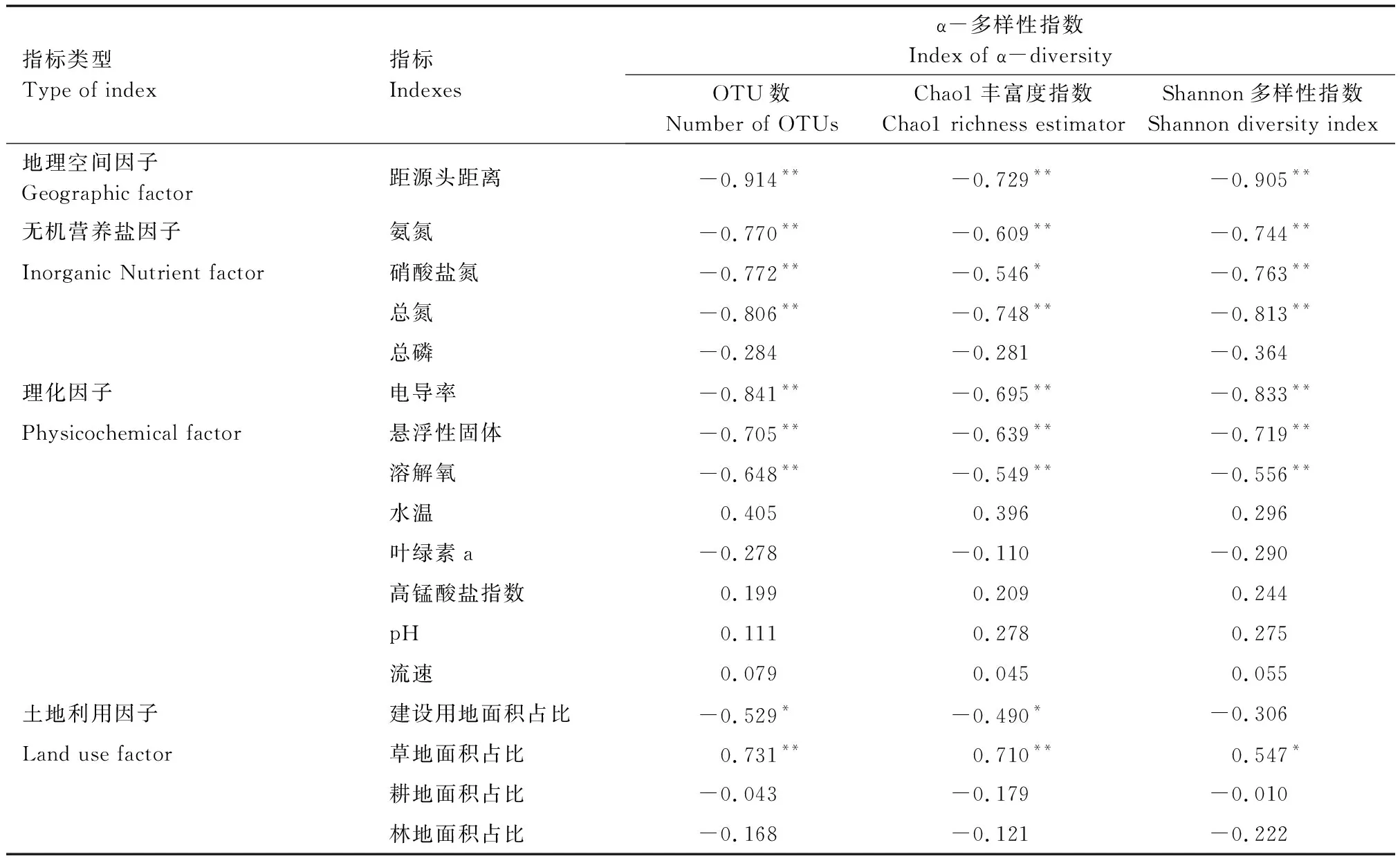
表3 浮游細菌群落α-多樣性指數與環境因子和土地利用因子的相關性分析Table 3 Correlation analysis between α-diversity index of bacterioplankton community and environment and land use factors
距源頭距離、建設用地占比(URBAN)、氨氮(NH3-N)、硝酸鹽氮(NO3-N)、總氮(TN)、電導率(EC)、總懸浮固體(SS)和溶解氧(DO)與OTU數、Chao1豐富度指數和Shannon′s多樣性指數間均存在顯著負相關(P<0.01)。
草地占比(GRASS)與OTU數、Shannon′s多樣性指數(P<0.05)和Chao1豐富度指數均存在顯著正相關(P<0.01);而總磷(TP)則與三項指標均不存在相關性。
2.4 灤河中上游浮游細菌群落的β-多樣性特征及其與水環境因子和土地利用因子的關系
基于Bray-Curtis距離的主坐標分析(PCoA)揭示了灤河中上游各樣點所取平行樣本中細菌群落的分布模式(圖5)。細菌群落PCoA排序圖的第一軸(PC1)和第二軸(PC2)分別能解釋55.27%和16.27%的物種差異,其中PC1軸與距源頭距離存在顯著負相關關系(Spearman相關系數-0.891,P<0.05)。樣本分為三部分:位于排序圖右側的樣本,距源頭距離較近,核心群落占比較低,而非典型淡水分類群細菌占比較高;位于排序圖左下側的樣本,距源頭距離較遠,核心群落占比較高,而非典型淡水分類群細菌占比較低。位于排序圖上側的樣本,位于河段的中部,其較為分散的分布模式暗示中游樣本的群落組成存在一定的差異,體現出過渡性的特征。

圖5 河流沿途樣點各平行樣本的浮游細菌群落組成的PCoA 分析Fig.5 PCoA analysis of bacterioplankton community of parallel samples along the river各樣點的平行樣本按距源頭距離由近及遠編號,距源頭距離最近的樣本編號為S001,以此類推
使用Mantel檢驗和偏Mantel檢驗,對各項水環境因子的歐式距離矩陣和細菌群落組成的Bray-Curtis距離矩陣進行相關性檢驗,結果顯示:距源頭距離與細菌群落組成存在顯著相關,相關系數為0.723(P<0.01),在將其他水質因子作為控制變量后,兩者依然顯著相關(P<0.01)(表4)。電導率、總氮、硝酸鹽氮、氨氮、總懸浮固體、溶解氧、葉綠素a、pH、流速均與細菌群落組成存在顯著相關(P<0.05);在將距源頭距離作為控制變量后,只有總氮、硝酸鹽氮、氨氮仍與細菌群落組成存在顯著相關(P<0.05),這說明這三種氮營養鹽指標其含量不完全隨滯水時間的增加而改變,存在空間異質性。
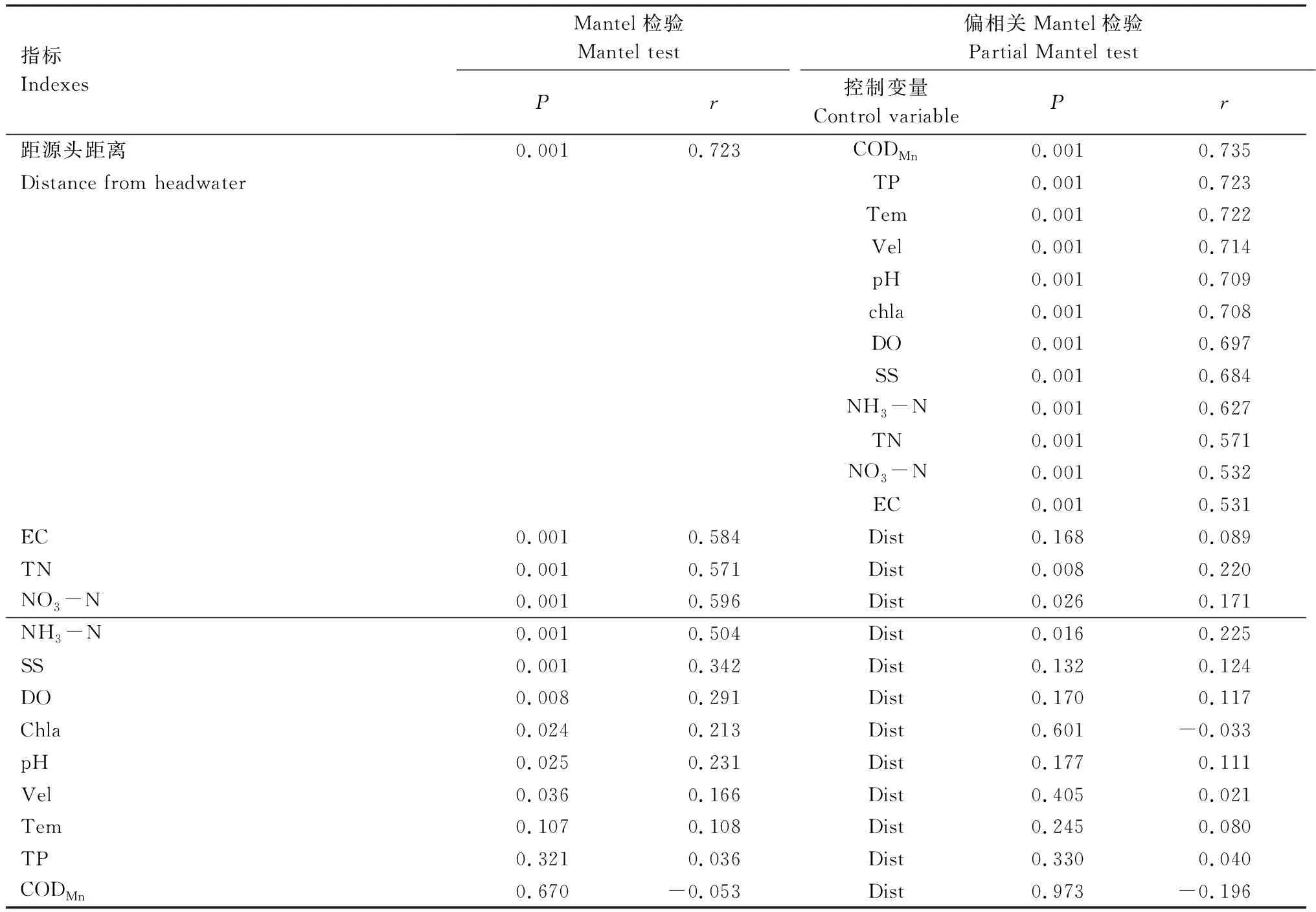
表4 群落Bray-Curtis距離矩陣與水質因子歐式距離矩陣的Mantel檢驗Table 4 Mantel test between community composition and water environment factors excluding land use factors
將土地利用占比作為控制變量后,距源頭距離與細菌群落組成仍存在顯著相關(P<0.01)(表5)。土地利用因子中,只有建設用地占比(URBAN)與細菌群落組成存在顯著相關(P<0.05);將距源頭距離作為控制變量后,建設用地占比與細菌群落組成在α=0.05的顯著性水平上不再相關,但在α=0.1的顯著性水平上依然相關(P<0.1),說明建設用地對細菌群落的影響存在空間異質性。
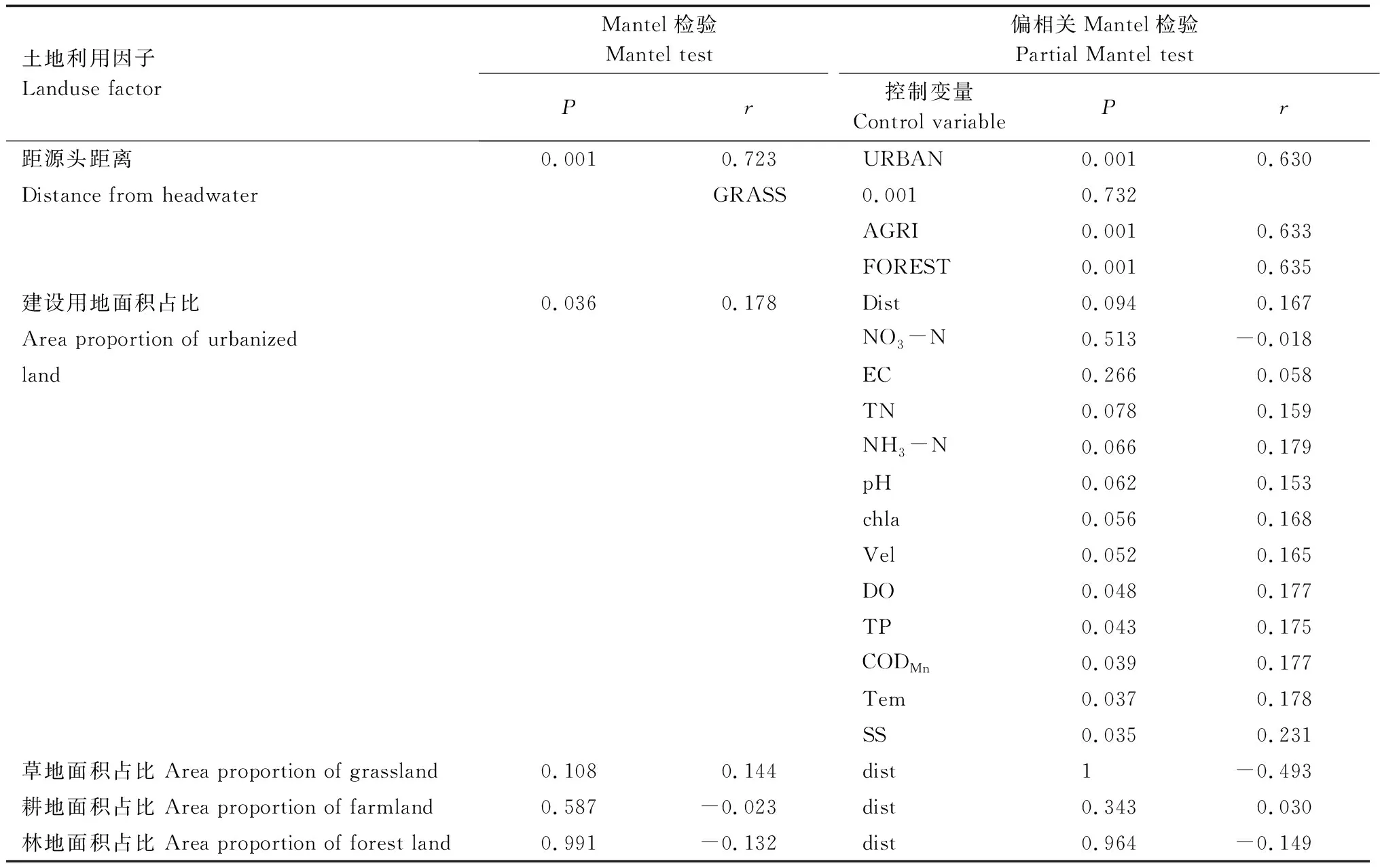
表5 群落Bray-Curtis距離矩陣與土地利用因子歐式距離矩陣的Mantel檢驗Table 5 Mantel test between community composition and land use factors
將溶解氧、總磷、高錳酸鹽指數和水溫作為控制變量后,建設用地占比與細菌群落組成依然存在顯著相關(P<0.05);將總氮、氨氮、pH、葉綠素a和流速作為控制變量后,建設用地占比與細菌群落組成在α=0.05的顯著性水平上不再相關,但在α=0.1的顯著性水平上依然相關(P<0.1);將硝酸鹽氮和電導率作為控制變量后,建設用地占比與細菌群落組成不再存在顯著相關。這說明在灤河干流中上游,建設用地可通過影響含氮物質排放,以及改變流速、pH等指標,對細菌群落組成產生影響。
3 討論
3.1 距河流源頭的距離對浮游細菌群落多樣性的影響
距源頭距離與灤河中上游浮游細菌群落的Bray-Curtis距離矩陣間的相關系數為各項水環境因子中最高(MantelR=0.723),且偏相關分析結果仍顯著。距源頭距離影響浮游細菌在河流中的滯留時間,細菌群落組成存在其獨特的影響[23]。在灤河干流上段,其主要的土地利用類型為自然程度較高的草原,向水體提供的細菌的多樣性也較高[24],而其中的核心群落一般來自于源頭附近的巖表生物膜或深層土壤[25]。隨著滯留時間的增加,從河岸帶周邊環境被徑流帶入水體的細菌之生態位,逐漸被能更好地適應淡水環境的核心群落細菌所取代。
此外,距源頭距離由于其數值上遞增的特點,與某些在河水流向下游過程中不斷積累的環境變量間存在某種相關性,例如各類非點源污染(氮、磷等),因此在一定程度上可以代表人類活動對河流水環境的擾動[8]。考慮到灤河中上游的中段的主要土地利用類型為林地,其生產力和能提供的外源性輸入量與草原相比較高;下段為承德市主城區,其城市規模較大,提供的外源性輸出更多。各種營養物質的累積,使得能高效利用養分的類群被選擇出來[22]。在這個過程中,浮游細菌群落由多樣性較高的近自然群落轉變為由靜水細菌占優勢地位的類湖泊群落[21,26]。在本研究中,這體現在沿河流方向,α-多樣性指數的降低和非典型淡水分類群占比降低,以及核心群落占比的升高[8,26—29]。
3.2 水環境因子對浮游細菌群落多樣性的影響
群落α-多樣性指數與氨氮、硝酸鹽氮、總氮、總磷、電導率、懸浮性固體、溶解氧間存在顯著的負相關關系。多樣性指數與營養鹽因子間呈顯著負相關,表明營養物質的增加可能導致了生態位分化,使得能夠高效利用營養鹽的類群比例逐漸上升而成為優勢類群;總懸浮固體可為水體中的一部分細菌提供定殖空間,灤河干流中上游的中段位于河谷地區,兩岸較為陡峭,水土流失情況比較嚴重,因此中段樣點的總懸浮固體量較上段和下段的多,這使得顆粒附著型細菌(particle-associated bacteria)的許多種類得以穩定存在并增殖,例如β-變形菌綱和擬桿菌門的細菌[27—28],而這些細菌也正是能高效利用營養鹽和藻類有機物的類群。
氮營養鹽對于河流浮游細菌群落結構的顯著影響已為多項研究所報道[30—32]。在以距源頭距離為控制變量后,各水質因子中只有以硝酸鹽氮、氨氮和總氮為代表的水體營養鹽因子依然與群落Bray-Curtis距離矩陣間存在顯著相關性(P<0.05)。這表明在灤河中上游,氮的排放變化與距源頭距離的增加并非完全一致,即并非自然狀態下的簡單累積過程,這很有可能是人類活動的空間異質性造成的。
3.3 河岸帶土地利用對浮游細菌群落多樣性的影響
Chao1豐富度指數是一種對偶見種較為敏感的多樣性指數[33],而Chao1豐富度指數在中端的波動,表明在河流的中段可能存在大量外源性的偶見種細菌輸入,而這些細菌在進入河流下段后又無法在種間競爭中勝出,因此在強烈的選擇作用下迅速消亡。外源細菌大量涌入的現象,可能與中段的各樣點處于地形和土地利用的過渡地帶有關。研究區的中段位于華北平原的丘陵地帶,與上游鹽堿化的壩上高原相比,在土壤類型、理化條件、植被類型等方面均存在較大差異,這些自然條件的差異會增加河流的其他外源性細菌輸入的可能性[34]。
此外,在研究區中段各樣點周邊的河岸帶緩沖區中,耕地和建設用地所占的比例與林地基本相當。已有研究表明,在農業土壤中,不同的作物種植、農藥和化肥的使用,以及機械化翻動土壤等做法,會改變土壤的理化性質,從而改變了土壤細菌群落及其生態功能這一變化使得農業土壤與近自然植被覆蓋下的土壤內的細菌群落結構間存在顯著差異,從而對隨地表徑流進入水體的外源性細菌輸入產生影響[35—36]。因此,河流中段樣點Chao1豐富度指數的異常升高,可以認為是自然條件差異以及土地利用類別發生變化后的綜合影響結果。
在將硝酸鹽氮和電導率作為控制變量后,建設用地占比的歐式距離矩陣與群落Bray-Curtis距離矩陣間的相關性不再顯著,這說明土地利用結構的改變對外源性營養輸入有著重要影響。在建設用地占比較多的地區,其郊區城鄉交錯帶種土地利用的變化劇烈[37],導致了含氮物質排放的增加。這對總氮輸出濃度和硝酸鹽氮輸出濃度有顯著源作用[38]。在灤河干流中上游沿岸,建設用地(包括旱地、居民地)的占比與距源頭距離并不存在線性關系,這說明城鎮的分布雖然有越向下游越多的趨勢,但其分布并不是完全成線性的,而這可能就是導致如上現象的原因。這也解釋了氮指標在以距源頭距離作為控制變量后,仍與浮游細菌群落結構間仍存在顯著相關的原因。
在將溶解氧、總磷、高錳酸鹽指數、溫度和總懸浮顆粒作為控制變量后,建設用地占比的歐式距離矩陣與群落Bray-Curtis距離矩陣間的相關性依然顯著(P<0.05),這表明建設用地與這些變量對群落結構的影響并不重合。在將距源頭距離、總氮、氨氮、pH、葉綠素a、流速作為控制變量后,建設用地占比的歐式距離矩陣與群落Bray-Curtis距離矩陣間在α=0.05的顯著性水平下不再呈顯著相關,但仍在α=0.1的顯著性水平下呈顯著相關,表明建設用地除了通過影響以上諸因子間接影響浮游細菌群落外,還可能通過其他效應影響著浮游細菌群落。
土地利用變化,特別是城市化會改變河流棲息地的局部物理條件,例如,使河流河段遭受泥沙侵蝕或沉積、電導率升高、溫度變化或其他環境脅迫[10—11]。此外,集水區的不透水地表占比(Watershed% IC)與城市化地區河流中的細菌群落關系密切,在土地利用結構向城市化轉變的過程中,地表不透水層的擴大切斷或改變了水源河流與地下水的供應,從而影響到細菌通過徑流進入河流[39]。不同類型的土地利用,由于其物質循環和能量流動的差異,也可能向水體輸出不同種類的菌種。與人畜病原體、腸道細菌和塑料降解功能等有關的菌種,會隨著一部分基礎設施(如流域內的滲漏管道)等被直接輸送到城市化程度高的河流中[40]。
4 結論
(1)灤河中上游的浮游細菌群落α-多樣性較高,細菌種類較為豐富。沿河流方向,β-變形菌綱和擬桿菌門相對豐度逐漸增加,放線菌門和γ-變形菌綱相對豐度保持穩定,其他門類細菌的相對豐度則隨距源頭距離的增加而逐漸降低。最終在下游形成了主要由β-變形菌綱、擬桿菌門、放線菌門占優勢地位的群落結構,與傳統研究中的湖泊細菌群落的組成類似。
(2)灤河中上游的浮游微生物群落組成受距源頭距離(營養滯留時間)、氮磷營養鹽的綜合影響。其中距源頭距離與細菌群落的是影響灤河中上游細菌群落結構的主要因素,很可能是伴隨著細菌滯水時間增加而隨之發生的群落演替過程。總氮、氨氮和硝酸鹽氮是影響灤河中上游群落結構的重要因素,且其分布存在空間異質性,這是由于建設用地的空間異質性分布導致的。
(3)建設用地占比對灤河中上游的浮游微生物群落組成存在顯著相關性。建設用地通過影響河流的水溫和理化指標,對浮游微生物群落產生復雜影響。該結果可為河流水生態保護和流域土地利用結構優化提供科學依據。
