基于轉(zhuǎn)錄組測序技術(shù)研究三角魴和團(tuán)頭魴抗嗜水氣單胞菌感染基因表達(dá)差異
2022-08-27 08:03:44戴瑜來阮松林馮曉宇
漁業(yè)研究 2022年4期
劉 凱,謝 楠,戴瑜來,阮松林,馮曉宇
(杭州市農(nóng)業(yè)科學(xué)研究院,浙江 杭州 310024)
魴屬(Magalobrame)魚類,在分類地位上屬于鯉形目(Cypriniformes)、鯉科(Cyprinidae)和鲌亞科(Cultrinae),該屬主要包括4種魚類:三角魴(Megalobramaterminalis)、團(tuán)頭魴(M.amblycephala)、廣東魴(M.hoffmanni)和厚頜魴(M.pellegrini)等。團(tuán)頭魴,又名“武昌魚”,是我國特有的優(yōu)良草食性魚類,因食性廣、生長快、養(yǎng)殖成本低等優(yōu)點(diǎn),其還是我國主要的大宗淡水水產(chǎn)養(yǎng)殖品種之一[1]。三角魴,在錢塘江附近地區(qū)被稱為“三角鳊”“塔鳊”,在黑龍江附近地區(qū)被稱為“法羅”。相對于團(tuán)頭魴,三角魴的養(yǎng)殖區(qū)域較小,但因具有快速生長期長、商品個體大等特點(diǎn),三角魴在浙江、湖南、山東、江蘇等地也具有一定的養(yǎng)殖規(guī)模,具有良好的養(yǎng)殖前景[2-3]。
近年來,團(tuán)頭魴病害頻發(fā),由嗜水氣單胞菌(Aeromonashydrophila)引起的細(xì)菌性敗血癥是團(tuán)頭魴養(yǎng)殖過程中危害較大的疾病之一,嚴(yán)重影響?zhàn)B殖效益[4]。而養(yǎng)殖實踐發(fā)現(xiàn),與同屬的團(tuán)頭魴相比,三角魴的細(xì)菌性敗血癥發(fā)生率較低,即使發(fā)生了該病,使用常規(guī)藥物就可較好地控制,未有大面積發(fā)病的情況,且病死率低。三角魴和團(tuán)頭魴之間差異小,親緣關(guān)系較近[5],但2種魚類在感染嗜水氣單胞菌后所表現(xiàn)出的差異,除去環(huán)境因素外,遺傳因素是其中一個重要的原因。此前,方獻(xiàn)平等[6]采用非標(biāo)記定量蛋白質(zhì)組學(xué)技術(shù)分析了在嗜水氣單胞菌侵染脅迫后3 h、10 h和24 h肝組織蛋白質(zhì)組的變化,結(jié)果表明三角魴在戊糖磷酸途徑、脂肪酸代謝、亮氨酸降解等通路與團(tuán)頭魴存在顯著差異。為在基因組注釋的豐富度以及期望于基因水平上了解三角魴和團(tuán)頭魴在嗜水氣單胞菌侵染過程中的表達(dá)差異,本文基于轉(zhuǎn)錄組測序技術(shù)研究了三角魴和團(tuán)頭魴在感染嗜水氣單胞菌后轉(zhuǎn)錄組的變化,以期了解三角魴和團(tuán)頭魴在嗜水氣單胞菌感染上的差異性,為闡明嗜水氣單胞菌致病機(jī)制乃至抗病育種提供研究基礎(chǔ)。
1 材料與方法
1.1 實驗材料
實驗用三角魴和團(tuán)頭魴由杭州市農(nóng)業(yè)科學(xué)研究院水產(chǎn)研究所提供,嗜水氣單胞菌株分離自江蘇武進(jìn)某養(yǎng)殖場。致病實驗與方獻(xiàn)平等[6]報道實驗相同,選取了3 h和24 h兩個時間點(diǎn),每個時間點(diǎn)分別取三角魴和團(tuán)頭魴各5尾,體質(zhì)量為130.5~150.5 g。每5尾魚分別取肌肉、肝臟、血液3個組織并分別浸泡于3份RNAfixer無液氮RNA樣品儲存液中,于-80℃冰箱中保存?zhèn)溆谩y序時,每3個組織等質(zhì)量混合后作為1份實驗材料,進(jìn)行建庫和測序。各實驗樣品被分別標(biāo)記為SJF3、SJF20、TTF3、TTF20,表示3 h和24 h兩個時間點(diǎn)采集的三角魴和團(tuán)頭魴樣品。
1.2 RNA提取及建庫測序
RNA提取及建庫測序委托杭州英睿生物科技有限公司完成。取總RNA樣品,利用試劑盒合成cDNA,通過末端修復(fù)、連接接頭和純化,獲得三角魴和團(tuán)頭魴的cDNA文庫,建好的文庫通過Illumina HiseqTM 2000平臺進(jìn)行測序。測序樣本的原始數(shù)據(jù)(Raw Reads)過濾與組裝等步驟參照劉凱等[7]的方法進(jìn)行,過濾數(shù)據(jù)時,采用Cutadapt去除3′端帶接頭的序列以及去除平均質(zhì)量分?jǐn)?shù)低于Q20的Reads,使用Trinity軟件對所有樣本的Clean Reads進(jìn)行拼接,最終組裝獲得Unigenes,作為參考序列。
1.3 功能注釋、分類及代謝途徑分析
通過blastx、Blast2GO和KOBAS等程序?qū)nigenes比對到數(shù)據(jù)庫NR(NCBI non-redundant protein sequences)、COG(Cluster of orthologous groups of proteins)、GO(Gene ontology)和KEGG(Kyoto encyclopedia of genes and genomes)等,進(jìn)而得到Unigenes的功能注釋信息。
1.4 差異基因分析
使用轉(zhuǎn)錄組表達(dá)定量軟件RSEM,以組裝獲得的Unigenes作為參考序列,分別將每個樣品的Clean Reads比對到Unigenes序列上,統(tǒng)計每個樣品比對到每一個基因上的Reads數(shù),并計算每個基因的FPKM值,最終獲得基因表達(dá)譜。分別使用DESeq[8]和DESeq2[9]的R包對比分析三角魴和團(tuán)頭魴在感染嗜水氣單胞菌后的差異基因表達(dá),將P<0.05并且log2|(Fold change)|>1的基因定義為差異基因,分析示意圖見圖1。使用clusterProfiler[10]的R包通過GO和KEGG數(shù)據(jù)庫對主要的差異表達(dá)基因進(jìn)行富集分析。
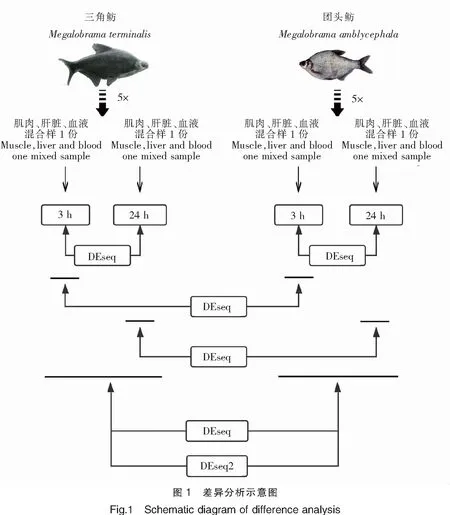
2 結(jié)果與分析
2.1 轉(zhuǎn)錄組數(shù)據(jù)與組裝
三角魴和團(tuán)頭魴的樣本經(jīng)建庫、測序后,每個樣本獲得了超過4 G的數(shù)據(jù)量,共獲得Raw Reads 114 827 718條。去除接頭序列、低質(zhì)量序列后,共獲得Clean Reads 114 824 154條,占總Reads的99.996 8%(表1)。序列經(jīng)拼接和組裝后,共獲得62 780個Unigenes,平均長度達(dá)到531.883 bp,Unigenes的N50為652 bp,組裝完整性較高(表2)。
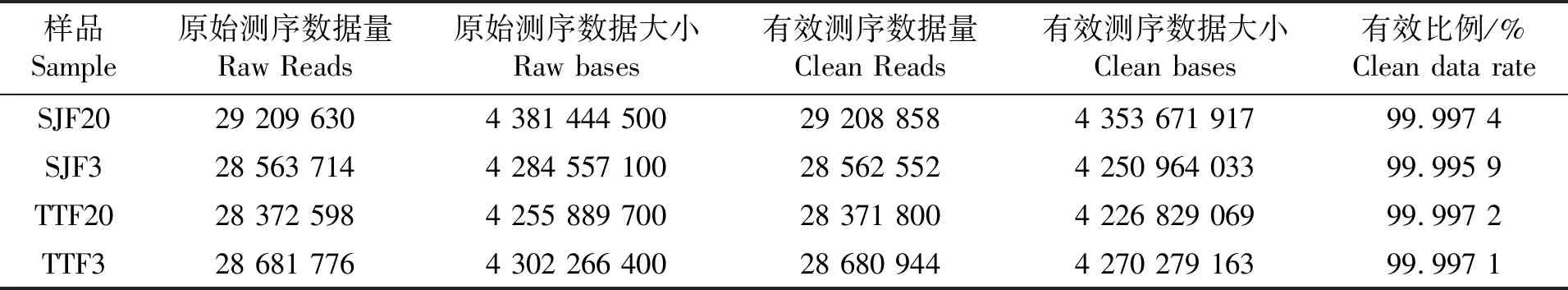
表1 測序數(shù)據(jù)質(zhì)量評估

表2 測序數(shù)據(jù)的統(tǒng)計匯總
2.2 轉(zhuǎn)錄組功能注釋
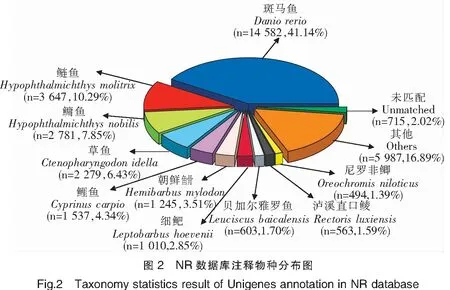
在拼接獲得Unigenes后,與公共數(shù)據(jù)庫進(jìn)行比對注釋,62 780個Unigenes中有35 443個可注釋到NR數(shù)據(jù)庫,比例最高,為56.46%;僅有8 632個可注釋到COG數(shù)據(jù)庫,比例最低,為13.75%(表3)。在NR數(shù)據(jù)庫注釋中,注釋來源最多物種為斑馬魚(Daniorerio),其次為鰱魚(Hypophthalmichthysmolitrix)、鳙魚(Hypophthalmichthysnobilis)等(圖2)。在GO數(shù)據(jù)庫注釋中,11 622個Unigenes被注釋到38個GO Term分類上,其中生物學(xué)過程GO Term分類的相關(guān)基因較多,如GO:0009987 細(xì)胞過程GO Term分類上注釋了4 599個Unigenes;分子功能GO Term分類中,GO:0005488 細(xì)胞結(jié)合GO Term分類相關(guān)基因最多,注釋了7 413個Unigenes。在COG數(shù)據(jù)庫注釋中,一般功能預(yù)測分類注釋的Unigenes最多,為1 979個;其次為翻譯后修飾、蛋白質(zhì)周轉(zhuǎn)、伴侶的分類,注釋Unigenes 1 353個;之后為轉(zhuǎn)錄分類,注釋Unigenes 684個。在KEGG數(shù)據(jù)庫注釋中,注釋Unigenes 最多的前3個通路分別是補(bǔ)體和凝血級聯(lián)、金黃色葡萄球菌感染、吞噬體,注釋Unigenes 達(dá)到9 898、3 721和1 653個。

表3 基因注釋結(jié)果統(tǒng)計
2.3 基因表達(dá)的差異分析
62 780個Unigenes中,有33 693個Unigenes在三角魴中表達(dá),有32 936個Unigenes在團(tuán)頭魴中表達(dá),其中有24 834個Unigenes在三角魴和團(tuán)頭魴中共表達(dá)。基于DESeq,分別比較了三角魴、團(tuán)頭魴在3 h和24 h時間點(diǎn)上基因表達(dá)的差異變化(圖3)。對TTF20-TTF3的差異分析表明,團(tuán)頭魴感染24 h后與感染3 h后相比,顯著上調(diào)表達(dá)的基因有4個,分別為FN1b、PAH、CBLN14和INIH3b.2,顯著下調(diào)表達(dá)基因的前10個主要有ApolipoproteinA-I、C-typelectin14以及UBA、MHCclassⅠantigin和MHCclassⅡalphaantigen等。對SJF20-SJF3的差異分析表明,三角魴感染24 h后與感染3 h后相比,顯著上調(diào)表達(dá)的基因的前10個主要有Ovary-specificC1q-likefactor、ComplementC3以及bain-DAA*0101、Hymo-UA和Meam-UBA等主要組織相容性復(fù)合物(Major histocompatibility complexes,MHC)相關(guān)基因,顯著下調(diào)表達(dá)基因的前10個主要有CBLN6LOC793037、Phosphotransferase、TBL2和Cyca-UA1*01等。對SJF3-TTF3的差異分析表明,在感染3 h后,三角魴相對于團(tuán)頭魴,顯著上調(diào)表達(dá)的基因的前10個主要有Preprohepcidin、FN1b、TBL2、Phosphotransferase和Cyca-UA1*01等,顯著下調(diào)表達(dá)基因的前10個主要有MHCclassⅡalphaantigen、HBB、HBBLOC113082649和Ba1globin等。對SJF20-TTF20的差異分析表明,在感染24 h后,三角魴相對于團(tuán)頭魴,顯著上調(diào)表達(dá)的基因的前10個主要有ApolipoproteinA-I、Ovary-specificC1q-likefactor、Serotransferrin、Hymo-UA和bain-DAA*0101等,顯著下調(diào)表達(dá)基因的前10個主要有CFHL4、BHMTwu:fb53h01、G6PC1a.2和CBLN10RP71-1G18.9等。將以上顯著上調(diào)或下調(diào)表達(dá)的基因進(jìn)行表達(dá)趨勢聚類分析(圖4),三角魴、團(tuán)頭魴在3 h和24 h時間點(diǎn)上基因表達(dá)趨勢可劃分為6類,其中聚類模式4所包含的差異表達(dá)基因最多,為16個;聚類模式1所包含的差異表達(dá)基因數(shù)量為15個;聚類模式6包含基因12個;聚類模式3包含基因10個;聚類模式2、5分別包含基因5個、4個。由此可見,三角魴、團(tuán)頭魴基因表達(dá)趨勢聚類主要模式為4、1、6和3。從以上主要聚類模式可見,三角魴、團(tuán)頭魴之間差異表達(dá)的基因主要以三角魴或團(tuán)頭魴下調(diào)表達(dá)的較多,而上調(diào)表達(dá)的基因較少。
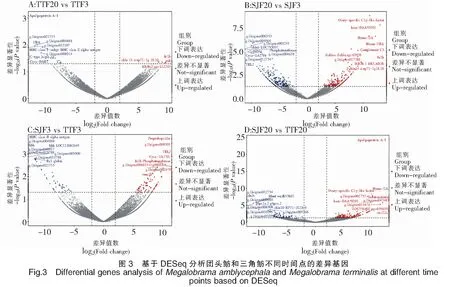
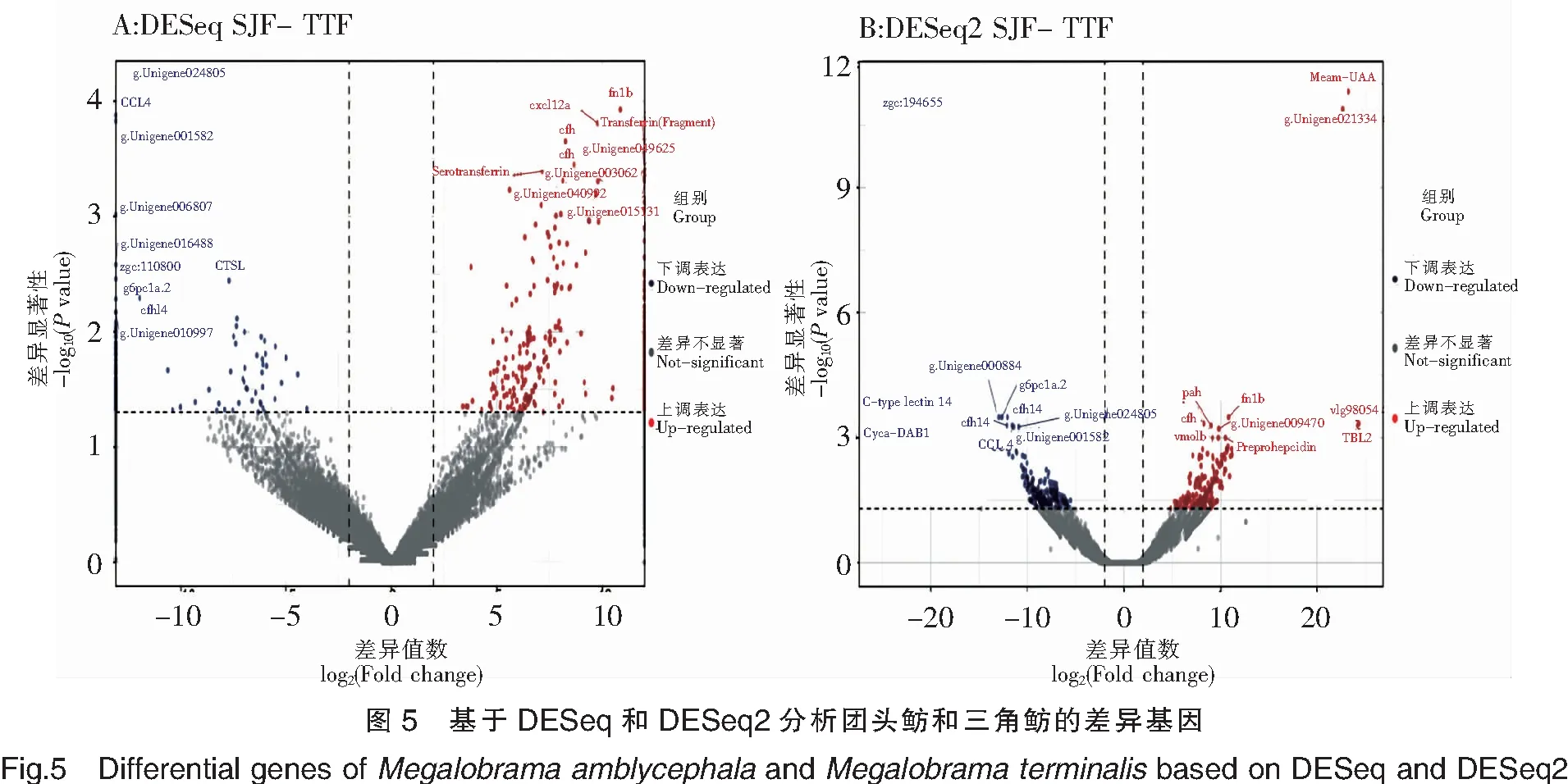
注:紅色點(diǎn)表示顯著上調(diào)的基因,其中顯著上調(diào)的前10個基因標(biāo)注了基因名。藍(lán)色點(diǎn)表示顯著下調(diào)的基因,其中顯著下調(diào)的前10個基因標(biāo)注了基因名。圖5同此。
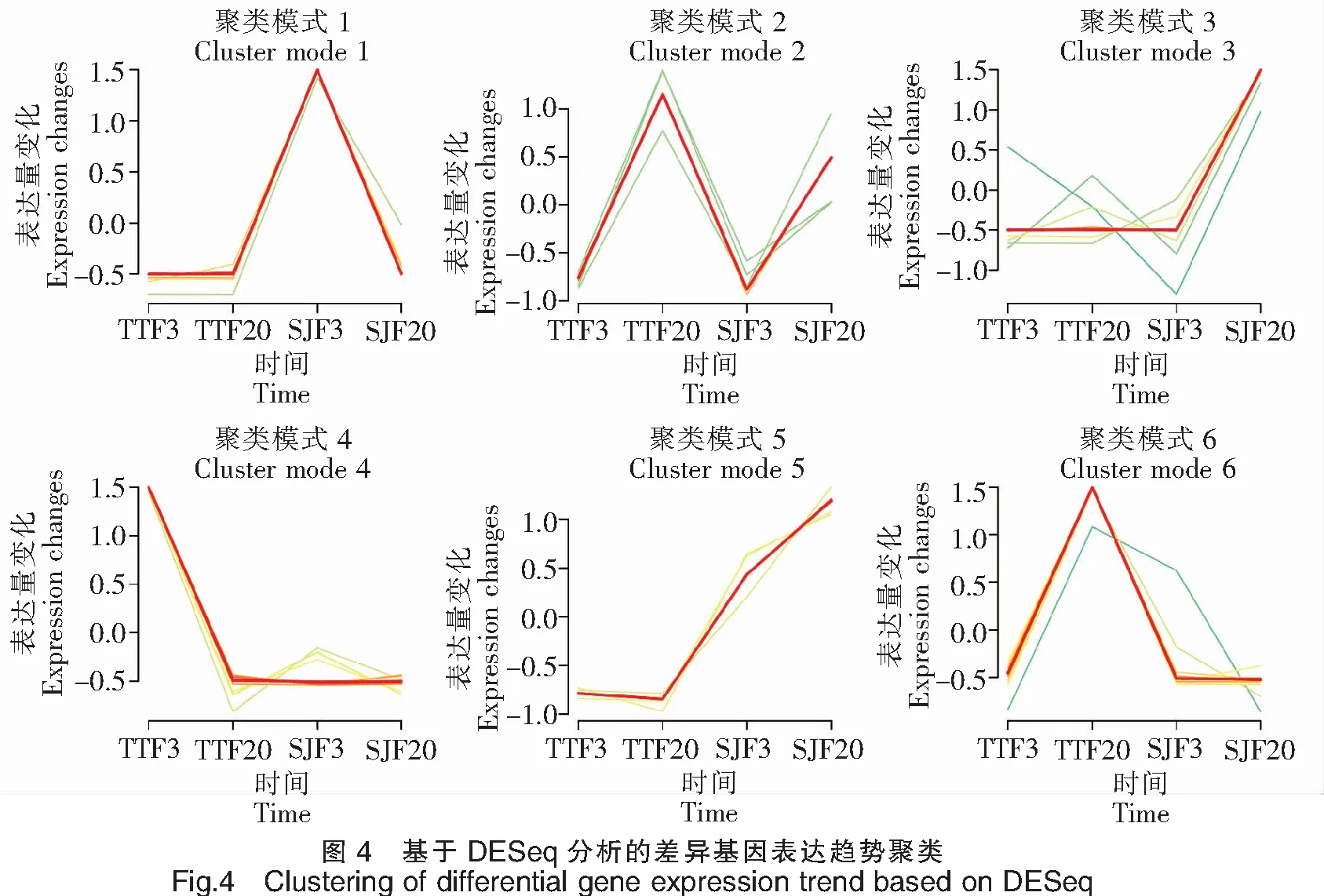
注:不同顏色線表示基于DESeq分析的差異基因在不同時間點(diǎn)和樣本之間的表達(dá)變化。
基于DESeq和DESeq2,從總體上比較了三角魴相對于團(tuán)頭魴基因表達(dá)的差異變化(圖5)。基于DESeq對SJF-TTF的差異分析表明,顯著上調(diào)表達(dá)的基因的前10個主要有FN1b、CXCL12a、CFH、Transferrin(Fragment)和Serotransferrin等,顯著下調(diào)表達(dá)基因的前10個主要有CCL4、CFHL4、G6PC1a.2和CTSL等。基于DESeq2對SJF-TTF的差異分析表明,顯著上調(diào)表達(dá)基因的前10個主要有Meam-UAA、TBL2、CFH、PAH、FN1b和Preprohepcidin等,顯著下調(diào)表達(dá)基因的前10個主要有Cyca-DAB1、CFHL4、C-typelectin14、G6PC1a.2和CCL4等。比較DESeq和DESeq2分析結(jié)果發(fā)現(xiàn),有7個差異表達(dá)的基因在DESeq和DESeq2中均被鑒別出來,分別是上調(diào)表達(dá)基因FN1b和CFH,下調(diào)表達(dá)基因CCL4、CFHL4、G6PC1a.2,以及2個未注釋的基因g.Unigene024805和g.Unigene001582。
2.4 差異表達(dá)基因的富集分析
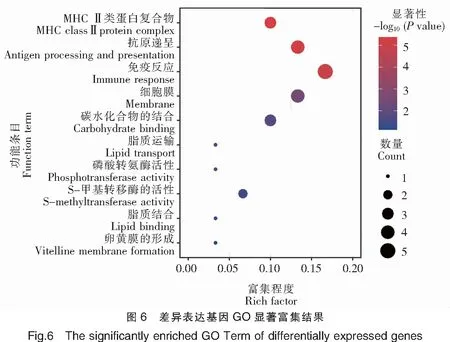
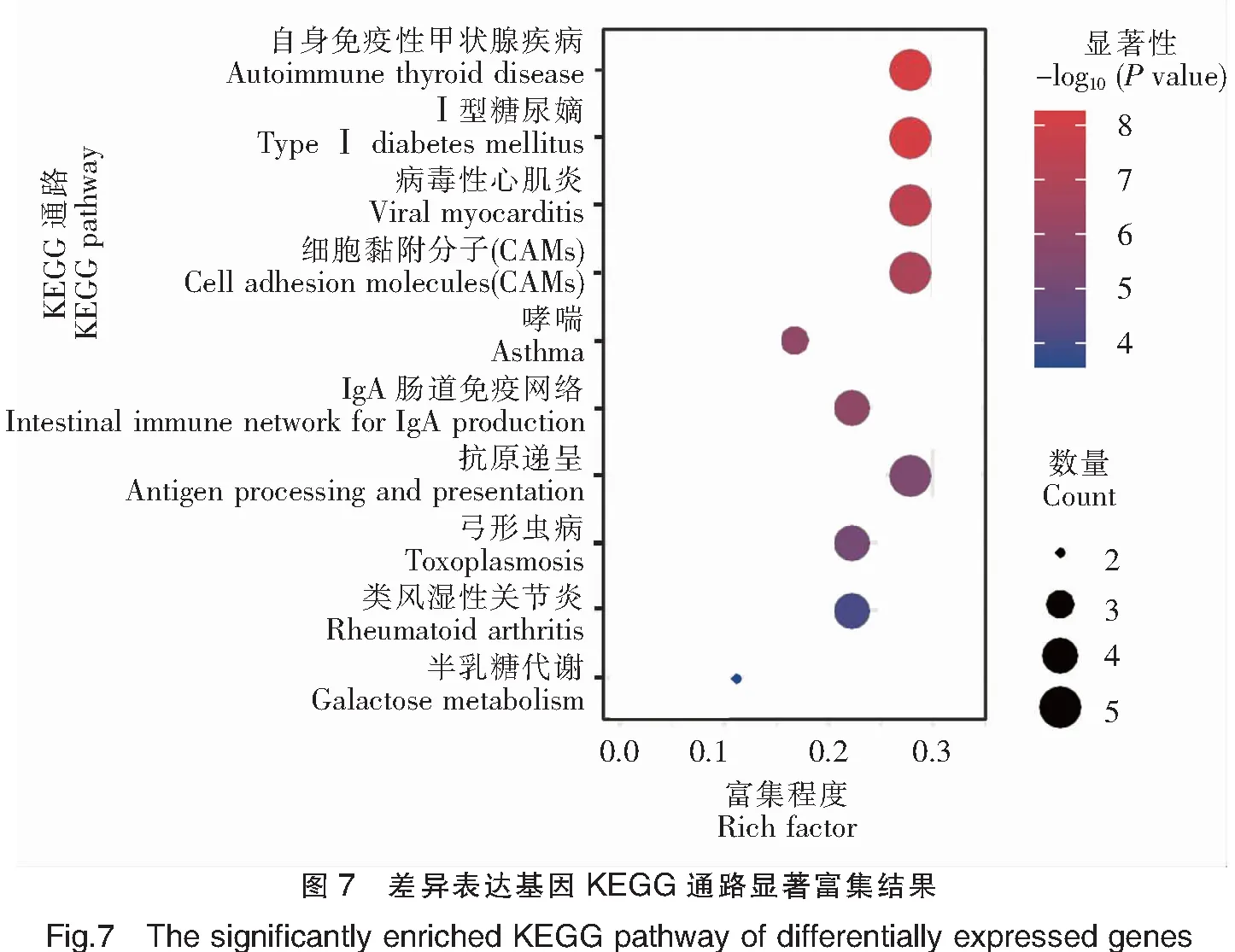
對基于DESeq和DESeq2方法分別獲得的前20個表達(dá)差異最顯著的基因進(jìn)行GO富集分析,通過Rich factor、Pvalue值和富集到GO Term上的基因個數(shù)來衡量富集的程度(圖6)。其中,免疫反應(yīng)(GO:0006955)條目下富集的差異表達(dá)基因最多,30個GO注釋的差異基因中有5個差異基因在此條目下;其次為抗原遞呈(GO:0019882)和細(xì)胞膜(GO:0016020)2個條目,以及MHCⅡ類蛋白復(fù)合物(GO:0042613)和碳水化合物結(jié)合(GO:0030246)等條目。以上結(jié)果表明,三角魴和團(tuán)頭魴之間的抗感染差異與抗原遞呈相關(guān)基因關(guān)系密切。進(jìn)一步對以上獲得的差異表達(dá)基因進(jìn)行KEGG富集分析,富集程度的表征與GO富集分析類似(圖7)。其中,自身免疫性甲狀腺疾病(ko05320)、Ⅰ型糖尿病(ko04940)、病毒性心肌炎(ko05416)、細(xì)胞黏附分子(ko04514)和抗原遞呈(ko04612)5個KEGG通路是富集程度最高的,18個KEGG注釋的差異基因中,均有5個基因富集在這些通路下。剩下的通路還包括IgA腸道免疫網(wǎng)絡(luò)(ko04672)、弓形蟲病(ko05145)和類風(fēng)濕性關(guān)節(jié)炎(ko05323)等。綜上,依然可見抗原遞呈相關(guān)基因與三角魴和團(tuán)頭魴之間的抗感染差異有關(guān),此外細(xì)胞黏附通路(ko04514)也與細(xì)菌感染密切相關(guān)。
3 討論
由于轉(zhuǎn)錄組測序無需事先以已知序列為目標(biāo),因此可以對任意物種進(jìn)行測序而無需知道物種的基因組背景,這為研究物種間基因表達(dá)差異提供了方便[11-12]。因此,本研究基于轉(zhuǎn)錄組測序分析三角魴和團(tuán)頭魴抗嗜水氣單胞菌感染的基因表達(dá)的差異。將三角魴和團(tuán)頭魴樣本的測序數(shù)據(jù)經(jīng)過濾后組裝Unigenes作為參考序列,使用RSEM軟件比對Unigenes序列確定表達(dá)量,以團(tuán)頭魴為對照,分析三角魴Unigenes表達(dá)量的相對變化。此前,方獻(xiàn)平等[6]采用非標(biāo)記定量蛋白質(zhì)組學(xué)技術(shù),分析在嗜水氣單胞菌侵染脅迫后3 h、10 h和24 h時三角魴和團(tuán)頭魴肝組織蛋白質(zhì)組的變化。基于方獻(xiàn)平等[6]的蛋白應(yīng)答聚類分析發(fā)現(xiàn),10 h和24 h的應(yīng)答蛋白質(zhì)優(yōu)選聚類后再與3 h的應(yīng)答蛋白質(zhì)聚類,表明10 h和24 h蛋白質(zhì)應(yīng)答具有相似性。基于此,本研究選擇3 h和24 h作為時間節(jié)點(diǎn)開展轉(zhuǎn)錄組分析。由于方獻(xiàn)平等[6]的分析僅基于肝臟組織,不能完全反映整個機(jī)體對嗜水氣單胞菌侵染的應(yīng)答情況。因此,本研究進(jìn)一步分析了肌肉、肝臟和血液的混合組織樣,以更全面了解三角魴和團(tuán)頭魴在抗嗜水氣單胞菌侵染過程中基因表達(dá)的差異性。此外,由于DESeq2可以消除DESeq的邊界效應(yīng),因此在進(jìn)行整體分析中采用了DESeq2方法與DESeq方法共同進(jìn)行了分析,減少單方法分析的誤差。
基于DESeq的基因表達(dá)趨勢聚類分析表明,三角魴和團(tuán)頭魴在3 h和24 h時間節(jié)點(diǎn)上的基因表達(dá)均表現(xiàn)為表達(dá)量無變化、表達(dá)上調(diào)和表達(dá)下調(diào)等3種情況,但其中主要以三角魴表達(dá)量無變化、團(tuán)頭魴表達(dá)下調(diào)(Cluster 4),團(tuán)頭魴表達(dá)量無變化、三角魴表達(dá)下調(diào)(Cluster 1)為主。可見,三角魴、團(tuán)頭魴之間差異表達(dá)的基因主要以三角魴或團(tuán)頭魴下調(diào)表達(dá)為主,這與羅非魚(Oreochromisniloticus)的情況類似[13],但與蛋白質(zhì)組學(xué)分析結(jié)果存在差異[6]。此外,為從總體上評估三角魴和團(tuán)頭魴感染后基因表達(dá)差異,進(jìn)一步基于DESeq和DESeq2方法進(jìn)行了分析,并進(jìn)一步鑒定出上調(diào)表達(dá)基因FN1b和CFH、下調(diào)表達(dá)基因CFHL4和CCL4等,提示以上基因表達(dá)與三角魴和團(tuán)頭魴的感染差異相關(guān)。FN1b是編碼纖維連接蛋白(Fibronectin,F(xiàn)N)的兩種基因亞型之一[14],纖維連接蛋白則是細(xì)胞外基質(zhì)的重要成分之一,而細(xì)胞外基質(zhì)被認(rèn)為是影響器官形成與修復(fù)的關(guān)鍵因子[15]。CFH(Complement factor H)即補(bǔ)體因子H ,調(diào)節(jié)血漿中補(bǔ)體的替代途徑,抑制補(bǔ)體活性并介導(dǎo)細(xì)胞表面對替代途徑激活劑和非激活劑的區(qū)分[16]。CFH蛋白產(chǎn)生缺陷后可致替代途徑異常激活,導(dǎo)致細(xì)胞損傷[17]。CCL4[Chemokine(C-C motif)ligands 4 ]即CC型趨化因子配體4,是CC型趨化因子家族的一員,因其從脂多糖激活的巨噬細(xì)胞中被分離出來,亦稱MIP-1β[18],它在免疫細(xì)胞遷移、活化、分化和增殖等方面具有重要作用[19]。三角魴相對于團(tuán)頭魴,F(xiàn)N1b和CFH基因表達(dá)顯著上調(diào),表明三角魴在受到細(xì)菌攻擊時更傾向于自我修復(fù),而CCL4基因的下調(diào),則表明團(tuán)頭魴調(diào)動了更多的細(xì)胞因子用于消除細(xì)菌。CFHL4則是補(bǔ)體因子H相關(guān)蛋白(Complement factor H-related protein,F(xiàn)HRs)之一[16],目前關(guān)于其具體機(jī)制并未明確,但越來越多的數(shù)據(jù)表明,F(xiàn)HRs與CFH具有相反的作用,即它們直接增強(qiáng)補(bǔ)體活性[20]。因此,推測三角魴相對于團(tuán)頭魴C(jī)FHL4基因表達(dá)的下調(diào)也與三角魴細(xì)胞自我修復(fù)基因有關(guān)。
基于GO和KEGG富集分析,對DESeq和DESeq2方法鑒定出的主要差異基因進(jìn)行了富集分析,均表明三角魴和團(tuán)頭魴的感染差異與抗原遞呈相關(guān)基因密切相關(guān)。基于轉(zhuǎn)錄組學(xué)對羅非魚感染后的基因表達(dá)分析表明,吞噬作用在羅非魚的抗感染免疫中起到重要作用,其中主要組織相容性復(fù)合物基因就是主要的吞噬相關(guān)基因[13]。本文結(jié)果與此類似,也確定了MHC基因在感染后的重要作用,并表明其與三角魴和團(tuán)頭魴抗感染差異相關(guān)。基于團(tuán)頭魴MHCⅡα基因的研究也表明,MHC基因與團(tuán)頭魴抗病性密切相關(guān)[21]。在DESeq和DESeq2共同鑒定出的差異基因中,僅FN1b被KEGG注釋,其富集在ECM-受體相互作用(ko04512)通路(圖7中未顯示),表明ECM即細(xì)胞外基質(zhì)與三角魴和團(tuán)頭魴抗感染差異密切相關(guān)。此外,在GO和KEGG富集分析中均富集到CXCL12a基因,分別富集于免疫反應(yīng)(GO:0006955)條目和IgA腸道免疫網(wǎng)絡(luò)(ko04672)通路下。CXCL12[Chemokine(C-X-C motif)ligand 12]即CXC型趨化因子配體12,是一種小分子的細(xì)胞因子,與CCL4同屬于趨化因子家族,對淋巴細(xì)胞有強(qiáng)趨化作用[22]。而CXCL12基因在DESeq分析中,三角魴相對于團(tuán)頭魴顯著上調(diào),與團(tuán)頭魴C(jī)CL4基因顯著上調(diào)不同,表明三角魴和團(tuán)頭魴抗感染差異可能與招募淋巴細(xì)胞、促進(jìn)細(xì)胞吞噬的機(jī)制不同相關(guān)。方獻(xiàn)平等[6]基于蛋白質(zhì)組學(xué)技術(shù)發(fā)現(xiàn),相對于團(tuán)頭魴,三角魴β-免疫球蛋白感染后顯著上調(diào)表達(dá)。但在本研究中,盡管檢測到了β-免疫球蛋白基因表達(dá),但在三角魴和團(tuán)頭魴間差異不顯著,這可能是由于蛋白應(yīng)答與基因表達(dá)的不同步性[23],或存在其他原因,都有待進(jìn)一步研究。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機(jī)設(shè)計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
鄭州大學(xué)學(xué)報(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
山東女子學(xué)院學(xué)報(2014年6期)2014-03-01 02:24:55
