紫花苜蓿與不同生活型多年生禾本科牧草混播生長生理特征
2022-08-31 03:33:00張輝輝師尚禮李自立李小龍康文娟陳新棟
草地學報 2022年8期
張輝輝, 師尚禮, 武 蓓, 李自立, 李小龍, 吳 芳, 康文娟, 陳新棟
(甘肅農業大學草業學院, 草業生態系統教育部重點實驗室, 中-美草地畜牧業可持續研究中心, 甘肅 蘭州 730070)
以單一作物耕作生產的傳統農業是一種生產力低、不可持續的生產方式,用糧飼兼顧的草地農業可發揮4倍于土地資源傳統耕作農業利用的潛力[1]。應用優良的豆科牧草與禾本科牧草混播建植人工草地能協同發揮豆科牧草和禾本科牧草的適應性和抗逆性[2],能再次提高自然資源利用率,達到增產[3-6]、提質[7-9]及改土培肥[10-13]的目的。上述混播優勢在生產實踐中能否持續存在,是豆禾混播系統具備優良生產性能的關鍵。在豆禾混播系統中草種組分對水分、養分、光資源和生態位的競爭以及高稈草種對矮桿草種的遮蔭作用會使混播系統組分間產生逆境脅迫,牧草所受逆境脅迫超過其可調節范圍時會限制各組分牧草的生長[14-15]。在混播系統中,植物所受到的逆境脅迫主要來自種間競爭,當受到逆境脅迫時,植物會通過調節自身的抗氧化物酶活性來清除體內積累的活性氧和自由基,減小氧化損傷[16],增加滲透物質含量以減輕或避免丙二醛等有害物質積累造成的組織或器官生物膜系統功能和結構的傷害,通過提高抗氧化能力和滲透調節能力,達到種間生理協同[17],同時,混播系統通過協調植物光合生理來協調種間對光資源的利用,達到提高干物質積累的目的[2]。
楊志超等[15]在紫花苜蓿分別與無芒雀麥、垂穗披堿草(Elymusnutans)、虉草(Phalarisarundinacea)和貓尾草(Phleumpratense)不同比例建植混播草地的研究中發現,混播組合和成分比例顯著影響各組分牧草抗氧化酶活性和抗氧化劑含量。黃麗琴等[18]對苜蓿與虉草混播草地的研究發現,虉草根系可溶性糖含量在不同混播比例下均高于單播,在豆禾間行比為2∶1時最高;苜蓿根系可溶性糖含量在豆禾間行比為1:1時最高,董世魁等[19]發現混播草地光能利用率較單播顯著提高。在豆禾混播系統中,生理調節是牧草響應競爭環境的主要生存策略,目前對混播草地牧草生理響應混播草種和比例的研究較少。本研究以紫花苜蓿分別與3類不同生活型多年生禾本科牧草建植的混播草地為研究對象,通過比較分析不同混播組合和成分比例下牧草抗氧化能力、膜脂過氧化物含量、滲透調節物質含量以及光合生理的差異,探討牧草生長生理對混播系統種間協同效應的響應,為混播草種搭配及比例設置提供參考。
1 材料與方法
1.1 試驗區自然概況
試驗地位于甘肅農業大學蘭州牧草實訓基地,地理位置105°41′ E,34°05′ N,地處黃土高原西端向青藏高原過渡地帶,海拔1 595 m,屬溫帶半干旱大陸性氣候,年平均氣溫10.3℃,年平均日照時數2 374 h,無霜期172 d,年降水量300 mm左右,集中分布于6—9月,年均蒸發量1 664 mm。土壤為黃綿土,肥力均勻,有機質含量為0.96%,堿解氮含量為65.92 mg·kg-1,速效鉀含量為104.21 mg·kg-1,速效磷含量為10.06 mg·kg-1。
1.2 供試材料
供試紫花苜蓿品種為‘清水’(Medicagosativa‘Qingshui’),發芽率86.67%,由甘肅農業大學教育部草業生態系統重點實驗室提供;不同生活型禾本科材料:叢生型-葦狀羊茅,品種為‘雨姿’(Festucaarundinacea‘Yuzi’),發芽率85.56%,匍匐莖型-草地早熟禾,品種為‘蓋世’(Poapratensis‘Gaishi’),發芽率81.11%,由甘肅創綠草業科技有限公司提供;根莖型-無芒雀麥,品種為‘原野’(Bromusinermis‘Yuanye’),發芽率82.22%,由北京正道草業有限公司提供。
1.3 試驗設計
試驗設3個混播組合,每個混播組合設3個混播比例(混播播種量按占單播量的比例計算),另設置4個單播處理作為對照(表1),共13個處理,小區面積15 m2(5 m×3 m),重復3次,總共39個小區,隨機區組排列,小區間距0.4 m,行距0.2 m,采用同行條播的播種方式,播種深度2 cm。播種時間2020年5月25日,底肥施用量:磷酸二銨(P2O5≥46%)120 kg·hm-2;尿素(總氮≥46%)80 kg·hm-2;硫酸鉀(K2O≥51%)50 kg·hm-2,底肥播種前均勻撒布于地表,翻耕入土,整地種植。試驗地采用滴灌方式進行灌溉,生長期間不追肥,采用人工除草。2020年5月25日播種,2020年8月18日和2021年6月2日紫花苜蓿初花期取樣并測定相關指標。
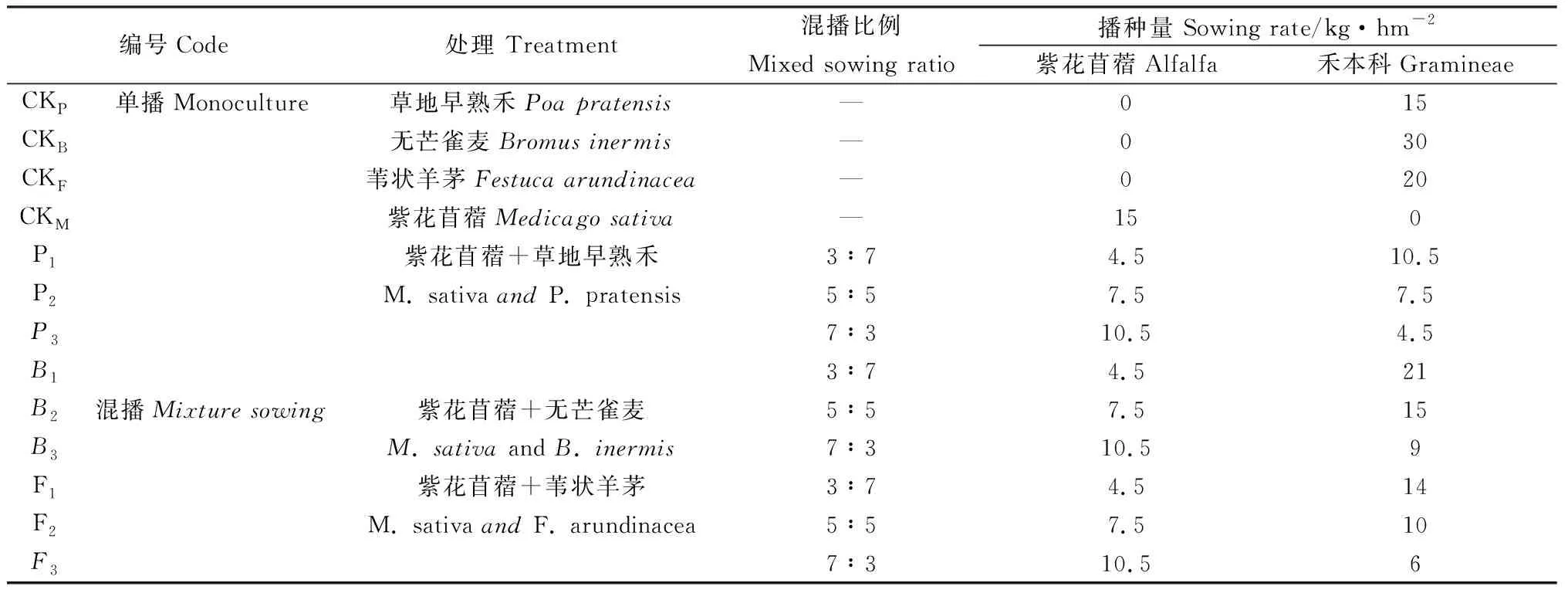
表1 試驗處理Table 1 Experimental treatments
1.4 測定指標與方法
單播區每小區取3個1 m樣段,稱鮮重,然后每個樣段取500 g鮮樣;混播區每小區取3個1 m樣段,取樣后豆禾分開稱鮮重,每個樣段豆禾各取鮮樣500 g;鮮樣帶回實驗室后于105℃下殺青20 min,80℃下烘干至恒重,2年數據取平均值,通過測產面積和草樣干鮮比換算獲得混播草地產量,并根據產量計算各組分增產貢獻率:
式中:YIR表示混播系統增產率,Yij和Yji分別表示混播系統中豆科牧草和禾本科牧草的產量,Yi和Yj分別表示豆科牧草和禾本科牧草的單播產量,Pi和Pj分別表示豆科牧草和禾本科牧草的混播比例,且Pi+Pj=1。

株高:用卷尺測量任意一株牧草從地面至葉尖或花序頂端的絕對高度,10株為一個重復,取其平均值,共3次重復。
紫花苜蓿初花期時于晴天9:30—11:00采用GFS-3000便攜式光合儀(Walz,Germany)對紫花苜蓿功能葉片倒4葉的光合氣體交換參數進行測定,空氣中CO2濃度固定為400 μmol·mol-1,光照強度設定為1400 μmol·(m2·s)-1;對草地早熟禾頂第2片完全展開的功能葉的氣體交換參數進行測定,空氣中CO2濃度固定為400 μmol·mol-1,光照強度設定為1 000 μmol·(m2·s)-1;對無芒雀麥頂端第2片完全展開的功能葉的氣體交換參數進行測定,空氣中CO2濃度固定為400 μmol·mol-1,光照強度設定為2 000 μmol·m-2·s-1;對葦狀羊茅頂端第2片完全展開的功能葉的氣體交換參數進行測定,空氣中CO2濃度固定為400 μmol·mol-1,光照強度設定為800 μmol·m-2·s-1;測定指標包括:凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)和胞間CO2濃度(Ci)。
于紫花苜蓿初花期隨機選取單株,剪取從頂部向下第3~5片完整葉,每個小區10個重復單株。將取好的葉片裝入錫箔紙袋在液氮中迅速冷凍,帶回實驗室后于-80℃超低溫冰箱儲藏[20],用于測定指標:超氧化物歧化酶(SOD)活性采用氮藍四唑(NBT)光還原法測定[21],過氧化物酶(POD)活性采用愈創木酚法測定[22],過氧化氫酶(CAT)活性采用試劑盒法(試劑盒由蘇州格銳思生物公司提供),丙二醛(MDA)含量采用硫代巴比妥酸法測定[23],脯氨酸(Pro)含量采用酸性茚三酮法測定[24],可溶性糖(SS)含量采用蒽酮法測定[25],可溶性蛋白含量(SP)采用考馬斯亮藍染色法[26],并用牛血清白蛋白作標準曲線。
1.5 數據分析
采用Excel2010進行數據統計、計算及隸屬函數分析,SPSS(IMB SPSS Statiatics 19)進行數據單因素方差分析,并用Duncan法對各測定數據進行多重比較。
利用下列公式計算不同混播處理標準化數據綜合指標的隸屬函數值,與植物生長生理協同能力呈正相關的參數,如SOD,POD,CAT,Pro,SS和SP等采用公式:
u(Xj)=(Xj-Xmin)/(Xmax-Xmin),
j=1,2,3,…,n
與植物生長生理協同能力呈負相關的參數如MDA采用公式:
u(Xj)=1-(Xj-Xmin)/(Xmax-Xmin),
j=1,2,3,…,n
式中,Xj表示第j個綜合指標,u(Xj)表示j個綜合指標的隸屬函數值,Xmax和Xmin分別表示第j個綜合指標的最大值和最小值。
權重采用客觀賦權法進行計算:
式中,Wj表示第j個綜合指標的權重,Pj表示第j個綜合指標的貢獻率。
綜合評價D值計算公式:
2 結果與分析
2.1 混播成分與比例對牧草產量的影響
不同混播處理下牧草產量如表2所示。混播草地產量均較單播草地顯著提高(P<0.05),隨紫花苜蓿混播比例的下降和禾草混播比例的上升,紫花苜蓿產量下降,禾草產量上升,混播草地總產量下降。不同處理的混播草地各組分增產率和混播系統增產率均為正值,較單播草地增產。紫花苜蓿與草地早熟禾P1處理下紫花苜蓿增產率最大,P2處理下草地早熟禾增產率最大;紫花苜蓿與無芒雀麥B2處理下紫花苜蓿增產率最大,B3處理下無芒雀麥增產率最大;紫花苜蓿與葦狀羊茅P1處理下紫花苜蓿與葦狀羊茅增產率均最大。
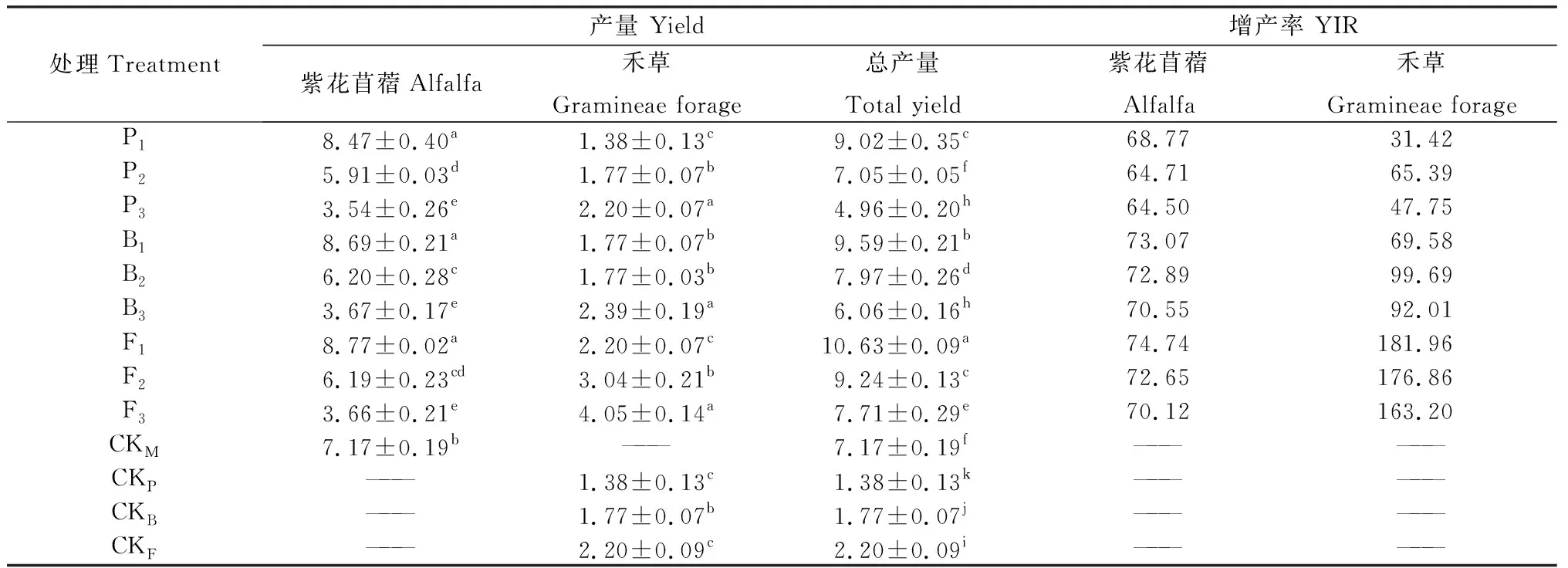
表2 不同混播處理下牧草產量的差異Table 2 Effect of different mixed sowing treatment on yield of forage
2.2 混播成分與比例對牧草株高的影響
不同混播處理下牧草株高如圖1所示,在紫花苜蓿與草地早熟禾混播草地中,混播紫花苜蓿株高2020年和2021年分別為75.94~77.51 cm和86.44~101.64 cm,在兩年試驗各混播處理下均顯著低于單播(P<0.05);混播草地早熟禾株高2020年和2021年分別為40.33~44.78 cm和49.83~58.13 cm,各年份混播處理均高于單播,其中在2020年各混播處理及2021年P1處理下顯著高于單播(P<0.05)。在紫花苜蓿與無芒雀麥混播草地中,混播紫花苜蓿株高2020年和2021年分別為68.07~72.17 cm和85.94~99.04 cm,在兩年各混播處理下均顯著低于單播(P<0.05);混播無芒雀麥株高2020年和2021年分別為44.17~47.83 cm和49.83~58.13 cm,在2020年B1和B2處理以及2021年各處理下顯著高于單播(P<0.05)。在紫花苜蓿與葦狀羊茅混播草地中,混播紫花苜蓿株高2020年和2021年分別為74.22~77.75 cm和85.42~96.99 cm,在兩年F2和F3處理下均顯著低于單播(P<0.05);混播葦狀羊茅株高2020年和2021年分別為68.52~72.63 cm和82.70~92.40 cm,在兩年各處理下顯著高于單播(P<0.05)。
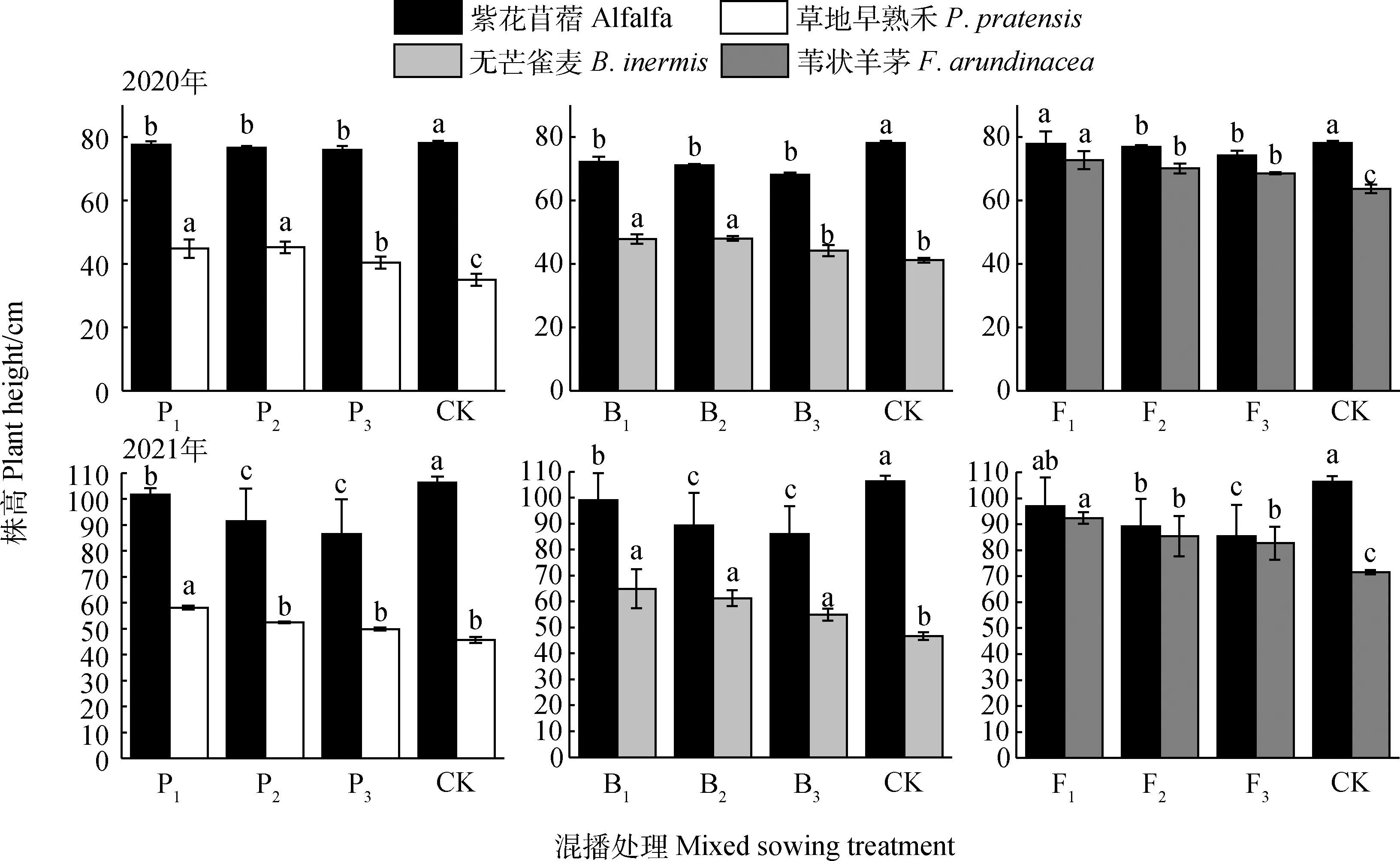
圖1 不同混播處理對牧草株高的影響Fig.1 Effects of different mixed sowing treatment on plant height of forage
2.3 混播成分與比例對牧草葉片膜脂過氧化物含量的影響
如圖2所示,不同混播處理下牧草葉片MDA含量具有差異。在紫花苜蓿與草地早熟禾混播草地中,紫花苜蓿葉片MDA含量在兩年試驗各混播處理下均顯著低于單播(P<0.05);草地早熟禾葉片MDA含量在兩年各混播處理下均顯著高于單播(P<0.05)。在紫花苜蓿與無芒雀麥混播草地中,紫花苜蓿葉片MDA含量在兩年試驗P2和P3處理下均顯著低于單播(P<0.05);無芒雀麥葉片MDA含量在2020年B2和B3處理下顯著高于單播(P<0.05),在2021年各處理與單播差異不顯著。在紫花苜蓿與葦狀羊茅混播草地中,紫花苜蓿葉片MDA含量在2020年各處理以及2021年F2和F3處理下顯著低于單播(P<0.05);葦狀羊茅葉片MDA含量在2020年F2和F3處理以及2021年各處理下顯著低于單播(P<0.05)。隨紫花苜蓿比例的下降和禾草比例的上升,紫花苜蓿與3種禾草葉片MDA含量均呈下降趨勢。
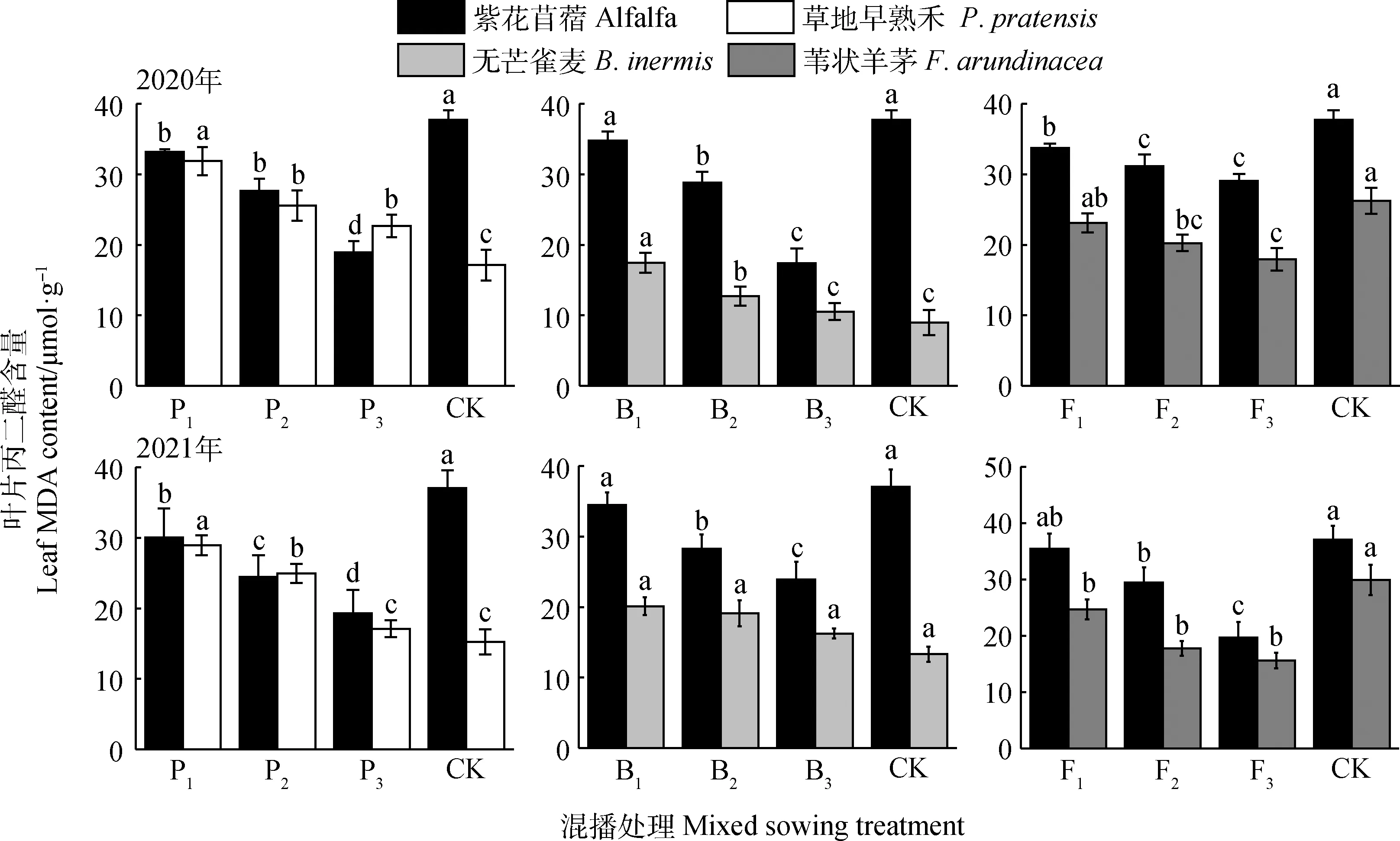
圖2 不同混播處理對葉片丙二醛含量的影響Fig.2 Effects of different mixed sowing treatment on malondialdehyde content of leaf
2.4 混播成分與比例對牧草葉片抗氧化酶活性的影響
不同混播處理下牧草葉片SOD酶活性如圖3所示。在紫花苜蓿與草地早熟禾混播草地中,紫花苜蓿葉片SOD酶活性在2020年P1和P2處理以及2021年各處理下均顯著高于單播(P<0.05);草地早熟禾葉片SOD酶活性在2020年P2和P3處理以及2021年P1處理下顯著高于單播(P<0.05)。在紫花苜蓿與無芒雀麥混播草地中,紫花苜蓿葉片SOD酶活性在2020年各處理及2021年P1和P2處理下均顯著高于單播(P<0.05);無芒雀麥葉片SOD酶活性在2020年B2和B3處理以及2021年各處理下顯著高于單播(P<0.05)。在紫花苜蓿與葦狀羊茅混播草地中,紫花苜蓿葉片SOD酶活性在2020年各處理以及2021年F1和F2處理下顯著高于單播(P<0.05);葦狀羊茅葉片SOD酶活性在兩年F1和F2處理下顯著高于單播(P<0.05)。
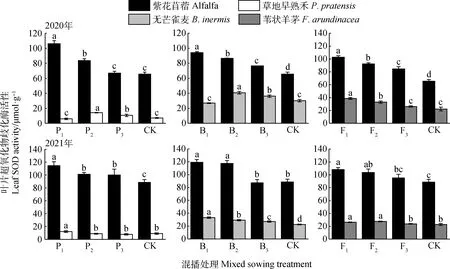
圖3 不同混播處理對葉片超氧化物歧化酶活性的影響Fig.3 Effects of different mixed sowing treatment on SOD activity of leaf
如圖4所示,在紫花苜蓿與草地早熟禾混播草地中,紫花苜蓿葉片POD酶活性在2020年P1和P2處理以及2021年各處理下均顯著高于單播(P<0.05);草地早熟禾葉片POD酶活性在2020年P2和P3處理以及2021年P1處理下顯著高于單播(P<0.05)。在紫花苜蓿與無芒雀麥混播草地中,紫花苜蓿葉片POD酶活性在2020年各處理及2021年P1和P2處理下均顯著高于單播(P<0.05);無芒雀麥葉片POD酶活性在2020年B2和B3處理以及2021年各處理下顯著高于單播(P<0.05)。在紫花苜蓿與葦狀羊茅混播草地中,紫花苜蓿葉片POD酶活性在2020年各處理以及2021年F1和F2處理下顯著高于單播(P<0.05);葦狀羊茅葉片POD酶活性在兩年F1和F2處理下顯著高于單播(P<0.05)。
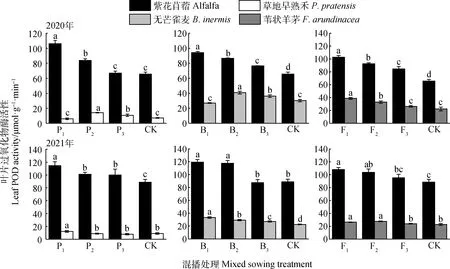
圖4 不同混播處理對葉片過氧化物酶活性的影響Fig.4 Effects of different mixed sowing treatment on POD activity of leaf
如圖5所示,在紫花苜蓿與草地早熟禾混播草地中,紫花苜蓿葉片CAT酶活性在2020年各處理以及2021年P1和P2處理下均顯著高于單播(P<0.05);草地早熟禾葉片CAT酶活性在2020年各處理以及2021年P1和P2處理下顯著高于單播(P<0.05)。在紫花苜蓿與無芒雀麥混播草地中,紫花苜蓿葉片CAT酶活性在2020年B1和B2處理及2021年各處理下均顯著高于單播(P<0.05);無芒雀麥葉片CAT酶活性在2020年B2處理以及2021年各處理下顯著高于單播(P<0.05)。在紫花苜蓿與葦狀羊茅混播草地中,紫花苜蓿葉片CAT酶活性在2020年F1處理以及2021年各處理下顯著高于單播(P<0.05);葦狀羊茅葉片CAT酶活性在2020年F1和F2處理以及2021年各處理下顯著高于單播(P<0.05)。
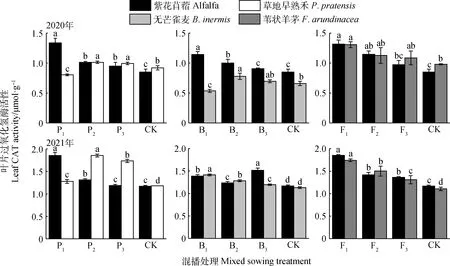
圖5 不同混播處理下葉片過氧化氫酶活性的差異Fig.5 Effects of different mixed sowing treatment on CAT activity of leaf
總體而言,混播處理下多組分刺激了SOD酶活性、POD酶活性及CAT酶活性的變化,改變了植物清除體內活性氧的能力,以響應組分間競爭。
2.5 混播成分與比例對牧草葉片滲透調節物質含量的影響
如圖6所示,不同混播處理下牧草葉片Pro含量具有差異。在紫花苜蓿與草地早熟禾混播草地中,紫花苜蓿葉片Pro含量在2020年各處理以及2021年P1和P2處理下均顯著高于單播(P<0.05);草地早熟禾葉片Pro含量在2020年P1和P2處理以及2021年各處理下顯著高于單播(P<0.05)。在紫花苜蓿與無芒雀麥混播草地中,紫花苜蓿葉片Pro含量在2020年各處理及2021年B1和B2處理下均顯著高于單播(P<0.05);無芒雀麥葉片Pro含量在兩年各處理下顯著高于單播(P<0.05)。在紫花苜蓿與葦狀羊茅混播草地中,紫花苜蓿葉片Pro含量在2020年F1和F2處理以及2021年各處理下顯著高于單播(P<0.05);葦狀羊茅葉片Pro含量在2020年F1處理以及2021年F1和F2處理下顯著高于單播(P<0.05)。
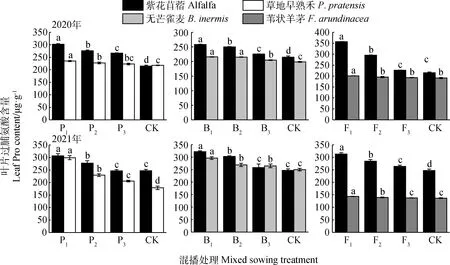
圖6 不同混播比例處理下葉片脯氨酸含量的差異Fig.6 Effects of different mixed sowing ratio treatment on Pro content of leaf
如圖7所示,在紫花苜蓿與草地早熟禾混播草地中,紫花苜蓿葉片SS含量在2020年P1和P2處理以及2021年P1處理下均顯著高于單播(P<0.05);草地早熟禾葉片SS含量在2020年P2處理以及2021年P1處理下顯著高于單播(P<0.05)。在紫花苜蓿與無芒雀麥混播草地中,紫花苜蓿葉片SS含量在兩年B1和B2處理下均顯著高于單播(P<0.05);無芒雀麥葉片SS含量在兩年B1處理下顯著高于單播(P<0.05)。在紫花苜蓿與葦狀羊茅混播草地中,紫花苜蓿葉片SS含量在2020年F1處理以及2021年F1和F2處理下顯著高于單播(P<0.05);葦狀羊茅葉片SS含量在2021年F1處理下顯著高于單播(P<0.05)。
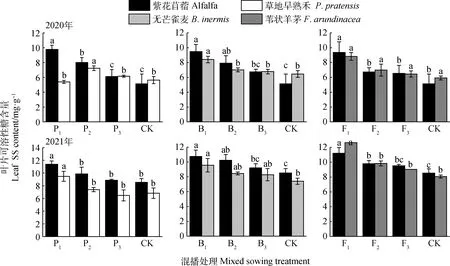
圖7 不同混播比例處理下葉片可溶性糖含量的差異Fig.7 Effects of different mixed sowing ratio treatment on SS content of leaf
如圖8所示,在紫花苜蓿與草地早熟禾混播草地中,紫花苜蓿葉片SP含量在2020年P1和P2處理以及2021年各處理下均顯著高于單播(P<0.05);草地早熟禾葉片SP含量在2020年P2處理以及2021年P1和P2處理下顯著高于單播(P<0.05)。在紫花苜蓿與無芒雀麥混播草地中,紫花苜蓿葉片SP含量在兩年各處理下均顯著高于單播(P<0.05);無芒雀麥葉片SP含量在2021年B1處理以及2021年B1和B2下顯著高于單播(P<0.05)。在紫花苜蓿與葦狀羊茅混播草地中,紫花苜蓿葉片SP含量在兩年各處理下顯著高于單播(P<0.05);葦狀羊茅葉片SP含量在2021年F1處理下顯著高于單播(P<0.05)。
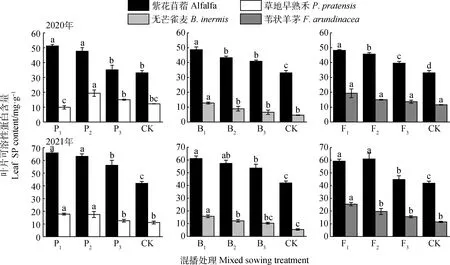
圖8 不同混播比例處理下葉片可溶性蛋白含量的差異Fig.8 Effects of different mixed sowing ratio treatment on SP content of leaf
總體而言,混播較單播相比,多組分刺激了Pro含量、SS含量及SP含量的變異,調節了細胞滲透勢平衡,以響應組分間激烈的競爭。
2.6 混播成分與比例對牧草光合生理的影響
不同混播處理下牧草光合生理變化如表3所示,不同混播處理下牧草光合生理具有差異。各混播處理下紫花苜蓿凈光合速率、蒸騰速率、氣孔導度和胞間CO2濃度均顯著大于單播處理(P<0.05)。草地早熟禾凈光合速率在兩年P1混播處理下均顯著小于單播(P<0.05),在2020年P2和P3混播處理以及2021年P2處理下顯著大于單播(P<0.05);草地早熟禾蒸騰速率在2020年P1混播處理下顯著小于單播(P<0.05),在兩年P2和P3混播處理下均顯著大于單播(P<0.05);草地早熟禾氣孔導度在2020年P2和P3混播處理以及2021年P2混播處理下顯著大于單播(P<0.05),在2021年P1混播處理下顯著小于單播(P<0.05);草地早熟禾胞間CO2濃度在兩年P2和P3混播處理下均顯著大于單播(P<0.05),在P1混播處理下與單播差異不顯著。無芒雀麥凈光合速率在2020年B2和B3混播處理以及2021年各混播處理下均顯著大于單播(P<0.05);無芒雀麥蒸騰速率在兩年各混播處理下顯著大于單播(P<0.05);無芒雀麥氣孔導度在2020年B2和B3混播處理以及2021年各混播處理下顯著大于單播(P<0.05);無芒雀麥胞間CO2濃度在2020年各混播處理以及2021年B2處理下顯著大于單播(P<0.05)。葦狀羊茅凈光合速率在2020年F1和F2混播處理以及2021年各混播處理下均顯著大于單播(P<0.05);葦狀羊茅蒸騰速率在兩年各混播處理下顯著大于單播(P<0.05);葦狀羊茅氣孔導度在2020年F1和F2混播處理以及2021年各混播處理下顯著大于單播(P<0.05);葦狀羊茅胞間CO2濃度在2020年F1混播處理以及2021年各處理下顯著大于單播(P<0.05)。
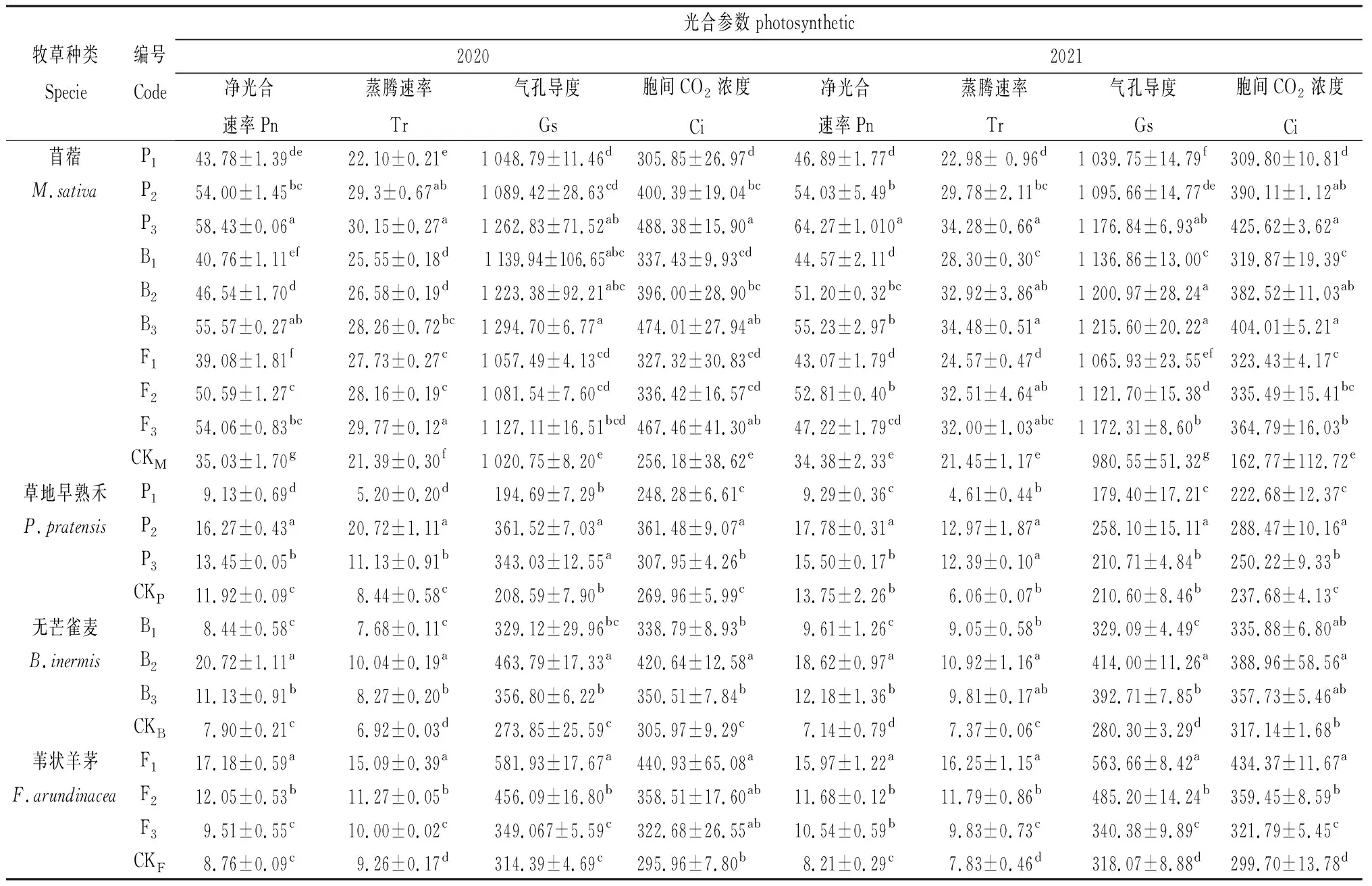
表3 不同混播處理下牧草光合指標的差異Table 3 Effects of different mixed sowing treatment on photosynthetic index of forage
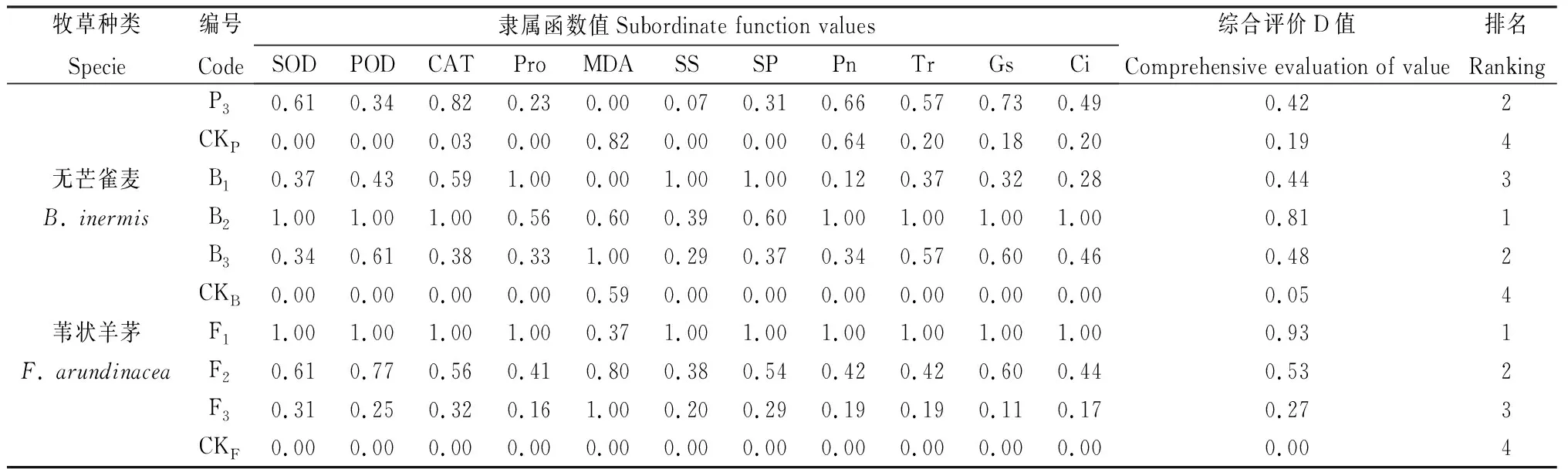
2.7 紫花苜蓿與3種生活型禾草混播牧草生長生理指標綜合評價
混播草地牧草通過調節多種生理物質來提高生理協同效應,單一的生理指標難以準確反映牧草的生理協同能力,本研究采用隸屬函數法對紫花苜蓿與禾草的生長生理指標進行綜合評價,綜合評價D值越大,說明牧草生長生理表現越好,結果如表4所示。根據綜合評價D值對各混播及單播處理牧草生理排序,結果顯示混播紫花苜蓿綜合評價D值均大于單播,綜合評價D值由大到小的排列為F1>B2>P1=B1>F2>P2>B3>P3=F3>CKM,混播草地早熟禾綜合評價D值大于單播,綜合評價D值由大到小排列為P2>P3>P1>CKP,混播無芒雀麥綜合評價D值在混播處理下均大于單播,綜合評價D值由大到小排列為B2>B3>B1>CKB,葦狀羊茅綜合評價D值在混播處理下均大于單播,綜合評價D值由大到小排列為F1>F2>F3>CKF。
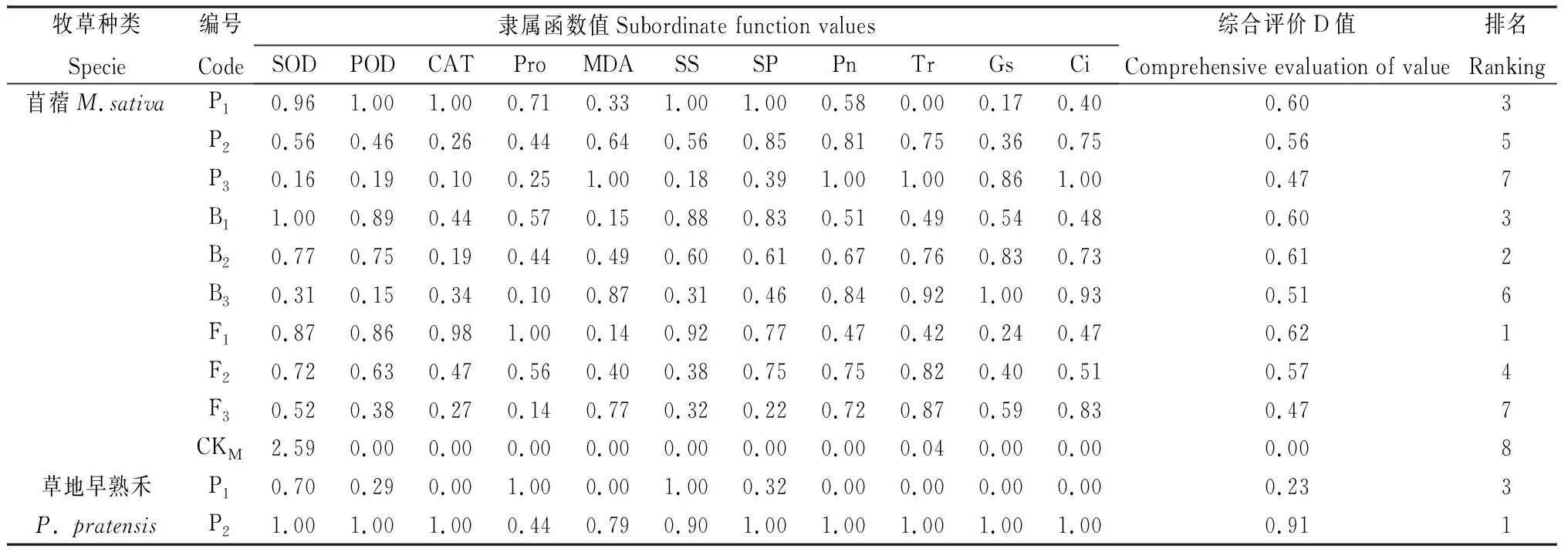
表4 不同混播處理下牧草各生長生理指標隸屬函數值及綜合評價Table 4 Subordinate function values and comprehensive evaluation values of various mixed pasture treatments
續表4
3 討論
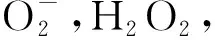
MDA是植物體內自由基作用于膜脂而發生過氧化反應的主要產物,是加劇生物膜損傷的主要物質,其含量的高低可以間接反映膜脂過氧化程度[32]。已有研究表明逆境脅迫增加了植物體內MDA的含量,其含量與脅迫程度有關[33]。本研究發現,在兩年試驗中,紫花苜蓿與3種多年生禾草混播時紫花苜蓿葉片MDA含量均較單播降低,草地早熟禾和無芒雀麥葉片MDA含量均較單播增加,葦狀羊茅葉片MDA含量均較單播降低,根據前人研究[34]可以看出混播紫花苜蓿和葦狀羊茅所受逆境脅迫小于單播,混播草地早熟禾和無芒雀麥受到逆境脅迫大于單播。同時,本研究發現隨著紫花苜蓿混播比例的減小和禾草混播比例增大,紫花苜蓿的MDA含量呈下降的趨勢,推斷混播紫花苜蓿種內對自然資源和生態位的競爭大于種間;草地早熟禾和無芒雀麥的MDA含量隨這種混播比例的變化呈下降的趨勢,推斷在混播草地中株高較高的紫花苜蓿對光資源的競爭力大于地早熟禾和無芒雀麥,草地早熟禾和無芒雀麥在光資源競爭中處于劣勢,同時矮稈、下繁草的草地早熟禾和無芒雀麥受到高稈、上繁草紫花苜蓿的遮蔭作用,造成一定的逆境脅迫[34],隨紫花苜蓿混播比例的減小,紫花苜蓿的競爭力減小,遮蔭作用減小,草地早熟禾所受逆境脅迫減小,MDA含量下降,符合陳意蘭等[35]研究發現的MDA含量隨脅迫程度的增加而增加的結果,與付娟娟等[36]發現的遮蔭程度越強,禾草所受逆境脅迫越大的結果一致。混播葦狀羊茅葉片MDA含量隨這種比例變化呈下降的趨勢但始終低于單播處理,可能是因為紫花苜蓿與草地早熟禾株高差異較小,葦狀羊茅不受紫花苜蓿遮蔭競爭造成的壓迫,兩草種種間對資源的協同性良好,與秦燕[37]研究發現的燕麥與箭筈豌豆在2∶1混作下兩種牧草對資源協同性最好的研究結果相似,但因草種不同而協同比例存在差異。混播草地牧草受到的逆境脅迫來源于多方面,本研究只對牧草是否受到逆境脅迫進行了分析,對于造成逆境脅迫的具體原因需要日后進一步探討。
雖然ROS的累積能引起膜脂過氧化,進而破壞生物膜及細胞正常的生理過程,但生物體內存在抗氧化系統,主要由SOD,POD和CAT組成,能有效清除ROS[38],使得混播牧草種間生長生理達到正常水平。SOD是抗氧化系統清除ROS的核心酶,主要功能是催化超氧化物的歧化反應,將OH-和O2-轉化形成H2O2,再由POD和CAT進一步將其轉化為H2O[39-40]。吳明浩[41]以苜蓿分別與3種禾草以不同比例混播后發現,當豆禾比為1∶2時,禾草葉片中SOD和CAT活性顯著高于其他處理,丙二醛含量低于其他處理,種間競爭脅迫較小。本研究發現紫花苜蓿與3種禾草混播時紫花苜蓿葉片SOD,POD和CAT酶活性大于單播,說明與禾草混播能有效提高紫花苜蓿的抗氧化能力,適當的MDA能上調紫花苜蓿抗氧化能力,草地早熟禾和無芒雀麥的SOD,POD和CAT酶活性在2020年5∶5和3∶7豆禾比下高于單播,在7∶3豆禾比下低于單播,2021年各混播處理下均大于單播,說明適當比例的紫花苜蓿增大了草地早熟禾和無芒雀麥的抗氧化能力,但建植初期過大比例的紫花苜蓿對草地早熟禾和無芒雀麥的競爭脅迫造成了兩種禾草抗氧化系統損傷,建植第二年逐漸適應了競爭環境。兩年試驗中混播葦狀羊茅的SOD,POD和CAT酶活性均大于單播,說明與紫花苜蓿混播增大了無芒雀麥抗氧化能力。本研究中,3種抗氧化酶活性的變化趨勢基本一致,且與MDA含量的變化基本一致,符合蔣明義[42]提出的SOD,POD和CAT三種酶協同一致維持自由基平衡的說法,與張春平[43]發現的植物所受脅迫程度影響抗氧化物質含量結果一致。
另外,植物在逆境條件下會迅速積累某些小分子化學物質如Pro,SS和SP等來維持細胞的滲透勢,維持體內水分平衡和細胞膨壓,從而提高混播草地牧草種間協同性。Pro是一種具有重要滲透調節作用的電解質,在逆境條件下植物會升高Pro含量來維持細胞基質與環境之間的滲透平衡和減少水分散失,其含量可從一定程度上反映植物抗逆性的強弱[44]。可溶性糖是保障植物體內物質正常運輸的主要物質[45],其也參與滲透調節過程,同時還是重要的能量來源。可溶性蛋白參與植物細胞的滲透調節,可以增強植物耐脫水能力,抗性越強,植物體內可溶性蛋白越高[46]。本研究發現混播草地牧草Pro含量,SS含量和SP含量的變化趨勢與MDA基本一致,與陳意蘭[35]在蒭雷草(ThuareaPers)對鹽脅迫的生理響應的研究中發現的蒭雷草SP和Pro含量隨脅迫程度增大而增大的結果相似,與賈新平[47]發現的植物通過提高滲透調節物質含量以響應逆境脅迫的研究結果一致,說明混播植物可在逆境條件下可通過提高滲透調節物質含量來維持滲透壓平衡,但建植第一年7∶3豆禾比下草地早熟禾滲透調節物質含量低于單播,說明過大的紫花苜蓿比例造成的逆境脅迫使草地早熟禾過氧化物質清除能力下降,MDA積累,造成膜脂過氧化損傷,建植第二年逐漸適應了競爭環境,為造成膜脂過氧化損傷。
在水肥相對充足的條件下,光資源是植物種間競爭的主要因素,高稈作物對矮稈作物會造成遮蔭脅迫以增強自身對光資源的競爭能力[34],同時,適度的遮蔭可緩解植物光抑制現象[48],增大葉綠素含量,進而提高光合利用效率[49],過度的遮蔭會引起植物體內養分供應障礙,光合速率受限[50]。已有研究發現禾本科牧草生長受到氮素的的限制,而豆科牧草生長往往依賴磷素[51],本研究結果顯示,混播紫花苜蓿凈光合速率、蒸騰速率、氣孔導度和胞間CO2濃度均高于單播,且隨紫花苜蓿比例的減小和禾草比例的增大呈增大的趨勢,說明混播系統中高稈、上繁草的紫花苜蓿具有光競爭優勢,同時禾本科牧草活化了土壤中的磷,促進了紫花苜蓿光合作用[52],草地早熟禾在7∶3豆禾比下光合作用低于單播,可能是因為過大的紫花苜蓿比例造成了草地早熟禾遮蔭脅迫,引起了養分供應障礙,使光合作用受限,其他混播處理禾草光合能力均高于單播,說明紫花苜蓿給禾草提供了適度的遮蔭,同時供給附近禾草氮素,進而緩解了禾草光制止現象,使禾草上調酶活性、酶量以及葉綠素等,提高了光合作用[53]。在本試驗中,豆禾混播系統中紫花苜蓿表現為高光效、高蒸騰作用,禾草均表現為低光效、低蒸騰作用,光合生理互補優勢明顯[54]。
本研究對不同混播處理草地牧草生理指標應用隸屬函數法進行綜合評價,結果顯示混播紫花苜蓿生理指標綜合評價D值均大于單播且在F1處理下最大,同時混播紫花苜蓿較單播增產且在F1處理下紫花苜蓿增產率最高;P1處理下草地早熟生理綜合評價D值小于單播,增產率最小,其他混播處理下草地早熟禾綜合評價D值大于單播,草地早熟禾增產率較P1處理增大;B2處理下無芒雀麥綜合評價D值最大,增產率最大;F1處理下葦狀羊茅綜合評價D值最大,增產率最大。
4 結論
混播紫花苜蓿和葦狀羊茅均具有較低的MDA含量和較高的抗氧化能力、滲透調節物質含量和光合能力,2020年過大的紫花苜蓿比例降低了草地早熟禾的抗氧化能力、滲透調節物質含量和光合能力以及無芒雀麥的抗氧化能力,適當減小紫花苜蓿比例能使草地早熟禾和無芒雀麥的生長生理指標好于單播。混播草地中紫花苜蓿表現為高光效、高蒸騰作用,禾草均表現為低光效、低蒸騰作用,二者光合生理協同效應明顯。紫花苜蓿與與叢生型葦狀羊茅7∶3混播時兩種牧草生理協同性最好,紫花苜蓿和葦狀羊茅均具有最高的增產率,混播草地生產力最高。
