基于葉綠體基因組分析我國苜蓿屬植物演化路徑
2022-08-31 06:08:24王曉娟董文攀周世良
生態學報 2022年15期
王曉娟,董文攀,周世良
1 上海科技館,上海自然博物館自然史研究中心,上海 200127 2 北京林業大學生態與自然保護學院,北京 100083 3 中國科學院植物研究所,系統與進化植物學國家重點實驗室,北京 100093
苜蓿屬(MedicagoLinn.)隸屬于豆科蝶形花亞科車軸草族,屬下物種經歷數次劃分調整,物種數量從最初的46種不斷增加至目前的87種,我國有15種[1—3]。苜蓿屬植物的分類困難與其起源、地理分布以及相似的易混淆物種有關,例如,區分苜蓿屬、胡盧巴屬(TrigonellaLinn.)和草木犀屬(MelilotusLinn.)的一些物種,一直是令人棘手的難題[4—5]。1977年,Ivanov[6]提出苜蓿屬植物的起源地在高加索地區,之后有學者認為是地中海北部海岸[1]。目前發現的苜蓿屬大多數物種分布于地中海沿線、阿拉伯半島、伊拉克和巴爾干東部地區,而部分成員如紫花苜蓿復合體中的少數種則因栽培逃逸擴展至中亞、北亞和東亞地區[2—3]。苜蓿屬植物為一年生或多年生草本,稀灌木,在整個生態系統乃至農牧生產系統中均具有重要的功能,廣泛分布于山地、農田、草地、森林等不同生態系統類型,其中,紫花苜蓿因其高蛋白、全營養和耐消化等優良特性,成為世界上最為重要的飼料作物之一[7];截形苜蓿因其為二倍體、自花授粉、種子量大和遺傳操作方便等特性,成為豆科植物研究的模式植物[8]。
據《中國植物志》記載,我國現有苜蓿屬植物15種,隸屬于木本苜蓿組、天藍苜蓿組、紫苜蓿組、闊莢苜蓿組和南苜蓿組,而《Flora of China》未做屬下等級的劃分[2]。鄧宏超等[4]研究了國內苜蓿屬和胡盧巴屬的分類劃分,并基于形態學特征等將苜蓿屬植物分為三類:一是開紫花而莢果螺旋盤卷,二是開黃花而莢果鐮刀狀彎曲或稍直,三是花雜色而莢果鐮刀狀彎曲至螺旋盤卷的過渡類型。但由于苜蓿屬植物耐受性強,分布廣泛,生活型多變,導致學界難以將苜蓿屬植物的進化歷史與其地理、生境特征直接關聯。如何通過科學方法探究苜蓿屬植物的生態演化路徑已經成為本學科關注的熱點之一。
隨著分子生態學的出現和發展,利用基因組學和分子生物學的方法解決生態學問題不斷報道,但目前我國有關苜蓿屬植物葉綠體基因組和系統發育方面的研究尚開展較少,通過葉綠體基因組揭示該屬植物的起源、演化方面的研究也少有報道。葉綠體是細胞內一類具有自主遺傳功能的重要細胞器。自1986年葉綠體基因組首次被測定以來[9—10],已有近6,549個物種的葉綠體基因組被測序,涵蓋大多數重要的真核光合類群(National Center for Biotechnology Information,NCBI,2021年6月8日登錄)。葉綠體基因組數據的迅速增加,表明本領域已經逐漸成為研究熱點,同時也為揭示葉綠體基因組在研究生態進化中的重要價值提供了依據。研究表明,葉綠體基因組不僅在探究物種起源、演化以及物種間親緣關系等方面具有重要貢獻[11—12],而且在作物遺傳改良、生物制劑生產等方面也顯示出了巨大潛力[13]。基于此,為了闡明我國苜蓿屬植物的分子生態學結構特征,探究其在我國的生態進化路徑,本研究選取我國10個苜蓿屬代表物種和葫蘆巴屬一個物種作為研究對象,采用高通量測序平臺測定其葉綠體全基因組和核基因ITS序列,通過分析其葉綠體基因組結構特征和變異規律,構建苜蓿屬植物的系統發育關系,結合上述苜蓿屬植物來源地的生境特征,探究我國苜蓿屬物種的葉綠體基因組進化式樣,揭示我國苜蓿屬植物的演化機理。
1 材料與方法
1.1 植物材料
苜蓿屬和胡盧巴屬植物種子來自中國科學院昆明植物研究所中國西南野生生物種質資源庫(Germplasm Bank of Wild Species,GBWS),截形苜蓿種子來自澳大利亞南澳研究開發院(South Austrulian Research and Development Institute,SARDI)牧草種質資源庫,每種供試材料的采集編號以及采集地的年均降雨量、年均日照時數、年均氣溫和氣候區特征見表1。
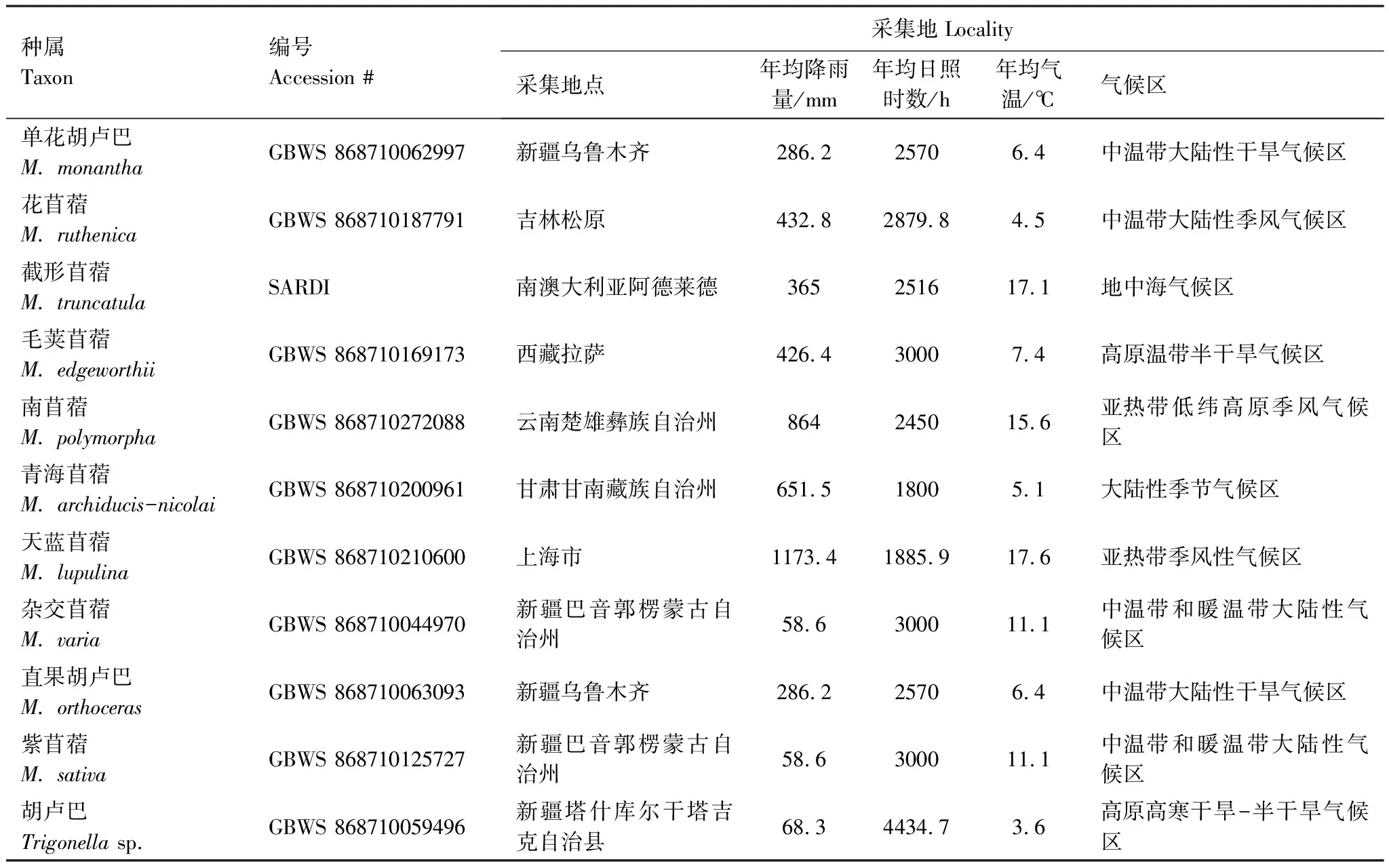
表1 取樣的苜蓿屬植物材料信息
1.2 溫室培養
2018年至2019年,表1中苜蓿屬和胡盧巴屬植物的種子于智能溫室(上海乾菲諾農業科技有限公司)進行萌發和培養。所有種子經溫水浸種8 h后,將種子置于25℃恒溫箱中催芽,每天投洗種子2次,直至種子露白后,進行播種。溫室栽培條件為溫度(25±2)℃,光照強度400 μmol m-2s-1,空氣濕度40%,始終保持基質濕潤。整個生育期內常規管理(圖1)。

圖1 供試苜蓿屬植物形態Fig.1 Medicago plants cultivated in green house1,紫苜蓿 M. sativa;2,雜交苜蓿 M. varia;3,截形苜蓿 M. truncatula;4,南苜蓿 M. polymorpha;5,天藍苜蓿 M. lupulina;6,青海苜蓿 M. archiducis-nicolai;7,花苜蓿 M. ruthenica;8,直果胡盧巴 M. orthoceras;9,單花胡盧巴 M. monantha;10,毛莢苜蓿 M. edgeworthii; 每個種各2張圖片,圖示各物種的葉片、花序、莢果或復葉等
1.3 DNA提取與高通量測序
采集幼嫩葉片立即置于液氮中冷凍后低溫保存。樣品總DNA采用mCTAB法提取,具體步驟參考李金璐等[14]的植物DNA提取方法。
總DNA采用超聲打斷,切膠回收350 bp的DNA片段,利用NEBNext?的建庫試劑盒構建350 bp大小的文庫,利用Hiseq 4000 PE150的測序平臺進行測序(北京諾禾致源科技股份有限公司,北京)。
1.4 葉綠體基因組和ITS序列拼接與注釋
高通量測序數據采用SPAdes3.6.1軟件進行質量控制和拼接,以已經發表的葉綠體基因組和ITS序列為參考,用Blast程序篩選出葉綠體基因組和ITS序列的contigs[15],篩選出的葉綠體基因組和ITS序列的contigs用Sequencher 4.10組裝成完整的葉綠體基因組和ITS序列。
利用Plann程序對獲得的葉綠體基因組進行注釋。利用已經公開發表的M.sativa(序列接收號:MK460489)為參考序列,對所有的蛋白質編碼基因,rRNA和tRNA基因進行注釋,并對注釋結果進行逐一檢查。缺失的基因或者自動注釋錯誤的基因,依照參考序列的注釋信息進行手動調整,得到完整的注釋結果。利用在線軟件OGDRAW(https://chlorobox.mpimp-golm.mpg.de/OGDraw.html)繪制苜蓿屬物種的葉綠體基因組物理圖譜。
1.5 簡單重復序列分析
利用MISA 軟件對苜蓿屬物種的葉綠體基因組的簡單序列重復(simple sequence repeats, SSRs)分別進行檢測。SSR位點的定義如下,單核苷酸單元的重復數≥10;二核苷酸單元的重復數≥5,三核苷酸單元重復數≥4,四核苷酸、五核苷酸和六核苷酸單元的重復數≥3。兩個SSRs之間的距離不少于20 bp。
1.6 系統發育分析
根據注釋結果,提取苜蓿屬葉綠體基因組的76個編碼基因,利用MAFFT 軟件進行比對,并利用MEGA X進行手工檢查。比對好之后串聯成一個數據集進行系統發育分析。用RAxML軟件以最大似然法進行系統發育分析。核苷酸替換模型采用GTR+G,系統發育各個分支長度的支持率通過進行500次自展重復分析獲得。
2 結果與分析
2.1 苜蓿屬葉綠體基因組結構特征
本研究測定了苜蓿屬10個物種完整的葉綠體基因組和1個胡盧巴屬物種的葉綠體基因組,平均測序深度從1,222×(青海苜蓿)到10,340×(花苜蓿)。供試材料的葉綠體基因組基本特征見表2。葉綠體基因組大小不盡相同,介于121—127 kb之間,其中,青海苜蓿葉綠體基因組最大,長度為127,358 bp,單花胡盧巴最小,長度為121,082 bp,兩者基因組大小相差約6 kb。GC含量介于33.8%—34.1%之間,平均值為34.02%,不同物種間GC含量差異較小,有5個物種的葉綠體GC含量完全一致,均為34.1%。

表2 苜蓿屬植物葉綠體基因組測序分析
由圖2可見,供試苜蓿屬和胡盧巴屬均非典型的“四區”結構,都缺失了一個大的反向重復序列,屬于豆科蝶形花亞科車軸草族反向重復序列缺失支(inverted repeat-lacking clade,IRLC)。依據組裝注釋的葉綠體基因組的結構特征,供試材料的葉綠體基因組分為以下4種類型:1)類型I最為普遍,包括圖2中所示的直果胡盧巴、單花胡盧巴、毛莢苜蓿、紫苜蓿、雜交苜蓿、南苜蓿和截形苜蓿;2)類型II,包括青海苜蓿和花苜蓿兩種;3)天藍苜蓿單獨作為類型III;4)胡盧巴單獨作為類型IV。對比后發現,四種類型的葉綠體基因組結構發生了不同程度的變化,包括基因的重排現象、片段丟失等。

圖2 苜蓿屬植物4種葉綠體基因組類型Fig.2 Four types of plastomes in Medicago plants最外圈的方框表示基因(圈內的表示順時針轉錄,圈外的表示逆時針轉錄);不同顏色表示不同功能的基因;陰影表示GC含量;類型I,直果胡盧巴、單花胡盧巴、毛莢苜蓿、紫苜蓿、雜交苜蓿、南苜蓿和截形苜蓿;類型II,青海苜蓿和花苜蓿;類型III,天藍苜蓿;類型IV,胡盧巴
供試材料的葉綠體基因組都含有111個不同的基因,包括77個蛋白編碼基因、30個tRNA基因和4個rRNA基因。其中,自我復制基因包括核糖體大亞基、核糖體小亞基的蛋白編碼基因、tRNA和rRNA基因;光合作用基因包括光系統I、光系統II、NADH脫氫酶、細胞色素b/f復合物、ATP合成酶和RubisCo大亞基的蛋白編碼基因;其他基因包括成熟酶K、囊膜蛋白、乙酰輔酶A羧化酶亞基、C-型細胞色素酶、蛋白酶基因以及未知功能蛋白編碼基因(表3)。

表3 苜蓿屬葉綠體基因組的基因類型、基因群和基因名稱
2.2 苜蓿屬物種葉綠體基因組簡單序列重復
由圖3可見,苜蓿屬葉綠體基因組的簡單序列重復數為102到129個,單堿基重復出現次數最多(62.14%),其次為二堿基重復(21.29%)。單堿基重復多是A/T,二堿基重復則多為AT/TA。

圖3 苜蓿屬物種葉綠體基因組簡單重復序列的數量及其分布特征Fig.3 Numbers and characterization of simple sequence repeat (SSR) in Medicago plastomes
2.3 苜蓿屬物種葉綠體基因組結構變異
如圖4所示,在供試苜蓿屬和胡盧巴屬的葉綠體基因組中,共發現10個倒位的結構變異:1)天藍苜蓿中2個,發生在基因atpB到ycf3、ndhC到trnLUAA之間的區域;2)青海苜蓿和花苜蓿共享4個倒位,發生在從基因psbM到psaA、ndhB到trnNGUU、ndhB到rpoA和clpP到rpl20之間的區域;3)胡盧巴中6個,發生在基因accD到trnPUGG、ndhB到rpoA、rpl20到psbF、ndhB到trnNGUU、psbJ到cemA以及psbB到ycf1之間的區域。其中,胡盧巴發生倒位變異最多,6處倒位既包括從psbB到ycf1共包含32個基因的長序列區段,也有從psbJ到cemA僅包含3個基因的短序列區段。其中,天藍苜蓿獨立發生2處倒位,而在青海苜蓿、花苜蓿和胡盧巴3個物種中則平行共享以下2處倒位:ndhB到trnNGUU(包含12個基因)、ndhB到rpoA(包含14個基因)。

圖4 基于葉綠體編碼基因的苜蓿屬植物系統發育關系和葉綠體基因組結構變異Fig.4 Phylogenetic relationships based on plastid coding genesand distribution of plastome structural mutations內含子丟失和倒位繪制在分支圖的分枝上;右上角的小圖表示帶枝長的系統發育樹,圖中數字表示所在節點的支持率,未標注數字的節點的支持率均為100
葉綠體基因組序列分析還在天藍苜蓿中發現了內含子丟失現象。3個發生內含子丟失的基因分別是clpP、rpoC1和atpF,其功能分別為編碼蛋白酶、DNA的RNA聚合酶以及ATP合成酶(圖4)。
2.4 基于葉綠體基因組序列和核ITS序列的苜蓿屬系統發育關系
基于葉綠體基因的系統發育關系顯示4個主要譜系分支,從基部開始分別包含毛莢苜蓿、直果胡盧巴和單花胡盧巴、青海苜蓿和花苜蓿以及其他5個種。基于核基因ITS的系統發育關系與以上結果略有不同,主要是天藍苜蓿單獨形成分支,而不是與截形苜蓿等組成一個分支。ITS系統發育分化式樣與傳統的5個組的劃分吻合。
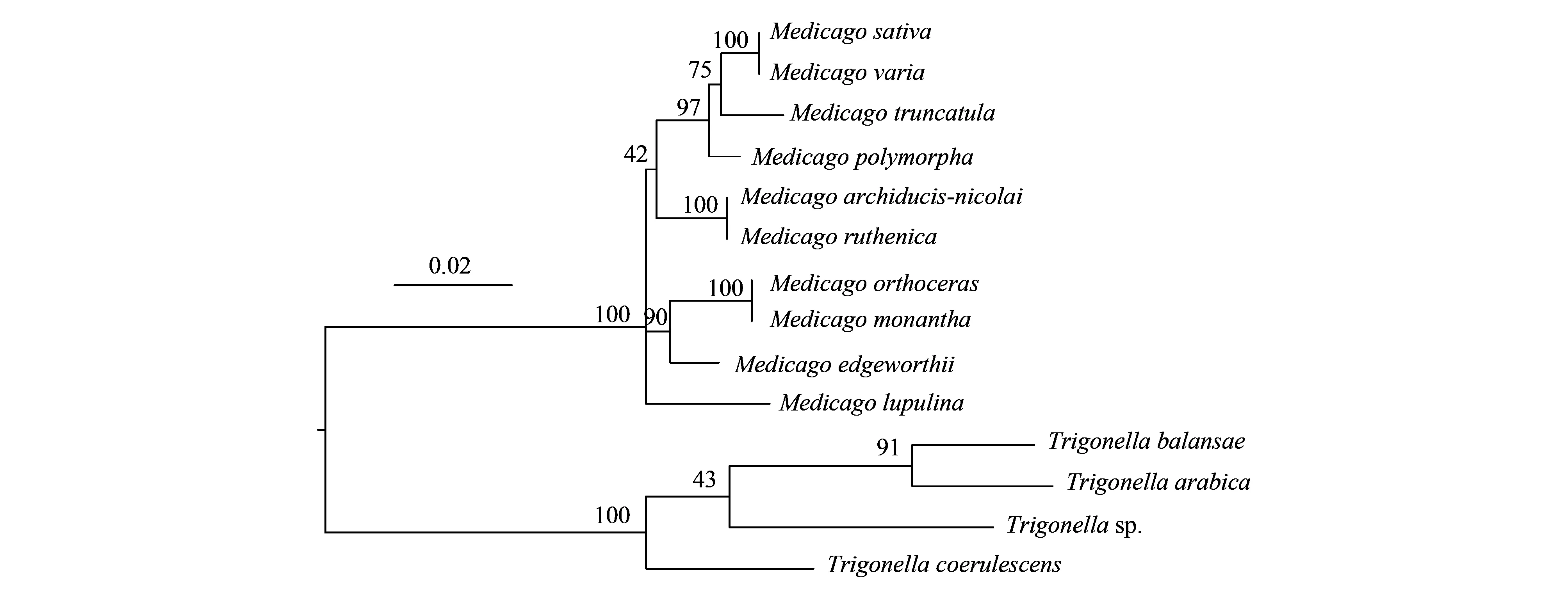
圖5 基于rDNA的ITS序列的苜蓿屬植物系統發育樹 Fig.5 Phylogenetic tree of Medicago species based on rDNA ITS sequences
3 討論
3.1 反向重復序列丟失和IRLC類群
作為分子生態學研究的基礎,分子測序技術的快速發展,直接推動了葉綠體基因組學的發展[16]。典型陸生植物葉綠體基因組為雙鏈環狀DNA分子,大小一般為115—165 kb,常編碼約101—118個基因。這些基因主要分為3類,即光合作用有關基因、基因自身表達有關的基因和其他生物合成有關的基因。植物的葉綠體基因組結構一般具有相對保守的“四區”結構:大單拷貝區(large single copy region,LSC)、小單拷貝區(small single copy region,SSC)和兩個序列完全相同、方向相反的反向重復區(inverted repeat region,IR)。在被子植物中,反向重復區域的基因高度保守,堿基的突變速率也低于單拷貝區,研究表明IR區的存在對于穩定葉綠體基因組結構起到重要作用[17]。IR區主要編碼4個rRNA基因(rrn16、rrn23、rrn4.5 和rrn5)和一些未知功能基因(ycf)[18]。被子植物葉綠體基因組的“四區”結構相對保守,但IR區域可擴張或縮減,是影響葉綠體基因組大小的重要因素之一,導致不同植物類群的葉綠體基因組差異很大[19]。例如,發生在豌豆(Pisumsativum)中IR完全刪減[20]和天竺葵(Pelargoniumhortorum)中IR極度增加(增至76 kb)[21]的極端報道。此外,已報道的樟科葉綠體基因片段丟失事件,導致該科植物葉綠體基因組整體縮小,并且和樟科種系發生進程一致[22]。
物種的進化必然是建立在基因組的進化之上,物種多樣性是生態、進化和歷史過程的產物,其演化路徑可能既有宏觀上的方向性,也有微觀成種的隨機性。葉綠體基因組的縮減、擴張及丟失可能與物種在進化過程中的物種分化和譜系分化相吻合,當然,兩者也不一定完全等同[23]。目前,盡管對植物葉綠體基因組的結構和功能有所認識,但有關葉綠體基因組結構變異與其生態進化起源、演化驅動機制等尚待深入研究。自Kolodner & Tewari[24]首次在豌豆葉綠體基因組中發現IR丟失現象,之后的研究又陸續在綠豆(Vignaradiata)、大豆(Glycinemax)、菜豆(Phaseolusvulgaris)和蠶豆(Viciafaba)中證實了其葉綠體基因組IR缺失現象[25—26]。隨著葉綠體基因組數據的不斷積累,豆科蝶形花亞科車軸草族物種的葉綠體基因組發生IR區的丟失現象,成為該類植物的指示特征,為此,該類群也被稱為反向重復序列缺失支,并且IR丟失事件(該支的分化事件)發生在—3900萬年前[27—30]。如圖2 所示,供試苜蓿屬及其近緣屬胡盧巴也呈現IR區域的丟失,均為IRLC類成員(圖2)。越來越多的證據發現,IR區段丟失現象還發生在其它植物類群中,如牻牛兒苗科(Geraniaceae)的老鸛草屬(Geranium)[31]和仙人掌科(Cactaceae)的仙人掌(Carnegieagigantea)[32]中,進一步表明IR區的丟失可能在被子植物譜系中多次獨立發生,而葉綠體基因組大的結構變異在特定類群因具有強烈的系統發育信號,常被用來定義單系類群[33]。
3.2 苜蓿屬葉綠體基因組結構變異的進化啟示
包括苜蓿屬在內的豆科IRLC類群是IR收縮的經典案例,目前認為大的IR收縮可能與基因組的異常重組有關[34]。研究發現,小苜蓿(M.minima)中一段新的IR位于ycf1和clpP附近,而這兩個基因序列通常被認為是高度變化的,并可能是基因重組的熱點區域[23]。堿基替換和結構變異引起的基因重組事件頻發也可能導致豆科植物IR收縮的發生[35]。葉綠體基因組的大小除了受IR擴張和收縮的影響,還與重復序列的含量相關。本研究測定的我國苜蓿屬植物的葉綠體基因組簡單序列重復數為102到129個,單堿基重復是出現次數最多的(62.14%),可作為分子標記,為研究苜蓿系統學和進化生物學提供依據。然而,葉綠體基因組的穩定性和重復序列累積以及IR缺失或是獲得之間的關系尚未明了,特別是IR擴張、收縮或缺失等結構變異對核酸替換率的影響尚不清晰。
除反向重復序列(IR)的缺失,供試苜蓿屬物種中還發現了其它顯著的葉綠體基因組結構變異,如天藍苜蓿clpP、rpoC1和atpF基因的內含子丟失、青海苜蓿和花苜蓿之間共享倒位等,這些獨立或是平行發生的結構變異,也是潛在的系統發育信號,可以作為區分不同類群的重要特征(圖4)。研究發現,豆科菜豆亞族的刺槐屬(Robinia)、綠豆、菜豆等物種,其葉綠體基因組發生50 kb倒位,和其它具有該結構變異的類群構成了50 kb倒位支[36—38]。事實上,包括苜蓿屬在內的核心車軸草族的葉綠體基因組也發生了不同程度的重排,包括基因的重復、丟失和序列的倒位等[39]。大于1kb的倒位在蝶形花亞科植物中較為常見,本研究中青海苜蓿和花苜蓿倒位發生在基因psbM到psaA、ndhB到trnNGUU、ndhB到rpoA以及clpP到rpl20之間,而近緣屬的胡盧巴倒位變異則高達6處,包括psbB到ycf1的區段(含32個基因),以及psbJ到cemA區段(含3個基因),顯示出葉綠體基因組結構變異的多樣性和復雜性。
本研究結果發現,我國天藍苜蓿中的內含子丟失與其它研究報道相一致,有必要通過更加廣泛的取樣和分析,進一步評估該分支苜蓿物種的倒位、內含子丟失及其演化模式[23]。研究表明,反向重復序列可能通過介導分子內重組而導致倒位的發生[28]。缺乏反向重復區的豆科蝶形花亞科IRLC物種如何控制葉綠體基因組穩定性還遠未被闡明,或許對苜蓿屬廣泛的葉綠體基因組分析可以找出結構穩定性與多樣性的關系,從而揭示IR多次得失的變異機制。
3.3 我國苜蓿屬植物的演化路徑
我國野生苜蓿分布在全國不同地理區域,適應各種生境特征,從寒冷的北部沙漠到溫暖潮濕的南部山區和中部平原地帶,從國內東部沿海到喜馬拉雅山脈[2]。這些野生種擁有豐富的自然變異來源,是了解苜蓿屬植物種群動態和改善苜蓿品種的重要基礎。對于植物的生長和分布來說,溫度、降水和光照是最重要的氣候因子,強烈地影響著物種的分布格局(表1)。由本實驗測序結果(圖4),對照表1生境特征可知,苜蓿屬植物的進化與年平均溫度、年均降雨量、年均日照時數等生境因子之間存在密切關系,特別是年平均溫度影響最大。年均溫度越高,物種出現的時間越晚,進化程度越高,如紫苜蓿(11.1℃)、截形苜蓿(17.1℃)、南苜蓿(15.6℃)、天藍苜蓿(17.6℃),反之,處于進化分支樹基部的物種,其生境條件均相對較為惡劣,如胡盧巴(3.6℃)、毛莢苜蓿(7.4℃)、單花胡盧巴(6.4℃)、花苜蓿(4.5℃)、青海苜蓿(5.1℃),表明溫度是苜蓿屬植物物種進化的重要影響因子,溫度越高,越利于苜蓿屬植物的生長,有利于其拓展更廣闊的生境,進而成為廣布種。其中,天藍苜蓿是一年生、二年生或偶爾短命的多年生植物,自交為主(自交率大于95%),分布廣泛,是易于繁衍的綠肥植物;而花苜蓿則是長壽的多年生植物,行異交(自交率低于30%),分布狹窄,適應干燥極寒的砂礫和沙漠等低投入生態系統。此外,天藍苜蓿具有不裂的小豆莢,通過生物和非生物因子促進種子的長距離傳播,而花苜蓿具開裂的豆莢并缺乏有效的種子傳播機制。從表型上看,天藍苜蓿的花相對較小,具2—4 mm的黃色蝶形花冠,而花苜蓿則有長達8 mm的黃色花冠,更大更艷麗,能夠吸引更多的昆蟲訪花,如蜜蜂、大黃蜂等。然而,隨著快速城市化和過度放牧,國內的野生苜蓿資源受到人為干擾和城市化進程的影響,種群數量和規模大幅減少,其野生種質資源調查和保護需引起重視。
苜蓿屬和胡盧巴屬植物具有相似的特征,長期以來,二者之間的進化分類界限不明顯,部分物種的歸屬存在爭議,兩個屬之間存在“類苜蓿植物(Medicagoid)”或“類胡盧巴植物(Trigonelloid)”[40]。鄧宏超等[4]開展了苜蓿屬和胡盧巴屬植物形態特征及數量分類研究發現,直果胡盧巴和單花胡盧巴屬于角形果組,聚類結果顯示它們與苜蓿屬植物之間存在較近的親緣關系,建議進一步開展深入研究確定其能否歸并到苜蓿屬。而早期Small等[41]也曾提出將胡盧巴歸至角形果組并合并到苜蓿屬中。本研究基于葉綠體基因組及ITS序列重新構建了苜蓿屬系統發育關系,我國苜蓿屬物種的基因分化式樣與傳統分類的紫苜蓿組、南苜蓿組、闊莢苜蓿組和天藍苜蓿組基本吻合。其中,直果胡盧巴和單花胡盧巴與毛莢苜蓿形成一枝,分子證據支持將苜蓿屬和胡盧巴屬之間的過渡類型“類苜蓿植物”獨立出來(圖4和圖5)。由此可見,葉綠體基因組大量的系統發育信息位點,不僅有助于闡明苜蓿屬和胡盧巴屬之間的系統發育關系,而且可建立高分辨率的苜蓿屬和胡盧巴屬種間的生態進化關系。生命形式、交配系統和種子傳播是影響苜蓿屬植物種群的遺傳結構塑造和地理分布的適應性性狀。以上苜蓿屬植物葉綠體和核基因組分析連同生態、表型數據可為該屬植物的演化路徑提供依據,并有助于確定自然種群的進化潛力和保護策略。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
