深海生物原位實驗與生態監測研究進展
2022-08-15 02:57:16鄭鵬飛賀麗生高兆明李文莉張艾群
應用海洋學學報 2022年3期
王 勇,鄭鵬飛,賀麗生,李 俊,陳 俊,高兆明,李文莉,張艾群
(1.中國科學院深海科學與工程研究所, 海南 三亞 572000; 2.清華大學深圳國際研究生院, 廣東 深圳 518055)
1 深海生態系統特征
海洋占據了地球表面積的71%,包涵全球97%的水體[1],其廣闊的水體為生命的起源和各種生命形態的蓬勃發展提供了可能。深海通常是指水深超過1 000 m的區域,一般認為是一個黑暗、高靜水壓、低溫(約4 ℃,熱液除外)和寡營養的極端環境。在古希臘時期,人們認為深海不存在生命。19世紀的海洋科考不斷從深海獲取大量生物樣本,探明深海存在大量人類未知的生物且具有超出人類想象的生物多樣性。20世紀30年代,美國斯科利普斯海洋研究所(Scripps Institution of Oceanography)發現在海床下數厘米至數米的深海沉積物內存在細菌。自從美國“阿爾文號”深潛器在1977年發現了深海熱液生態系統,人們認識到深海不是“生命的荒漠”[2]。因此,現在及可預見的未來,深海生物具有重要的科研和經濟價值[3]。“新大洋科學發現計劃”(IODP,2012—2023年)將地球深部生命研究(包括深海)作為其四個研究主題之一。深海生態系統同樣由生產者和各級消費者構成。深海微生物參與碳、氮、硫等關鍵元素循環,連接地球深部和海洋表面的物質循環,是生物地球化學循環過程中重要的組成部分,為各級消費者提供營養物質,維持著深海生態系統的正常運轉。這些年新型科考船,載人/無人深潛器等先進裝備的發展為人類了解深海提供了有力的載體和工具,擴展了人類探索深海的邊界,但是目前對深海生態系統和生命過程的一些基礎問題仍然難以回答,例如:①深海有機物的來源和主要貢獻者;②深海生態系統的時空動態變化規律和調控機制;③深海生物原位活性和生態功能。對這些問題的探索,有利于我們深刻理解深海生態系統的動態性、深海生命的生理生化狀態和物種的演化過程,同時為下一步的研究指明方向。
1.1 深海有機物的來源和主要貢獻者
在深海,生物的分布受到營養來源、溫度等各種環境因子的驅動。一般認為海洋表層初級生產力產生的大量有機質的垂直沉降是深海最重要的有機物來源[4],但是這些有機質是否足夠支持深海遠超我們想象的大量生物和生態系統呢?[5]有研究發現在3 000 m的深海區,維系生態系統的有機物只有約1%來源于海洋表層的有機顆粒物(particulate organic carbon, POC) 沉降[6]。那么,深海有機碳的主要來源是什么?化能自養微生物貢獻了多大比例?目前我們對此仍然知之甚少。大量研究也發現微生物在海洋垂直方向不同深度的水層具有迥然不同的分布特征[3]。據估算,海水中原核微生物細胞含量約為1.2×1029個/mL[7],而黑暗的深海水體中生活著約占全球生物總量65%的微生物。最近的研究表明,二氧化碳不但在海洋表面被光合微生物吸收,也在深海無光層中被水體中的微生物利用于合成有機碳[8]。據估計,大西洋深海無機碳固定的速率為每天1.0~2.5 mmol/m2,占真光層(0~200 m)匯入的有機碳的15%~53%[9]。這些由深海化能微生物提供的新鮮有機物支撐了深海生態系統的正常運作,也解釋了深海生物在寡營養條件下的生存之謎。傳統認為深海的碳固定與氨氧化過程密切相關,因為二氧化碳固定的主要承擔者可能是氨氧化古菌(ammonia-oxidizing archaea, AOA)。AOA利用氨的有氧氧化產生的還原力,然后通過3-羥基丙酸/4-羥基丁酸循環(3-hydroxypropionate/4-hydroxybutyrate cycle, 3-HP/4-HB cycle)途徑吸收和固定二氧化碳。AOA將氨轉化成亞硝酸鹽,而后被亞硝酸鹽氧化細菌(nitrospirae和nitrospinae)氧化成硝酸鹽。由于氨氧化古菌在海洋中的數量高于亞硝酸氧化菌(nitrite oxidizing bacteria, NOB),傳統認為前者固定的二氧化碳通量高于后者。但是,最近的原位碳固定實驗發現伴隨亞硝酸氧化過程,NOB固定的二氧化碳通量明顯高于AOA,最高占深海固碳量的45%[10]。這些參與硝化過程的自養微生物占深海微生物群落不足10%,豐度低于AOA,但是卻充當著深海初始生產者的重要角色。近期的研究發現屬于變形菌綱的一些種屬,比如SAR324,可以利用核酮糖1,5-二磷酸羧酶/加氧酶和硫化物氧化耦合來固定二氧化碳[11],說明依賴于硝化作用和硫氧化作用的化能自養型和混合營養型微生物在深海生物地球化學循環過程中也發揮著重要作用,被認為是深海有機碳來源的另一重要途徑。這些深海自養菌和兼性自養菌可以吸收和利用異養型微生物產生的二氧化碳,形成深海內部的碳循環,同時也可以吸收大量經過海水垂向循環輸送到深海的二氧化碳。深海異養型微生物以變形桿菌和綠彎菌為主,代表了深海最豐富的微生物類群,參與降解難降解(recalcitrant)有機質(dissolved organic matter, DOM)并釋放二氧化碳[12]。從深淵原位轉錄組研究獲知,綠彎菌門的SAR202是深海和深淵最活躍的微生物,對于有機物的降解和再利用起到至關重要的作用[13]。深海細菌SAR11和 SAR406,可能參與有機物降解和硫元素循環[14]。在冷泉和熱液噴口附近,來自地球深部和沉積物底部埋藏的水合物釋放了大量還原性物質(如氫氣、甲烷和硫化氫等)。這些物質的擴散導致化能自養微生物介導的有機物合成異常活躍[15],形成了局部繁盛的深海生態系統。在大型生物體內[如貽貝(Bathymodioelusplatifrons)和管蟲(Riftiapachyptila)]也含有大量利用還原性物質進行固碳的共生微生物[16]。深海微生物利用深海還原性物質固定二氧化碳能力為貧瘠的深海提供了豐富的營養來源,同時提示深海具有巨大的碳埋藏潛力,對實現全球碳中和目標有著重大的應用前景。因此,對深海固碳類群和固碳過程的精細量化和調控機制的探索,對進一步全面理解深海固碳過程有重要意義。
1.2 深海生物時空分布規律
幾乎所有的有機體,從單細胞的藍藻到結構復雜的哺乳動物,都具有一套感知環境變化的生物節律系統,控制著機體的行為、生理活動,使之更好的適應環境[17]。由于缺乏相關的研究,目前人們認為深海是不存在類似的節律性。但是最近對深海冷泉貽貝相關基因的研究表明,深海生物具有節律性的生物學基礎,可能受到深海潮汐的影響[18]。
在海洋表層,光照的晝夜變化直接調節了初級生產力的速率,構建了海洋表層生物的晝夜節律。但在深海,海洋表面的初級生產力對超過3 000 m的深海影響輕微。但長期來看,由于沉降帶來的有機物和宏生物(如魚類)垂直周期洄游活動帶來的有機物,使深海間接受到海洋表面光合生產力的影響。已有研究發現,深海微生物受到大洋環流等環境變化影響,存在明顯的季節性變化[19-21],但對深海微生物晝夜節律變化鮮有報道[18]。對中國南海北坡海洋中層帶魚類棲息環境中微生物群落結構研究表明,由于受到海洋魚類垂直周期洄游活動的影響,該魚類棲息環境中微生物群落結構也具有明顯的晝夜群落結構變化[22]。影響深海生態系統的節律因素必然與能量轉換和物質循環密切相關,因此探究深海生物的節律規律對進一步理解深海物質循環有重要意義。研究在深海不同層次和生境條件下,生物進化出何種節律特征,需要對深海進行大范圍和連續時空的生態觀測數據收集和統計分析。而目前主流的研究方法和手段只能獲得大洋某個深度和時間點的觀測數據,無法滿足我們對深海生態系統的動態規律研究。
2 深海生態系統原位觀測
2.1 深海研究的挑戰和技術瓶頸
人類對海洋的研究雖然經歷了一個多世紀的堅持不懈的努力,但我們對深海生態系統的理解仍然不完整。國際上,“TARA”和國際黃昏帶研究計劃(joint exploration of the twilight zone ocean network, JETZON)計劃完成了對全球海洋0~1 000 m的生物調查和相關環境因子的采集,實現了對不同大洋站位的海洋表面和黃昏帶的覆蓋[23-24]。中國科學家已經完成了針對挑戰者深淵的大量生物學調查和研究[25-27]。目前對深海生態系統的研究和調查正在從某個時間點在單個站位取樣進行研究,逐漸過渡到連續時空觀測階段。然而在深海實現對不同站位和時間季節的連續觀測和調查難度極高;深達萬米的高靜水壓對裝置的耐壓性能和自動化程度要求甚高;長時間連續觀測對裝置的能源供給和水下通訊提出了巨大的考驗。這些瓶頸問題的解決需要深海工程技術方面的配合和同步推進,而目前相關技術的研發基礎較為薄弱,限制了海洋學家對深海生物和生態系統的進一步探索。進入新世紀后,伴隨著國家深海戰略發展的需要,相關深海原位監測技術取得了長足的進步。比如,萬米載人深潛器,達到了世界先進水平,給我國科學家提供了在萬米深海長時間直接觀察深海生物的機會。
雖然人們認識到深海生物/微生物巨大的應用和科研價值,然而深海生物由于其獨特的生存環境,使得對其取樣、培養和研究整個過程都存在極大挑戰。深海的有機物相對匱乏,使得深海生物進化出各種探測環境變化的器官和胞外傳感系統[28]。深海生物,尤其是微生物對環境因素變化(氧含量變化、壓力變化等)的響應十分敏銳,相關表達系統可以快速響應并觸發相關功能基因的表達[29],尤其是涉及有機物合成必需的二氧化碳、甲烷和氨氣等。因此,通過對這些相關基因表達變化進行檢測,可以以此作為衡量深海環境變化的天然指標。然而傳統采樣使用水樣瓶將深海海水帶到船載實驗室過濾,這一過程中的壓力、溫度和氣體成分發生明顯改變,進而導致部分菌群快速增殖和發生應激性改變[29-30],使得原位基因表達信息丟失(圖1)。這樣的樣品在后續的實驗室研究中會產生很多不合理的假象,因而影響對生物學上相關指標(DNA、RNA和蛋白質等物質的含量與組成,及各種功能基因的量化指標)的精確測量。傳統采樣方式對深海生物樣品影響較大,限制了對深海極端環境下生命過程和適應機制的研究,甚至帶來錯誤的認知。其次,深海微生物所處環境的特殊性使在實驗室實現高仿真模擬培養變得幾乎不可能。
盡管利用深海環境模擬進行生物培養工作難度較大,但仍然是國際上微生物地球化學研究領域的一種重要方法。 在深海微生物培養系統研制方面比較先進的主要有美國明尼蘇達大學地質地球物理系、加州大學伯克利分校,伍茲霍爾研究所(Woods Hole Oceanographic Institution)以及日本海洋科學研究中心(Japan Agency for Marine-Earth Science and Technology)。國內在實驗室模擬培養方面的研究起步較晚。這些可培養微生物在實驗室條件下,由于生存條件改變,連續的傳代培養會產生“馴化效果”。培養基中添加有機碳源最終會導致化能自養菌基因組漸漸發生變化,出現異養性基因橫向轉移,變成兼性菌。因此,實驗室培養菌株很難真實反映其原位生境的真實代謝狀況。模擬培養可以解決微生物基礎生理特性等問題,但無法完成對微生物原位生態功能和動態變化的監測和評估。因此,深海原位研究在生態學上有更重要的意義。浙江大學等相關單位合作成功研制了我國首個深海水體原位微生物培養系統,由控制室、培養室、浮力材等組成。國家海洋局與杭州電子科技大學聯合研制的一套能夠在深海原位環境下獨立開展工作的深海原位生物實驗平臺,主體由16個培養倉、原位環境監測傳感器、聲學釋放器、浮力材料和重力錨等組成[31-32]。系統設計最大工作水深6 000 m,可布放到海水與沉積物界面處,進行深海微生物的原位長期培養。
隨著深海宏生物采樣和觀測手段的進步,依靠國內外不斷增加的海洋科考航次,我們對深海生態系統的認識在不斷加深和完善。由于技術條件限制,對深海宏生物的早期研究主要使用拖網等傳統方法,但捕獲的生物個體少且多數情況下難以獲得完整樣本。近年來依靠載人潛器和深潛機器人的發展,研究人員獲得高質量的生物樣品和原位實證資料(照片、視頻等),使人類對深海生態系統的基本構成和分布規律有了直觀的初步了解。例如,科學家發現,深海宏生物遠比想象的豐富,深海獅子魚最深分布于8 250 m左右,而鉤蝦和海參可生活在挑戰者深淵的最深處。冷泉附近聚集了大量的甲殼類和管蟲等生物[33]。由于深海與科考船或岸上巨大的靜水壓差,多數生物從深海到實驗室發生明顯的形態變化(圖1)。具體而言,對多數宏生物,巨大的氣壓差異,使細胞和組織發生爆裂,無法還原樣品原位環境下功能基因的表達信息和蛋白活性特征[34-36];海參等生物會發生自融;深淵獅子魚柔軟的表層皮膚會剝離(圖1)。
總之,盡管深海生物調查與研究已經開展了多年,目前所有深海典型生境的生態學研究結果也只能反映某個時間點的靜態狀況,對于深海生物的繁殖、進化、生理特性、種群演替和生態系統(種群間)物質能量傳遞交換知之甚少。實驗室難以模擬深海生物生長所占據的環境獨特特征(高靜水壓、獨特的營養構成)。為了能夠更加準確、全面的認知深海環境中獨特的生態系統和生命過程,進行深海生物原位光學成像觀測和功能基因檢測幾乎是唯一可行的、有效的、高保真的技術手段。
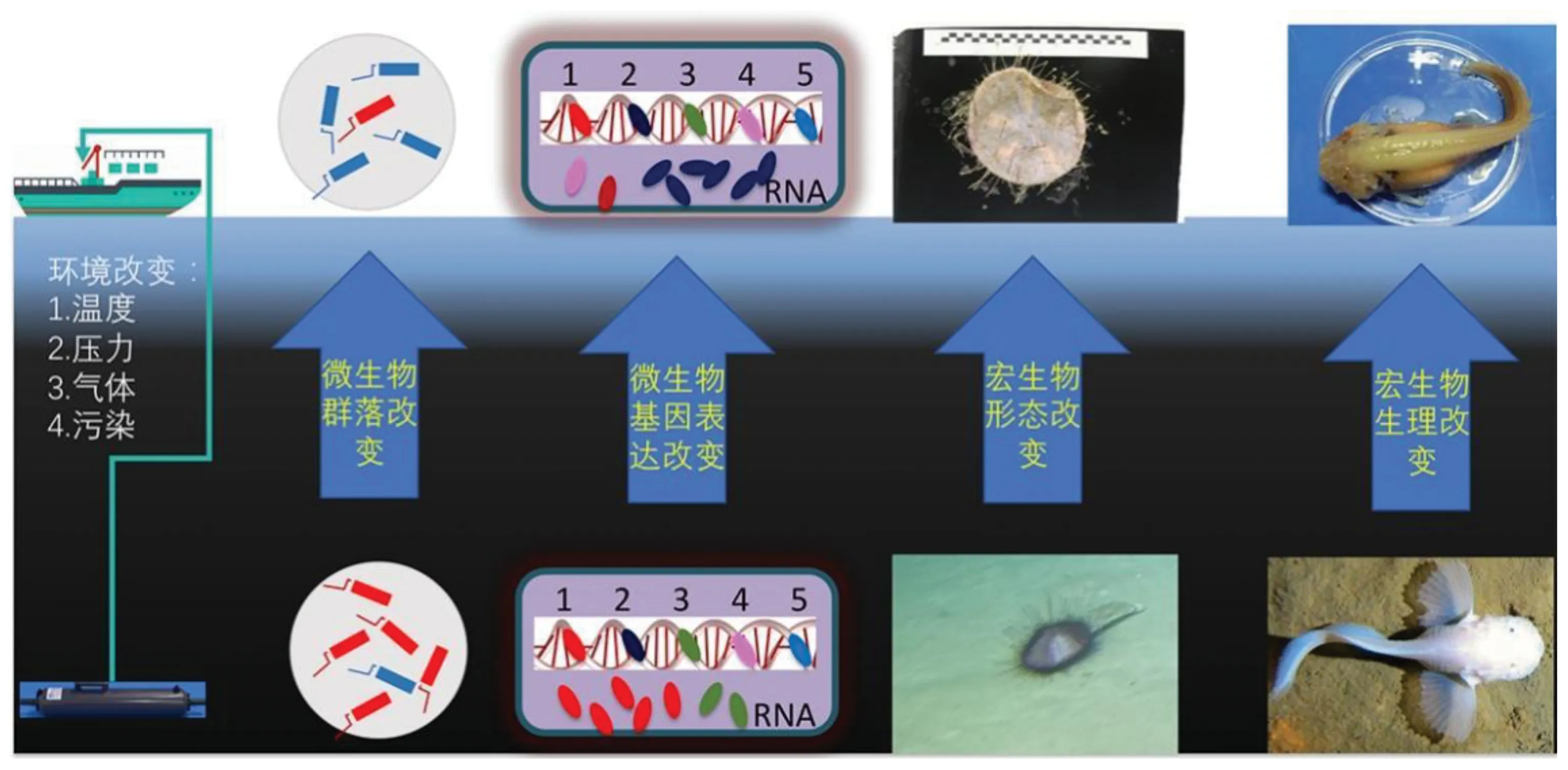
圖1 傳統深海生物采樣過程對深海生物樣品的潛在影響Fig. 1 Potential impacts of traditional deep-sea biological sampling process on deep-sea biological specimens
2.2 深海原位生物研究策略和裝備研發
深海生物的新陳代謝、適應性和生物多樣性是國內外海洋研究的前沿課題。由于生態脅迫作用, 深海生物在適應深海的極端環境過程中,必定產生了有著特殊功能的基因、蛋白和次級代謝產物。為深入研究深海生態系統構成、演化和功能等重要科學問題,我們急需新的研究平臺和技術,其中深海原位生物研究和探測是重點突破方向,它涵蓋了深海原位光學成像,理化因素檢測,生物原位采樣和基因活性測定等,將引領深海生命科學研究跨入一個新階段。
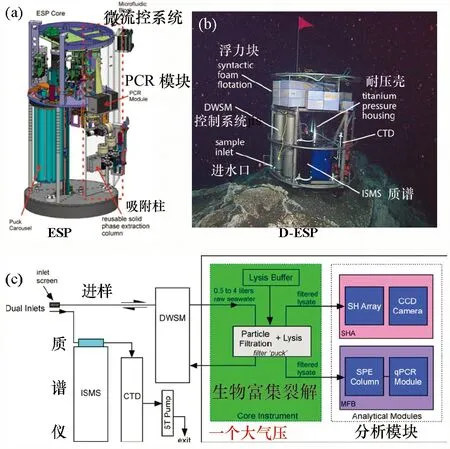
圖2 美國蒙特利灣研究所的環境樣品處理器和深海升級版D-ESPFig. 2 ESP and D-ESP from MBARI(a)環境樣品處理器;(b)、(c)深海升級版D-ESP。修改自文獻[39]和[40]。
盡管國內外已經開發了一些深海微生物原位保壓取樣裝置,但由于保壓取樣器容量小、不保溫等因素,深海特有基因和生命過程的研究仍無法進行[37-38]。2007年,美國蒙特利灣水族館研究所(Monterey Bay Aquarium Research Institute,MBARI)和伍茲霍爾海洋研究所實驗室的海洋學家研發了1 000 m級環境樣品處理器(environmental sample processor, ESP)[39],對海洋微生物和毒性物質進行了原位基因芯片檢測。2010年,他們還成功地利用4 000 m級環境樣品處理器(D-ESP)進行了深海甲烷氧化基因pmoA的定量PCR擴增(qPCR)實驗,對目標基因的表達量和特定菌群的豐度進行了實時檢測[40],而且利用生物基因探測到的甲烷濃度低于化學傳感器的檢測限。這些原位基因檢測都是依賴原位微生物的富集、過濾、裂解和核酸純化過程[圖2(a)、(c)]。原位過濾后的海水中的物質還通過D-ESP集成的深海質譜儀進行了檢測。這一整套裝置已經應用于4 000 m深的海底和800 m深的甲烷滲漏區的原位檢測,并接入MBARI的MARS(Monterey Accelerated Research System)海底觀測網絡。2014年,美國夏威夷大學利用ESP系統發現了海洋表面微生物存在短周期節律性過程[17,40-41]。總的來說,ESP的相關成果極大促進了科學家對深海原位觀測的重視[40,42]。
2.3 深海原位組學研究進展
深海原位組學是使用新一代測序技術對深海原位提取核酸和固定樣品進行高通量測序和組學分析,為全面準確地揭示深海微生物在深海的適應機制、物質循環和動態變化規律提供了可行性[13,36]。深海生物轉錄組的瓶頸問題是RNA分子容易降解,半衰期最多只有12 min[30]。對RNA分子進行原位固定或者提取,防止生物在回收過程中RNA出現變化和降解是研究深海生物原位活性和生理代謝特征的前提。通過利用深海原位提取的RNA進行宏轉錄組分析,我們可以在轉錄層面定量或半定量地揭示哪些微生物以何種方式參與深海碳、氮、硫循環,它們的貢獻率是多少,能夠更精細地量化深海物質循環過程。同時,深海的有機物相對匱乏,使得深海微生物進化出各種探測環境變化的跨膜傳感系統,對環境的響應十分敏銳[29]。原位宏轉錄組學或許能捕捉到深海微生物對環境響應的即時變化。為了準確地探知深海生物功能基因的表達和作用機制,發掘深海生物的未知功能基因,原位生物樣品處理與探測關鍵技術研發勢在必行。
深海原位宏轉錄組數據分析離不開相應的深海宏基因組作為本底參照。后者利用高通量測序和生物信息學方法對深海環境樣品DNA直接測序,通過組裝和分揀,獲得同一類群微生物的基因組草圖(metagenome-assembled genome, MAG),進而對其MAG的分類地位、新陳代謝方式進行解析。目前由于大多數微生物不可培養,對基因組草圖的研究是深海微生物研究的主要手段[43]。通過這種方法,近些年來報告了大量海洋廣泛分布的氨氧化古菌SAR202和 SAR406等類群的基因組及其特征[13-14,44-45]。通過相關算法,可以把宏轉錄組的表達譜和宏基因組中獲得的MAG聯系起來,進而揭示深海原位微生物類群中活躍的微生物,同時可以定量或半定量研究微生物的代謝過程。然而,深海原位宏基因組研究難點在于復雜生態環境中難以分離到高質量的MAG,其主要原因是目前技術手段得到的總DNA量較低且質量不高,并且群落復雜。這樣的深海DNA可通過加大二代高通量Illumina測序深度等手段進行補救,但難以用于三代高通量測序獲得長片段基因組序列。這一瓶頸問題可以通過深海原位大體積微生物過濾和固定裝置解決。中國科學院深海科學與工程研究所(深海所)自主研發了一套可在全海深環境下自動完成海水中微生物過濾和RNA固定的裝置(insitumicrobial filtration and fixation, ISMIFF),可以最大程度上避免了壓力和溫度等環境因子驟變對核酸的破壞[36]。
深海微生物原位核酸提取裝置(multipleinsitunucleic acid collections, MISNAC)是由深海所自主研發的一套可在深海環境下自動完成海水過濾、微生物裂解和核酸提取的深海原位生物實驗裝置[46][圖3(a)、(b)]。該裝置安裝于“鳳凰”號深海原位生物研究平臺上,最多可以完成9個不同時間序列的核酸樣品采集,而且所有步驟均自動完成[圖3(c)至(e)]。該平臺不僅可以用于研究不同時間段微生物群落變化,還可以開展微生物定向富集培養,研究不同底物作用下的原位微生物生理代謝活性。
2019年的“鳳凰”號海試中,MISNAC獲得的連續時段微生物核酸揭示了深海1 000 m水深的微生物群落結構[46]。最近,基于原位獲得的核酸進行的組學研究發現:相對于傳統采樣方式使用的Niskin瓶,MISNAC可以最大程度地還原深海原位的群落結構和原位微生物基因表達活性,可以對連續晝夜條件下的微生物動態變化進行深入解析。通過對MISNAC裝置和Niskin水樣瓶獲得樣品的比較發現,真核和原核微生物群落結構都具有明顯差異。Niskin采水瓶樣品中含有大量MISNAC原位樣品中很少的厚壁菌(firmicutes), 且在轉錄組中沒有活性,表示這些菌在大量增殖后變成了休眠的孢子[圖4(a)];Niskin水樣瓶樣品還含有MISNAC原位樣品中很少出現的子囊菌門(Ascomycota)和刺胞動物門(Cnidaria)。 通過比較原位宏基因組和宏轉錄組中的真核生物群落,我們發現MISNAC樣品中較少的纖毛亞門(Ciliophora),卻是轉錄組中最活躍的,而占真核微生物群落42%的有孔蟲門(Reraria)幾乎沒有轉錄活性,暗示這些深海的有孔蟲是休眠或死掉的生物[圖4(b)]。這些結果在不同時間段的樣品分析中重復出現,可信度很高,說明基于深海宏基因組的相關分析很可能無法真實反映深海生物原位的生態貢獻。
MISNAC可以對數天內若干個小的時間間隔(幾個小時到幾十分鐘)的微生物的基因表達和群落變化進行連續、比較精確的定量或半定量研究。在更長的時間尺度上,深海原位研究可聯合物理、化學傳感器提供的長期連續觀測數據,基于各種算法挖掘出與微生物的群落或微生物參與的某些代謝特征的相關痕跡,即通過多參數進行數據耦合,才能實現對某目標海域微生物介導的物質循環進行連續動態觀測和量化分析。中科院南海海洋研究所已經完成了“深海紫外激光微生物檢測儀”的研制和海試,通過對不同深海細菌的紫外光譜可以辨別出主要深海類群的特征譜峰[47],可以實現較長時間范圍內對微生物主要類群的檢測。
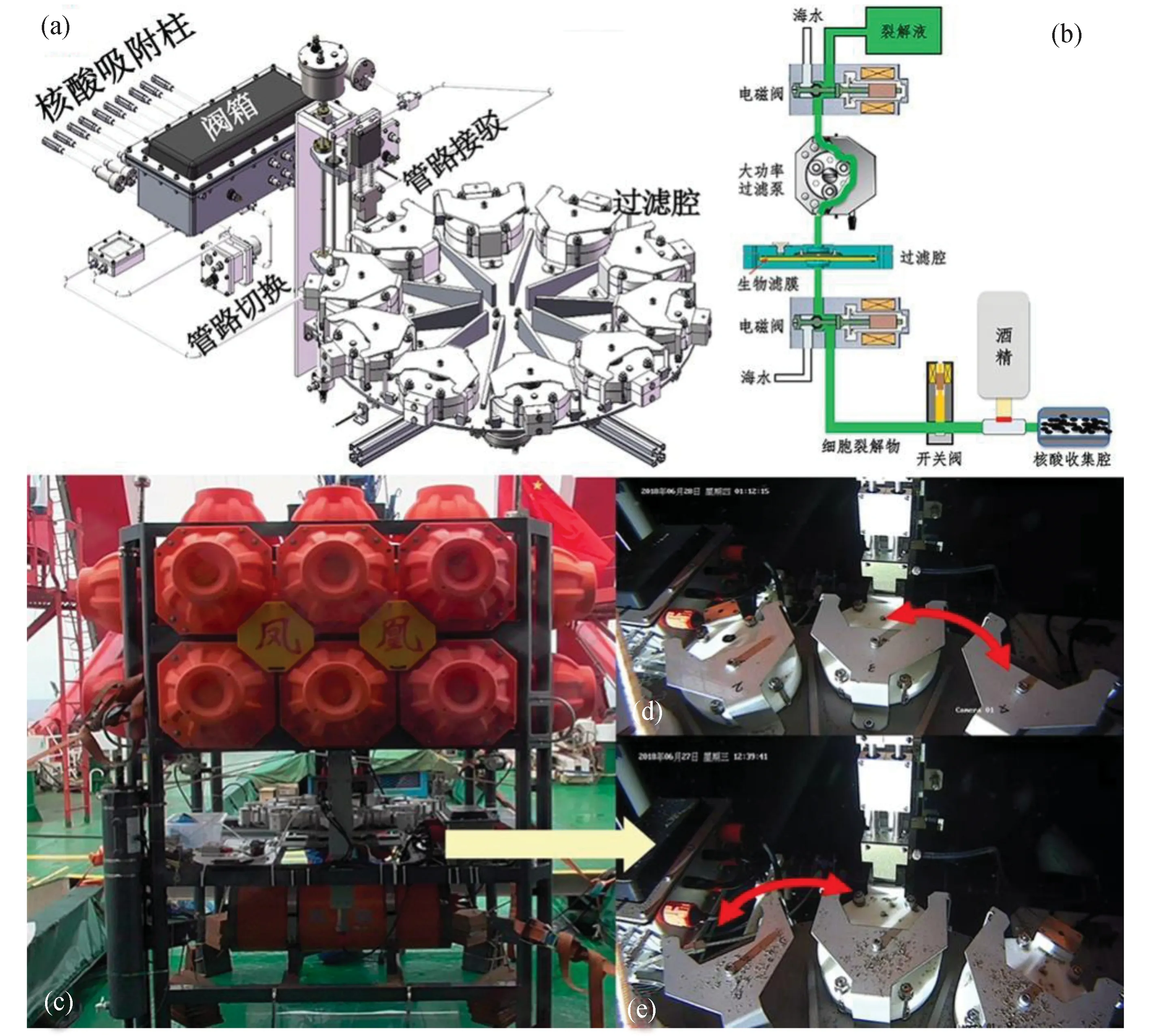
圖3 深海原位微生物核酸提取裝置工作原理圖和安裝于“鳳凰”號的深海原位生物實驗平臺的深海原位工作圖Fig. 3 Schematic model of MISNAC and in situ working in cruise on Phoenix deep-sea biology experimental station(a)、(b)為工作原理圖;(c)至(e)為深海原位工作圖。
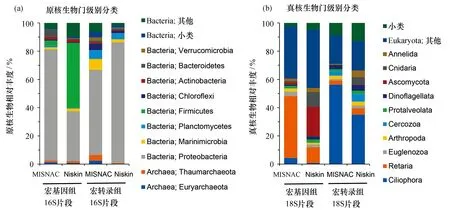
圖4 不同采樣方式對群落結構的影響Fig. 4 Impacts of sampling methods on microbial community structureMISNAC和Niskin獲得的南海1 000 m水深環境樣品的宏基因組和宏轉錄組中基于原核生物16S核糖體(a)和真核生物18S核糖體片段(miTags)(b)的群落結構比較分析。小類(<2%)進行了合并處理。
2.4 深海原位生態監測和原位生物實驗室構想
對深海進行原位、實時和連續的生態監測是未來揭示深海生物適應機制和深海物質循環的重要手段,各國深海科學研究機構都開始致力于探索和發展深海原位技術。原位生物實驗室,簡單來說,就是能夠在深海自動完成原位光學成像觀測、樣品采集、分離、處理和檢測等流程的綜合實驗體系。它包括了光學成像系統、實驗分析測試系統和支撐實驗室長期連續工作的數據傳輸、能源、電氣系統等(圖5),可以記錄樣品采集和實驗過程中的環境要素信息(溫度、鹽度和溶解氧等),把微生物樣品分離成單細胞,或者直接裂解后提取高純度核酸和蛋白質,甚至得到基因組測序數據和基因定量PCR數據。實驗室所需裝備(比如通用測序系統、基因檢測系統、化學物質檢測系統)的深海轉移應用需要對其進行改裝,使其小型化和低能耗,適合深海高壓環境下自動化作業和數據自行處理。同時利用聲學、激光等海洋通訊技術,結合船載和衛星通訊技術,可以將觀測數據和實驗數據傳回到岸基數據中心。利用高機動載體構建深海原位生物實驗室水下網絡,實現深海原位生物實驗室數據實時化、在線化。
2.4.1 深海實時生態觀測及數據傳輸系統 利用智能深海平臺的長期連續工作能力,深海生物小型化實時光學觀測裝置可以用于深海各深度的生態觀測和統計。深海影像技術是深海原位生物實驗室的重要應用領域,可開展遠程監測、遠程直播,甚至遠程實驗操控等應用前景。1992年美國伍茲霍爾海洋研究所提出了浮游生物錄像機(video plankton recorder,VPR),成為了現代浮游生物原位探測系統的先驅。伴隨著半導體技術的突飛猛進,浮游動物水下攝像機探測技術得到迅速發展,出現了多種基于水下攝像機的浮游動物探測成像系統,如UVP(underwater vision profiler)、VPRⅠ/Ⅱ/Ⅲ以及ISIIS(insituichthyoplankton imging system)等[48]。
2015年2月,歐盟UTOFIA (underwater time of flight image acquisition)計劃啟動了用于漁業資源評估的“下一代水下攝像機”的研制工作,由歐洲不同國家的7個單位參與。UTOFIA攝像機具備視頻幀頻三維成像功能,相比于傳統水下成像,探測距離提高2~3倍。由中國科學院半導體研究所研制的深海原位生物三維成像系統“鳳眼”, 以“鳳凰”號為載體,可在水下構建非機械光立體采樣區,并同時獲取高分辨率的生物二維強度圖像和三維圖形圖像,從而利用三維圖像中目標間的空間差異,提高交疊目標的數量統計準確度,結合光立體采樣區的體積信息還可反演出生物豐度信息等[49]。“鳳眼”系統最大工作深度3 000 m,是國際上首次將激光選通三維成像技術用于深海宏生物探測。
利用海底觀測網和海底光電纜可以實現我國南海從淺海到4 000 m深海的實時播放,對研究我國南海海域的生態系統特征、冷泉區演變過程以及海底生態年際變化等重大科學問題提供數據和樣品方面的需求[50-51]。2020年11月,中國科學院等多家機構聯合研制的“奮斗者”載人潛水器和深海視頻著陸器“滄海”和“凌云”在馬里亞納海溝進行了聯合作業,實現了全海深4K超高清視頻拍攝傳輸和全球首次萬米海底電視直播,對于實現深海生態的長期原位觀測驗證了技術上的可行性。
2.4.2 深海生物樣品處理和通用測序系統 隨著基因測序技術的發展,以納米孔測序為主的高通量且小型化的新一代單分子測序平臺逐漸成為主流,為其深海原位應用提供了可操作性。在已有的深海原位樣品富集、細胞裂解和核酸收集裝置的基礎上,我們可以進一步擴展下游核酸純化技術,整合實現深海原位條件下的一體化測序平臺。同時,在平臺上游,配套利用流式細胞儀對深海微生物和幼蟲進行原位分揀,亦或通過微液滴包埋分揀技術進行微生物高通量化的分揀和原位培養,實現對深海生物的隔離檢測,提高檢測的定向性(圖5)。
2.4.3 深海生物原位基因檢測系統 深海原位基因檢測系統是通過構建深海生物保守的核糖體 RNA基因片段和深海微生物物質循環關鍵功能基因(氨氧化、反硝化、產甲烷和硫還原等)的探針序列,集成高通量基因芯片技術作為深海生物原位核酸純化裝置的下游配套設備,實現不經測序的基因檢測和物種鑒定,以實時動態監測深海原位微生物的類群組成和功能基因代謝活性。
2.4.4 深海生物代謝物檢測系統 深海的高靜水壓給生命相關物質的檢測構成了巨大的挑戰。拉曼光譜是研究生命相關物質的有效方法之一,其通過對分子官能團震動模式的測量,得到具有指紋特性的震動光譜,從而反映分子的精細結構,可在接近自然狀態下研究生物大分子的結構及其變化。利用紫外拉曼和電化學傳感器等新興的物質檢測技術,建立深海生物代謝物質數據庫,將設備封裝應用于深海,實現原位條件下實時的代謝物質檢測,以利于還原深海生物真實的物質代謝類別和水平,整合利用多波段的激光檢測儀器可實現物質間的交差檢測。
近些年,我國多個科研團隊基于該技術在深海化學物質探測上取得了突破。中國科學院海洋研究所利用我國自主研發的深海激光拉曼光譜原位探測系統在深海熱液區(1 400 m)發現了具有超臨界二氧化碳流體噴發的熱液噴口[52],同時借助該工具首次在深海熱液區發現氣態水[53]。“深海原位探測深紫外激光拉曼光譜儀”由中國科學院大連化學物理研究所自主研發,在深海原位環境下深紫外拉曼光譜儀器完美解決了常規拉曼光譜易受海洋微生物及有機質熒光干擾的問題,并克服了深海高壓和著陸沖擊等困難,成功于南海獲得DNA分子信號,為進一步探究深海環境中存在的生物大分子信息提供了新的途徑。深海生物細胞內物質的原位紫外拉曼檢測方法和深海培養菌株的拉曼光譜數據庫也逐漸成熟。
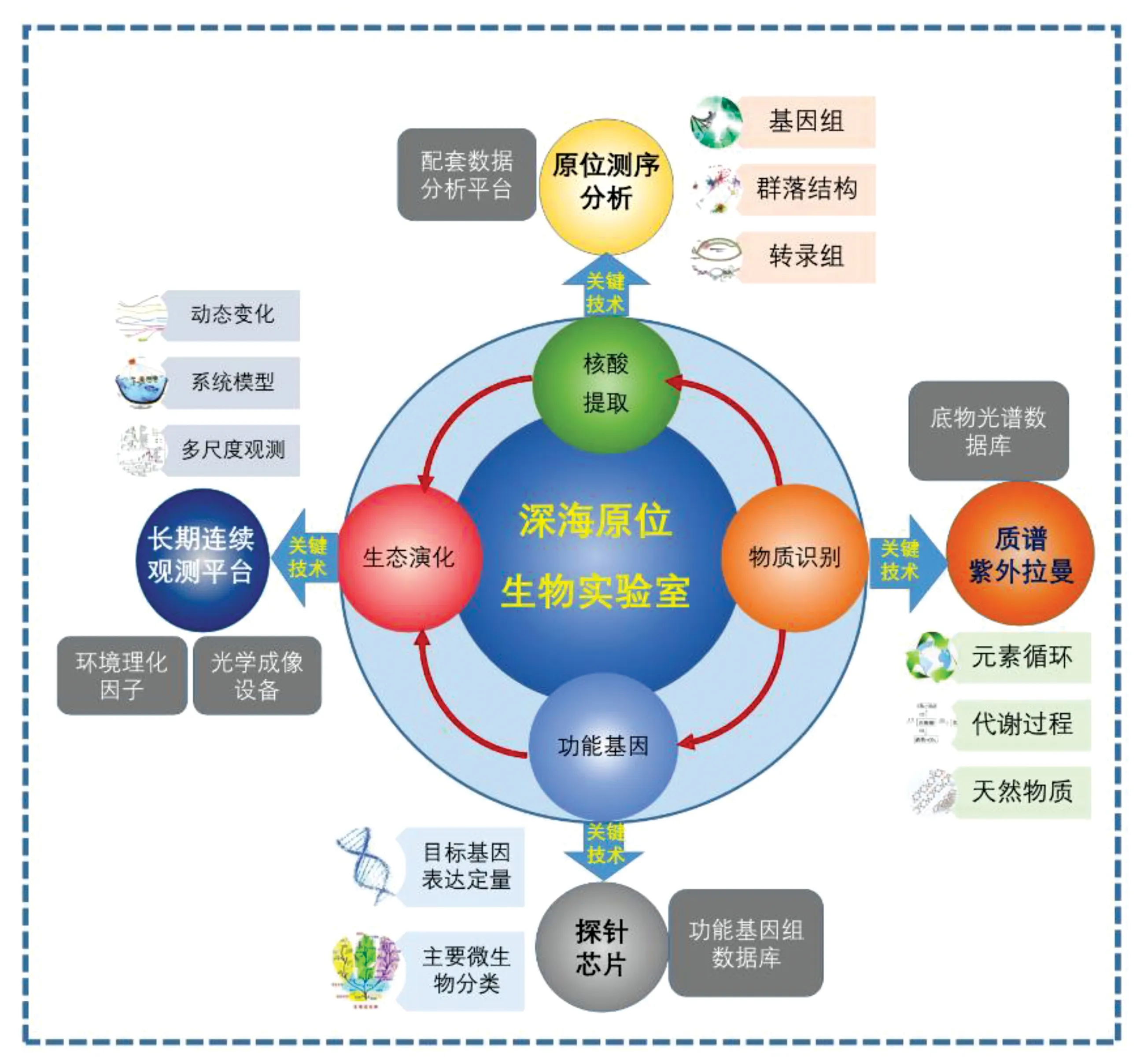
圖5 深海原位生物實驗室設想Fig. 5 Concept of deep-sea in situ biological laboratory
3 深海原位生物實驗室的應用前景和展望
3.1 深海原位微生物固碳與碳中和
針對地球二氧化碳濃度不斷升高,全球提出了“碳中和(carbon neutrality)”目標,這是本世紀中葉各個國家重要的控制目標。其中對碳匯研究方面,即碳固定方面,深海微生物豐富高效的碳固定途徑已經引起很多研究者的注意。深海1 000 m以下的微生物在全球碳吸收和控制溫室氣體排放方面可能具有不可估量的作用,但是深海固碳能力的評估和實現途徑依賴于人們對深海的認識,其中深海原位微生物無機碳吸收過程和量化的研究尚處于起步階段。
Jiao等(2010)提出的“微型生物碳泵(microbial carbon pump, MCP)指明了深海碳循環的可能機制[8],解釋了海洋巨大溶解有機碳庫的來源,得到國際同行的廣泛關注和認同。 為了響應國家的“碳中和”計劃,基于MCP理論的“海洋負排放”機制和技術實現是目前的研究熱點[54-55],但是目前對海洋儲碳機制還有很多問題需要研究,尤其是深海微生物原位固碳能力和機制資料較少[4],很多基本問題尚未解答,例如深海參與固碳的微生物有哪些?通量是多少?其代謝機制受到哪些因素調控?氨氧化古菌和細菌在深海固碳過程中,互作機制是什么?二氧化碳如何輸送到深海?這些問題的回答都需要長期、多學科交叉的持續研究。這些問題都需要深海原位生物實驗室提供長期、連續和第一手的資料為“碳中和”的“海洋負排放”機制和技術可行性、經濟性和可選路徑提供最基礎的數據支撐。這些凸顯出深海原位生物實驗室研發的重要性和必要性。
3.2 深海基因資源的挖掘和利用
深海環境原位宏基因組和宏轉錄組數據的獲得,將有助于我們發現更多新的深海微生物類群和深入探究深海“暗物質”微生物的生理特征[56]。同時深海微生物生命過程的原位觀測,有助于深入探討海洋環境中微生物的生態功能,精細量化其物質通量,揭示海洋環境物質循環和能量流動的分子機制和規律。隨著生物信息學分析、三代測序技術及分子生物學技術的發展,結合新型原位實驗設備的研制,我們不僅能通過分離培養獲得更多的深海微生物類群,還可以利用分子生物學技術,通過過表達細胞內目的基因、抑制和敲除等分子手段,探究大量未知基因的功能,挖掘和利用深海生物基因資源。
3.3 推動海洋高端裝備產業
工程技術與生物學研究的需求相結合將會產生更多功能強大的采樣設備。相關設備的研制可以帶動我國深海原位檢測和研究設備的研發和關鍵技術的突破。例如我國萬米載人深潛器“奮斗者”號的研制成功,不僅實現了相關產業和技術的國產化替代,同時培養了一大批相關產業技術團隊和公司,為國內深海裝備的產業化帶來了示范效應。深海原位生物實驗室的研發同樣需要材料、設備制造和配套軟件等相關產業的協同發展,對我國海洋產業走向深海、走向高端和產生高附加值是一個重要機會。
綜上所述,深海原位生物實驗室及其配套技術的發展是科學研究走向深海的必經之路,同時也為我國社會科技和經濟的發展提供了新的契機。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
科技傳播(2019年22期)2020-01-14 03:06:54
