單葉薔薇幼苗根系對不同潛水埋深的適應(yīng)機制
2022-08-31 06:02:14張曉龍劉學(xué)森鄭玲娜
生態(tài)學(xué)報 2022年15期
關(guān)鍵詞:生長
張曉龍,鄧 童,劉學(xué)森,鄭玲娜,于 超,6,羅 樂,6,*
1 北京林業(yè)大學(xué)園林學(xué)院,北京 100083 2 城鄉(xiāng)生態(tài)環(huán)境北京實驗室,北京 100083 3 花卉種質(zhì)創(chuàng)新與分子育種北京市重點實驗室,北京 100083 4 教育部園林環(huán)境工程研究中心,北京 100083 5 林木花卉遺傳育種教育部重點實驗室,北京 100083 6 國家花卉工程技術(shù)研究中心,北京 100083
根系是植物在適應(yīng)陸地生活中逐漸進化形成的器官[1],具有固定植株、吸收水分養(yǎng)分以及合成和貯藏營養(yǎng)物質(zhì)、激素并向地上部分輸送的功能[2]。根系的生長狀況與分布特征可以反映土壤中物質(zhì)能量的吸收利用程度[3],體現(xiàn)不同地理環(huán)境下植物的生態(tài)適應(yīng)對策[4]。根系形態(tài)、拓?fù)浣Y(jié)構(gòu)、生長速度、生長角度和扭曲程度等特征都屬于根系構(gòu)型研究的范疇[5]。有研究表明,不同植物根系在異質(zhì)環(huán)境下的適應(yīng)性表現(xiàn)不一,如沙柳(Salixpsammophila)作為深根系植物,但既可利用淺層土壤水又能吸取地下水,而烏柳(Salixcheilophila)、檸條(Caraganakorshinskii)等淺根系植物,根冠比小,主要水源是降水形成的淺層土壤水[6]。疏葉駱駝刺(Alhagigagnebin)可以通過增加垂直根系長度、擴大根冠比來應(yīng)對深層潛水帶來的干旱脅迫[7]。面對高中低水位時,檉柳(Tamarixchinensis)根系拓?fù)浣Y(jié)構(gòu)由叉狀分支向人字形分支模式過渡,表明其具有良好的環(huán)境適應(yīng)性和可塑性[8]。因此,開展不同潛水埋深條件下植物根系構(gòu)型研究,對于理解植物在不同生境中的資源配置機制和生態(tài)適應(yīng)性策略具有重要意義。
目前,限制我國西北干旱、半干旱地區(qū)植物生長發(fā)育的主要因子為水分,在降水稀缺的干旱區(qū)自然生態(tài)系統(tǒng)中,地下水的時空分布又將直接或間接影響生態(tài)系統(tǒng)過程、植物群落組成及植物用水策略[9—10]。雖然近60年來西北地區(qū)氣候暖濕化明顯[11],但年極端高溫事件發(fā)生頻次亦為增加趨勢[12],且未來氣候變化具有暖干化的潛在特點[13],所以,無論當(dāng)前還是未來,地下水對西北植被的生存都具有重要意義。
單葉薔薇(Rosapersica),亦稱為小檗葉薔薇(Rosaberberifolia,異名),是薔薇科(Rosaceae)薔薇屬(RosaL.)多年生落葉叢生灌木,在我國主要分布在新疆西北部地區(qū),以建群種的形式生長于海拔120—950 m的山坡、荒地或路旁等干旱地帶的灌叢群落中,具有抗寒、抗旱、耐鹽堿、根系發(fā)達等優(yōu)點[14],是用于防風(fēng)固沙、防止水土流失和控制荒漠化的優(yōu)良植物。近年由于北疆地區(qū)開墾農(nóng)田、擴建道路、興修水渠等人為活動,單葉薔薇生境急劇惡化,種群面積和數(shù)量迅速減少,若不進行及時保育,則該物種瀕危風(fēng)險將進一步加大[15]。作為國家二級保護植物,目前單葉薔薇的研究主要集中在地上器官形態(tài)學(xué)[16—18]、生境調(diào)查與群落特征[19—21]、分類學(xué)[22]、孢粉學(xué)[23]以及生物學(xué)特性[24]等方面,根系研究探討較少,特別是異質(zhì)性環(huán)境下根系構(gòu)型的適應(yīng)性機制尚不明晰;在野外,成年植株地下直根可達170 cm以上,且直根上有大量的地下莖,并靠地下莖進行無性繁殖,這種形態(tài)發(fā)育特征有利于其在干旱條件下充分利用土壤水分進行生長[15,18]。因此,本研究以單葉薔薇幼苗為試驗材料,目的是對比不同潛水埋深條件下單葉薔薇幼苗生長發(fā)育、根系形態(tài)、拓?fù)浣Y(jié)構(gòu)與分形維數(shù)以及表型可塑性的變化,初步闡明其根系的生長適應(yīng)性策略。研究結(jié)果可補充單葉薔薇根系生態(tài)學(xué)研究,對該物種的繁育也具有一定指導(dǎo)作用,亦可為今后單葉薔薇植被生態(tài)修復(fù)、抗逆性研究提供理論基礎(chǔ)。
1 材料和方法
1.1 試驗設(shè)計
試驗以一年生單葉薔薇播種苗為材料,種子采自新疆野外,選取健康飽滿的種子進行層積催芽。2021年1月15日播種,4月1日開始設(shè)置不同的潛水埋深進行試驗。
試驗容器為直徑16 cm的PVC管,對照組為70 cm高的種植土柱;結(jié)合前期預(yù)試驗情況和參照胡楊幼苗等試驗設(shè)置[8,25],本試驗組設(shè)置5個不同的種植土柱高度:30、40、50、60、70 cm,底部均設(shè)高度為10 cm潛水層,即為5個不同潛水埋深梯度,供試土壤(河沙∶田園土∶草炭土∶珍珠巖∶石子為2∶6∶2∶1∶1)與潛水層之間采用石子、帶孔塑料板隔離(圖1)。處理編號依次為CK、D30、D40、D50、D60、D70。5個取樣時間:第15、30、45、60、75天,每個處理6株幼苗,共180株單葉薔薇幼苗。
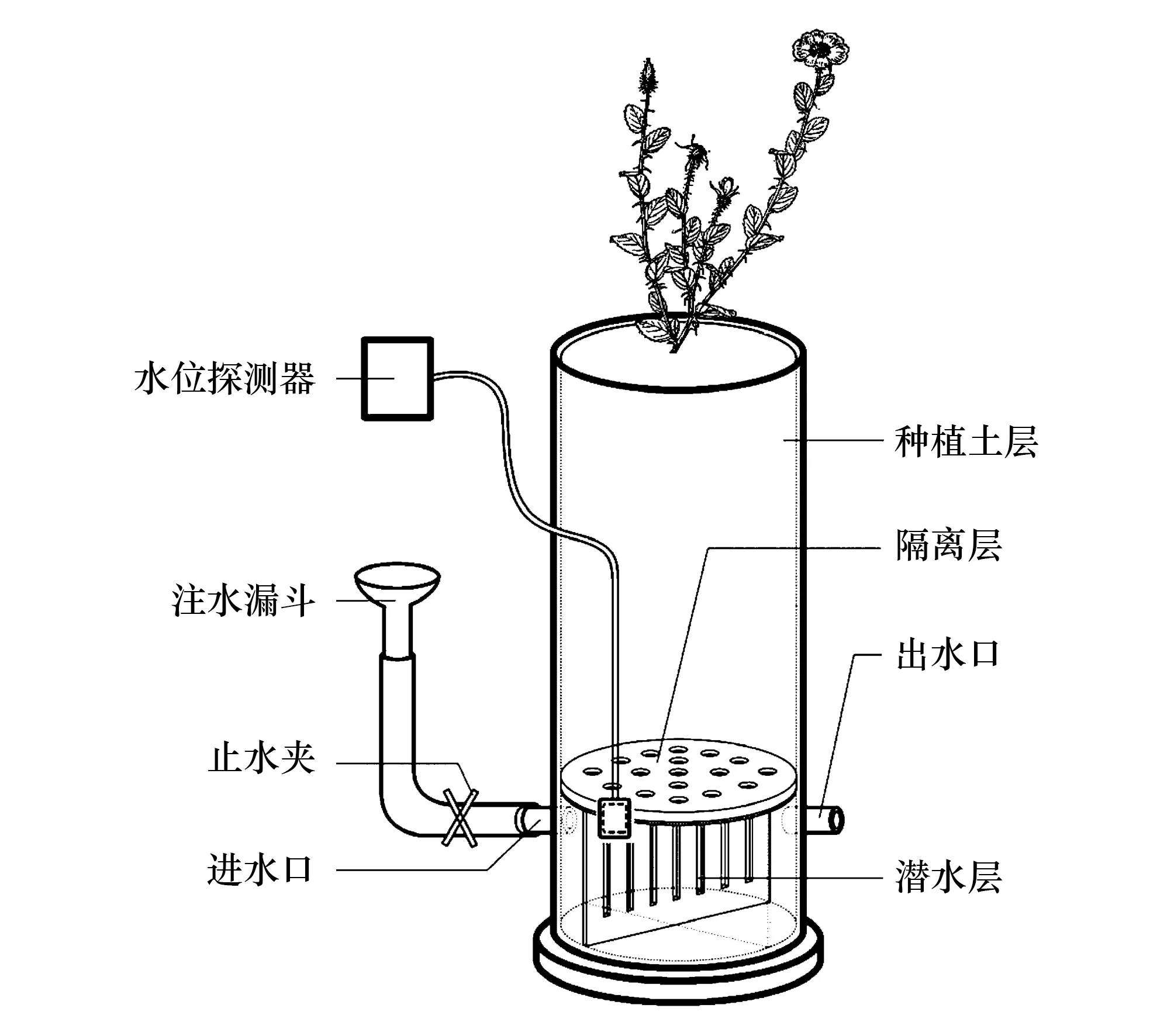
圖1 試驗裝置示意圖Fig.1 Schematic diagram of experiment device
試驗于北京林業(yè)大學(xué)小湯山基地的塑料大棚中進行,整個試驗期間不對單葉薔薇幼苗進行施肥、灌溉,每7天定時補充潛水層。
1.2 測定方法
分別于試驗開始后的第15、30、45、60天和第75天進行破壞性取樣,每個處理選取6株生長程度相近的健康個體作為樣株,用于試驗分析。取樣前用卷尺進行株高測定。取樣后,用水洗凈根系,將植株從基部剪開,向塑料盤中倒入少許蒸餾水并將根系平鋪在塑料盤中,用鑷子調(diào)整根系位置避免交叉重疊,使用臺式掃描儀(ScanMaker I800 Plus)進行掃描(分辨率600 dpi),后用根系分析系統(tǒng)(RhizoVision Explorer 2.0.2)處理掃描的圖片,獲得根系總長度、根表面積、根體積、根系內(nèi)部連接數(shù)量等相關(guān)指標(biāo)。分析完畢后用蒸餾水洗凈根系,用濾紙吸干水分,將地上部分與根系分別放入烘箱105 ℃下殺青15 min,再于65—75 ℃下烘干至恒重,用天平稱量其干重。從以下四個層次分析相關(guān)指標(biāo):
(1)生長發(fā)育:株高、扎根深度、垂直根系生長速率、地上干物質(zhì)、根系干物質(zhì)、根冠比、根尖數(shù)量;(2)根系形態(tài):總根長、根表面積、根體積、分支數(shù)量、比根長、比根表面積、根組織密度、分支密度,本研究以直徑級別進一步劃分細(xì)根(d<2.0 mm):0—0.50 mm(G1)、0.50—1.00 mm(G2)、1.00—2.00 mm(G3)[26],探究不同徑級根系之間的關(guān)系;(3)拓?fù)浣Y(jié)構(gòu)與分形維數(shù):拓?fù)渲笖?shù)、分形維數(shù);(4)可塑性分析:表型可塑性指數(shù)。
其中,比根長=總根長/根系干重;比根表面積=根表面積/根系干重;根組織密度=根系干重/根體積;分支密度=分支數(shù)量/總根長;拓?fù)渲笖?shù)Ti=lgA/lgM,Ti值越接近于1,根系越接近人字形分支模式,Ti值越接近于0.5,根系越接近叉狀分支模式;表型可塑性指數(shù)=(性狀的最大值-性狀的最小值)/性狀的最大值,范圍為0—1,值越大表示表型可塑性越大。
1.3 數(shù)據(jù)處理
利用Microsoft Office Excel 2021整理數(shù)據(jù),IBM SPSS Statistics 25.0進行單因素(ANOVA)分析, 鄧肯法(Duncan)分析顯著性(P<0.05),使用Origin 2021繪圖,各圖中的數(shù)值均為平均值±標(biāo)準(zhǔn)誤差。
2 結(jié)果與分析
2.1 不同潛水埋深對單葉薔薇幼苗生長發(fā)育的影響
隨著時間變化,不同潛水埋深對單葉薔薇幼苗株高、扎根深度、根尖數(shù)量、地上干物質(zhì)、根系干物質(zhì)、根冠比的影響日益顯著(圖2),對垂直根系生長速率影響不顯著(表1)。所有處理下的幼苗株高整體上均呈上升趨勢,其中D30、D40處理下的單葉薔薇幼苗株高增長尤為明顯,至第75天時,兩組較CK分別增長86.78%、75.05%,D50、D60、D70組株高較CK變化不顯著。對于扎根深度,相比第60天時,D30處理在第75天時下降13.00%,而其他處理下的單葉薔薇幼苗根系在整個生長季內(nèi)均表現(xiàn)出不同程度的增長,D30、D40、D50處理下的扎根深度分別在第45、60、75天時穿過隔離層到達潛水層;相比CK,扎根深度增幅最大出現(xiàn)在第75天時的D50處理組(111.00%)。第75天時D30處理下扎根深度相對下降,該處理在S5階段內(nèi)垂直根系生長速率表現(xiàn)出負(fù)增長(4.59 mm/d);相比CK,試驗組所有的垂直根系生長速率在S1、S2階段內(nèi)差異顯著,在S3、S4、S5階段內(nèi)不顯著;CK、D30、D40、D50、D60、D70垂直根系生長速率分別在S2、S4、S2、S5、S2、S5階段內(nèi)達到最大值,依次為5.55、14.30、10.17、10.42、11.47、8.12 mm/d(表1)。
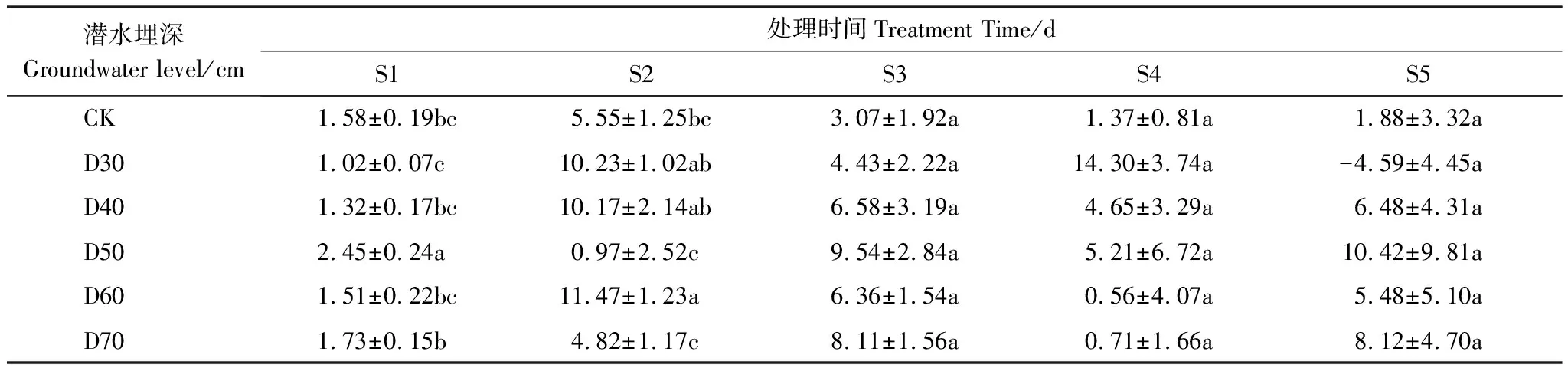
表1 不同階段單葉薔薇幼苗垂直根系生長速率/(mm/d)
就地上干物質(zhì)、根系干物質(zhì)而言,整個生長季內(nèi)不同處理下兩項指標(biāo)均呈增長趨勢,顯著差異性出現(xiàn)在試驗中后期(圖2)。第45天時,僅D40組地上干物質(zhì)量(0.0703 g)較CK(0.0392 g)差異顯著;D40、D50、D60、D70組的根系干物質(zhì)量依次為0.0217 g、0.0244 g、0.0268 g、0.0230 g,較CK(0.0086 g)差異顯著,D30(0.0170 g)與CK之間差異不顯著。第60天時,僅D30、D60兩組的地上干物質(zhì)量(0.1766 g、0.1422 g)、根系干物質(zhì)量(0.0605 g、0.0641 g)較CK(0.0326 g、0.0165 g)差異顯著。第75天時,D30、D40、D60組的地上干物質(zhì)量依次為0.4397、0.1966、0.2128 g,較CK(0.0639 g)差異顯著,其他處理與CK之間差異不顯著,根系干物質(zhì)量僅有D30、D60兩個處理較CK(0.0269 g)差異性顯著,為0.1547、0.1162 g。此外,相較CK,單葉薔薇幼苗地上干物質(zhì)、根系干物質(zhì)同是在第75天時的D30處理下增幅最大,為588.11%、475.46%;整個生長季內(nèi),CK處理下單葉薔薇幼苗地上干物質(zhì)、根系干物質(zhì)分別增長315.58%、375.81%,D30處理下分別增長1940.52%、1641.53%,D40處理下分別增長747.82%、564.67%,D50處理下分別增長640.08%、757.26%,D60處理下分別增長937.80%、1288.79%,D70處理下分別增長244.35%、409.20%,表明D30處理有利于單葉薔薇幼苗的生長,同時也證明單葉薔薇幼苗具有極強的抗旱能力,在完全干旱的條件下仍能繼續(xù)生長。
不同潛水埋深對單葉薔薇幼苗根冠比的顯著性影響僅出現(xiàn)在第45天和第60天(圖2)。第45天時,D30、D50、D60和D70處理下的根冠比較CK差異性顯著,分別增加115.38%、81.40%、81.83%、66.80%;第60天時,僅有D40處理下的根冠比與CK存在顯著性差異,下降47.81%。整體而言,D40處理下的單葉薔薇幼苗根冠比為W型變化趨勢,D30、D50、D60和D70均為“先升后降再升”的N型趨勢,而CK呈“先降后增再降”趨勢。
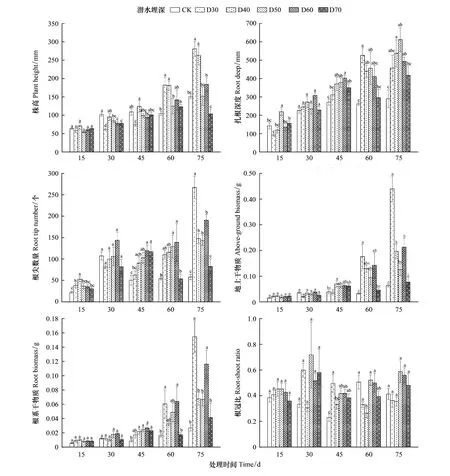
圖2 單葉薔薇幼苗生長指標(biāo)對不同潛水埋深的響應(yīng)Fig.2 Response of growth indexes of Rosa persica seedlings to different groundwater levels圖中不同小寫字母表示不同組之間差異顯著(P<0.05)
第15天時,D30、D40處理下的單葉薔薇幼苗根尖數(shù)量較CK存在顯著性差異;第45天時,D50、D60、D70組與CK存在顯著性差異;第60天時,D50和D60組較CK、D70存在顯著性差異;第75天時,D30、D40、D50、D60處理下的根尖數(shù)量與CK處理存在顯著性差異,D70與CK之間無顯著性差異。整個生長季內(nèi),試驗組和對照組處理下的單葉薔薇幼苗根尖數(shù)量均表現(xiàn)出“先升后降再上升”的趨勢,CK處理下的根尖數(shù)量從22.33增至58.00個,D30從37.83增至267.00個,D40從52.83增至147.69個,D50從36.50增至143.36個,D60從35.33增至190.55個,D70從30.17增至82.36個。
2.2 不同潛水埋深對單葉薔薇幼苗根系形態(tài)的影響
不同潛水埋深對單葉薔薇幼苗總根長、根表面積、根體積和分支數(shù)量均有顯著性影響,且時間越久,不同處理之間的差異愈加明顯。第75天時,D30組在總根長、根表面積、根體積和分支數(shù)量與其他處理均為顯著差異,四項指標(biāo)較CK分別增長3.94、5.93、10.36、4.57倍,是所有處理中增幅最大的一組試驗處理;D40、D50和D60處理下的四項指標(biāo)均顯著高于CK;而D70與CK之間差異不顯著。整個生長季內(nèi),CK處理下的幼苗在總根長、根表面積、根體積上始終保持“先升后降再上升”的N型變化趨勢,而D70處理下的幼苗在這三個指標(biāo)上始終保持“先降后升再降再升”的W型趨勢;在分支數(shù)量這一指標(biāo)上,CK和D70處理均為N型趨勢(圖3)。不同潛水埋深對單葉薔薇幼苗比根表面積無顯著影響(圖3)。第60天時D30組的比根長與D50、D60存在顯著差異,第75天時D70組比根長與D30、D60存在顯著差異。第45天的D30、D60組,第60天的D30、D40組均在根組織密度上與CK有顯著差異;第75天時,D30、D40、D50和D60組與CK之間均存在顯著差異。對于分支密度,僅第60天時的D50、D60與D30、D70之間存在顯著差異。不同處理下單葉薔薇幼苗的比根長、比根表面積總體上呈下降趨勢,而比根體積、分支密度為增長態(tài)勢。

圖3 單葉薔薇幼苗根系形態(tài)指標(biāo)對不同潛水埋深的響應(yīng)Fig.3 Response of root morphological indexes of Rosa persica seedlings to different groundwater levels圖中不同小寫字母表示不同組之間差異顯著(P<0.05)
與整個植株的總根長、根表面積、根體積相比,單葉薔薇幼苗各徑級細(xì)根三項指標(biāo)所占比例均隨潛水埋深的變化而發(fā)生改變(圖4)。試驗期內(nèi),幼苗G1級細(xì)根總根長占植株總根長的比例均呈整體上升趨勢,D70增幅最大(29.51%),CK緊隨其后(26.45%),而G2+G3級細(xì)根總根長所占比例均呈下降趨勢。針對根表面積占比、根體積占比,G1級細(xì)根在不同潛水埋深影響下表現(xiàn)出不同趨勢,基本可以分為兩類:D70、CK處理下的G1級細(xì)根表面積占比分別增長了12.49%、7.96%,根體積占比分別增長了20.94%、13.04%,而其他處理在兩項指標(biāo)上均為負(fù)增長,其中D30組的G1級細(xì)根表面積占比、根體積占比均是最大降幅(8.29%、16.25%)。由于本試驗中的幼苗粗根(d≥2 mm)數(shù)量較少,試驗苗根系主要由細(xì)根(d<2 mm)構(gòu)成,所以在進一步分級的情況下G1與G2+G3級根系在上述指標(biāo)上表現(xiàn)出相反的變化趨勢。

圖4 不同潛水埋深下單葉薔薇幼苗不同徑級根系性狀所占比例變化Fig.4 Changes in the proportion of root traits of different diameter classes in Rosa persica seedlings under different groundwater levelsCK,對照組 Control check;D30、D40、D50、D60、D70,均為試驗組,潛水埋深依次是30、40、50、60、70 cm;圖中不同小寫字母表示不同組之間差異顯著(P<0.05)
2.3 不同潛水埋深對單葉薔薇幼苗根系拓?fù)浣Y(jié)構(gòu)和分形維數(shù)的影響
如圖5所示,試驗中后期的單葉薔薇幼苗根系在拓?fù)浣Y(jié)構(gòu)和分形維數(shù)上存在顯著差異。D30處理下的拓?fù)渲笖?shù)從0.85(第15天時)下降至0.81(第75天時),CK組降幅0.03,其他處理下的拓?fù)渲笖?shù)基本保持穩(wěn)定。整體上不同處理下的拓?fù)渲笖?shù)保持在0.8—0.9之間,接近1,說明單葉薔薇幼苗根系屬于典型的人字形分支模式,受環(huán)境影響小。單葉薔薇幼苗分形維數(shù)經(jīng)歷了“先增后降再增”的趨勢,相比第15天,第75天時的CK和D70處理下分形維數(shù)分別下降了0.03、0.04,而其他四個處理均是微增,整體上分形維數(shù)保持在1.25—1.45之間,D30與CK、D70之間差異顯著。
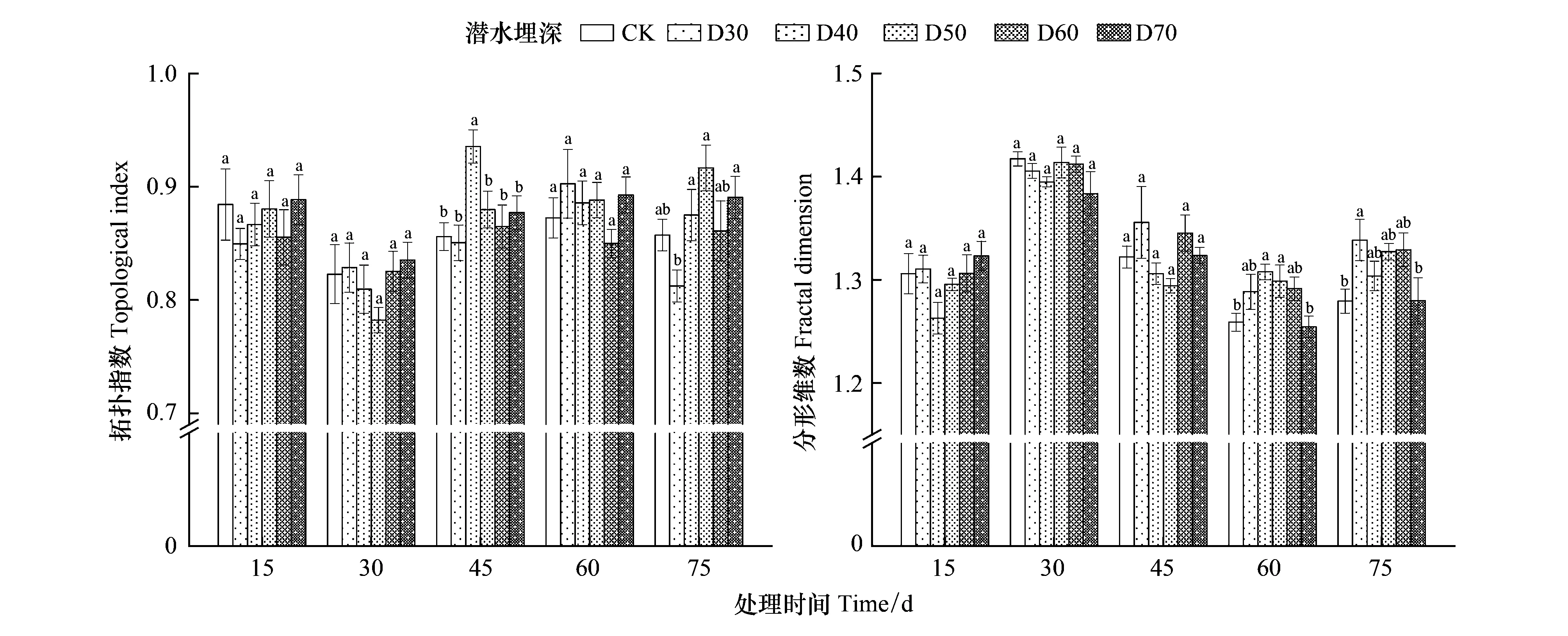
圖5 不同潛水埋深下單葉薔薇幼苗拓?fù)渲笖?shù)和分形維數(shù)的變化Fig.5 Changes in topological index and fractal dimension in Rosa persica seedlings under different groundwater levels圖中不同小寫字母表示不同組之間差異顯著(P<0.05)
2.4 不同潛水埋深對單葉薔薇幼苗表型可塑性的影響
由圖6可知,CK組單葉薔薇幼苗在總根長(0.212)、根表面積(0.333)、根體積(0.364)上可塑性指數(shù)未超過0.5,說明該組幼苗在三項指標(biāo)上可塑性相對較低;D30至D70五組幼苗在根冠比上均保持較低可塑性;D70組的株高(0.483)、D50組的總根長(0.483)可塑性指數(shù)也均低于0.5;D30組表型可塑性最為突出,在除根冠比外的9項指標(biāo)上均處于0.79—0.96之間。
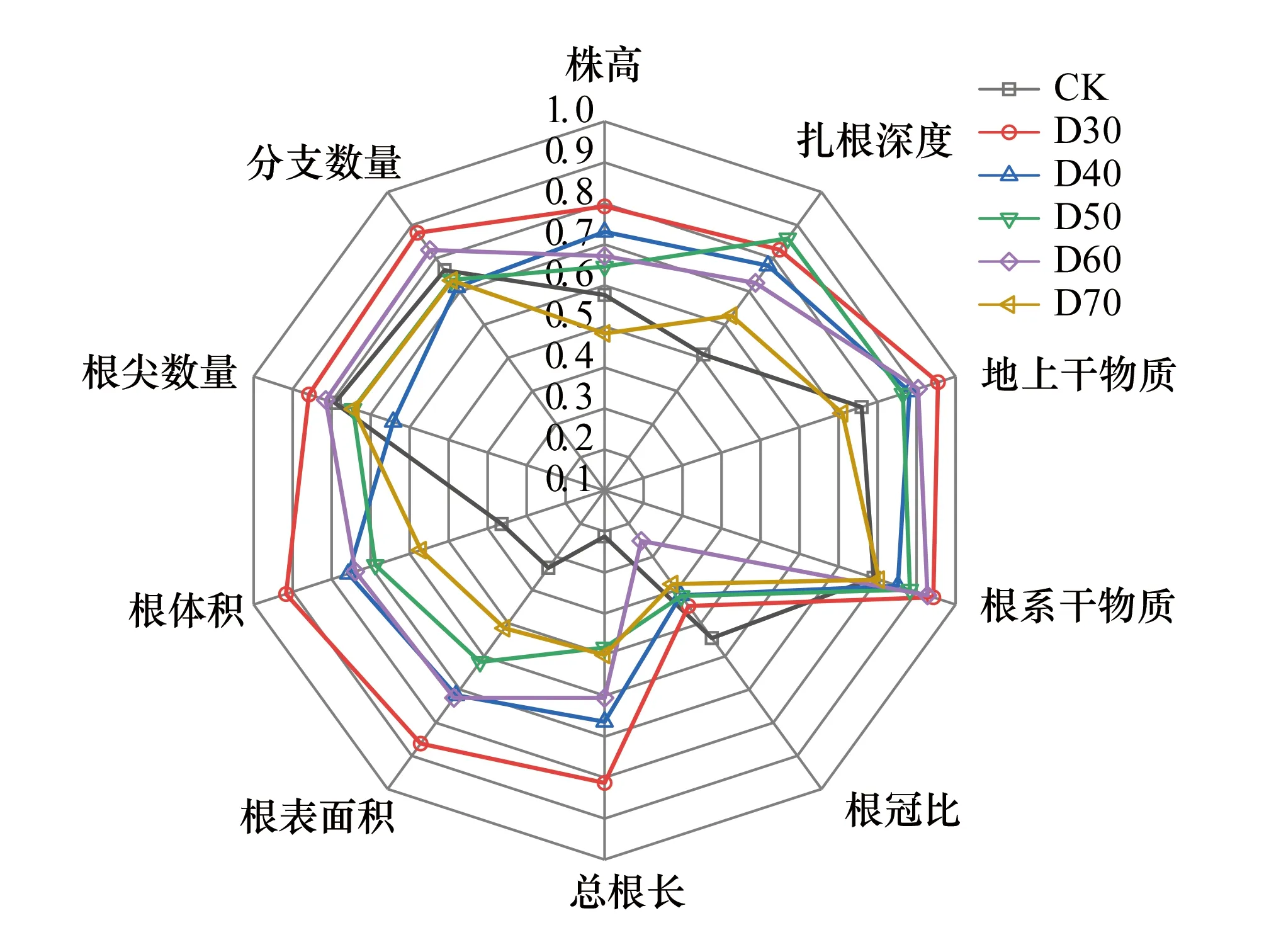
圖6 不同潛水埋深下單葉薔薇幼苗的表型可塑性 Fig.6 Phenotypic plasticity of Rosa persica seedlings under different groundwater levels
3 討論
3.1 單葉薔薇幼苗生長發(fā)育對潛水埋深的響應(yīng)
植物表型特征與水分變化有著密切關(guān)系,潛水埋深可以通過影響土壤水分進而影響根系發(fā)育與形態(tài)特征[27]。本試驗中發(fā)現(xiàn),在不同潛水埋深條件下,株高、扎根深度、根尖數(shù)量和分支數(shù)量對潛水埋深響應(yīng)具有一定規(guī)律,相比CK組根系指標(biāo)差異顯著,進一步證實水分對單葉薔薇幼苗根系影響很大。D30、D40處理下的單葉薔薇幼苗根系分別能夠在45、60 d內(nèi)到達潛水層解除干旱脅迫,進而轉(zhuǎn)向全面的生長階段,植株長勢旺盛,葉面積更大,主莖上甚至萌發(fā)出側(cè)枝(圖7),而新生側(cè)根能在水中生長,表現(xiàn)出與干旱脅迫下不同的新特點:頂部新生側(cè)根變長變直,基部根系的木質(zhì)化得以推進(圖8)。對于D30組幼苗在S5階段垂直根系生長速率為負(fù)增長的現(xiàn)象,很可能是充足的水分使得其面臨著新的脅迫,即水澇脅迫,該組容器深度10 cm處的土壤含水率已高達15.80%,所以幼苗很有可能通過降低垂直根系長度、增加分支和根尖數(shù)量來獲取更多氧氣以適應(yīng)新的環(huán)境。這點與王麗娟[25]、李強等[28]的研究發(fā)現(xiàn)一致:持續(xù)淹沒造成土壤含氧量降低,最終抑制了胡楊(Populuseuphratica)幼苗根系的生長;盛晉華等[29]采用全根挖掘法發(fā)現(xiàn),中齡與成齡的梭梭(Haloxylonammodendron)根系距地下水位50 cm處時通過毛細(xì)管和地下水相連接,土壤含水率分別為4.03%和4.59%,由于根不再下扎而是轉(zhuǎn)向水平方向延伸生長,故推測4%—5%左右土壤含水量會對梭梭根系生長產(chǎn)生明顯的抑制作用。
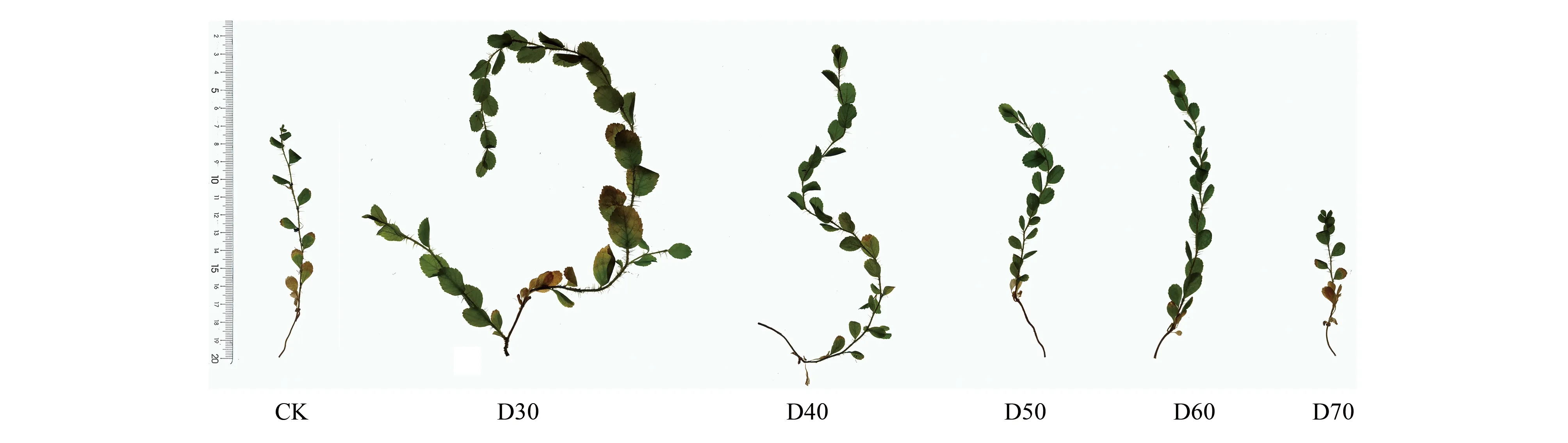
圖7 第75天時單葉薔薇幼苗地上部分對比Fig.7 Comparison of above-ground parts of Rosa persica seedlings on the 75th day
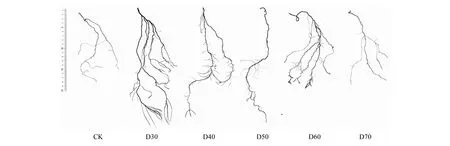
圖8 第75天時單葉薔薇幼苗根系對比Fig.8 Comparison of root of Rosa persica seedlings on the 75th day
第75天時CK、D70組幼苗在株高等18個指標(biāo)中差異均不顯著,但是二者之間又有所區(qū)別:CK組不設(shè)潛水層,所以幼苗面臨著嚴(yán)重且持久的干旱脅迫,而D70組設(shè)有潛水層,一方面土壤本身通過毛細(xì)現(xiàn)象可以向表層土運輸水分[30],另一方面幼苗增加扎根深度、根尖和分支數(shù)量,利用根尖的根毛區(qū)來吸收水分[31],所以D70組在總根長、根表面積、根體積等指標(biāo)上始終高于CK。這進一步說明適度的干旱可以促進單葉薔薇幼苗根系的生長,從而使植株表現(xiàn)出較強的抵抗能力[32—33]。
抗旱性的形成是植物形態(tài)結(jié)構(gòu)和生理生態(tài)功能對干旱環(huán)境適應(yīng)的結(jié)果[34]。當(dāng)受到干旱脅迫時,植物通常會降低生長速率來減少對水分的消耗,同時將更多的光合產(chǎn)物分配到根系以增強自身競爭能力,提高獲取土壤水分的能力,從而達到投資收益率最大化[35]。試驗組和對照組的單葉薔薇幼苗一方面調(diào)整根系形態(tài),即增加總根長、根表面積、根體積和分支數(shù)量,另一方面通過提升根系干物質(zhì)量來應(yīng)對干旱,使得根系支撐、分布范圍和單位根系所占的土壤空間不斷擴大,可以更有效地利用土壤中的水分,符合生物量分配理論。從整體來看,D50、D60、D70和CK可以通過增加根冠比應(yīng)對干旱,而D30、D40的根冠比經(jīng)歷了先升高后下降的過程,說明干旱脅迫最終得以解除,已不再需要過多地投入在根系干物質(zhì)上,進而轉(zhuǎn)向地上器官的生長,這都表明了單葉薔薇幼苗對水分脅迫響應(yīng)的敏感性和整體性,可以靈活調(diào)節(jié)生物量分配模式來更好的適應(yīng)異質(zhì)性環(huán)境。
3.2 單葉薔薇幼苗根系形態(tài)對潛水埋深的響應(yīng)
比根長和比根面積是指示根系功能的重要指標(biāo),可綜合反映植物吸收資源的能力與生態(tài)適應(yīng)性[36],值越大表明投入相同的成本會產(chǎn)生更高的養(yǎng)分利用效率[37—38],裸果木(Gymnocarposprzewalskii)[39]、楓香(Liquidambarformosana)[40]、杉木(Cunninghamialanceolata)[52]等即是通過增加比根長和比根面積來應(yīng)對干旱脅迫。但也有些研究指出:當(dāng)土壤水分缺乏時,植物根系組織密度增大,導(dǎo)致比根長、比根表面積會減小[41]。本研究中,單葉薔薇幼苗在比根長、比根面積兩個指標(biāo)上隨脅迫時間呈整體下降,根組織密度、分支密度增大,這與李金航等[26]研究結(jié)果一致,楊潔等[42]認(rèn)為這可能是試驗處理和水分梯度不同導(dǎo)致的結(jié)果,Zhou等[43]認(rèn)為這是植物在干旱脅迫下保存水分的策略之一,還有待深入探究。
粗根(d≥2 mm)被稱為結(jié)構(gòu)性根,細(xì)根(d<2 mm)又稱為功能性根,后者主要是從土壤中吸取營養(yǎng)物質(zhì)和水分,同時也是菌根形成的場所[44]。細(xì)根生理代謝極其活躍,它總是處于不斷地產(chǎn)生-生長-衰老-死亡-再生的周轉(zhuǎn)過程中,而該過程可使得植物地上與地下部分生物量得到重新分配[45],同時逆境下的細(xì)根通過主動死亡減少呼吸作用造成的能量消耗、釋放大量營養(yǎng)物質(zhì)以改良土壤,最大限度地確保植株存活生長,這是植物對環(huán)境脅迫的適應(yīng)機制[46]。本研究中的單葉薔薇幼苗根系主要由細(xì)根構(gòu)成,G1+G2+G3徑級總根長、根表面積、根體積始終占整個根系比例的80%以上,有時甚至占到100%;單葉薔薇幼苗細(xì)根的生長并非都表現(xiàn)為單一的被促進或抑制,而是與脅迫程度有關(guān),如在前60天內(nèi),G1徑級(0—0.50 mm)的細(xì)根又在整個根系中占有支配地位(69%以上),原因可能是單葉薔薇幼苗處于嚴(yán)重的干旱脅迫時,可通過增加代謝活躍的G1徑級(0—0.50 mm)細(xì)根來提高吸水能力,同時以自身的死亡完成物質(zhì)的再分配,增加根系資源利用效率,進而應(yīng)對不利的生長環(huán)境。干旱脅迫下的春玉米(Zeamays)[47]、黃櫨(Cotinuscoggygria)幼苗[48]也有類似現(xiàn)象。至第75天時,除了CK和D70外,其他組的G1徑級占比均下降,G2+G3徑級(0.50—2.00 mm)的細(xì)根總根長、根表面積、根體積占比伴隨著干旱的基本解除和根系木質(zhì)化而得以增加。
3.3 單葉薔薇幼苗拓?fù)浣Y(jié)構(gòu)和分形維數(shù)對潛水埋深的響應(yīng)
根系的空間分布決定植物對土壤的探索范圍和對土壤資源可用性進行動態(tài)響應(yīng)的能力[49—50]。基于根的連接和分支點的排列,Fitter等[51]總結(jié)出兩種根系拓?fù)浣Y(jié)構(gòu)類型,即人字形分支和二叉狀分支,并且通過拓?fù)渲笖?shù)來反映不同植物根系的分支模式。不同處理下的單葉薔薇幼苗根系拓?fù)渲笖?shù)保持在0.8—0.9之間,潛水埋深影響較小。雖然人字形分支模式根系建成成本高,但是根系內(nèi)部競爭小,是植物對營養(yǎng)匱乏生境的適應(yīng),有利于獲得更深土壤中的水養(yǎng)資源[52]。分形理論是對根系構(gòu)型定量化研究的新方法,根系分形維數(shù)越大,分支越多;相反,分形維數(shù)越小,反映出根系的分支能力相對越弱[7]。本試驗中,D30處理下的根系分形維數(shù)表現(xiàn)出先升后降再升的趨勢,在分支數(shù)量、根尖數(shù)量兩個指標(biāo)的變化中得以驗證,尤其是第60—75天時,D30組分形維數(shù)再次上升,分支能力加強,這最終也導(dǎo)致了D30的拓?fù)渲笖?shù)下降,有向叉狀分支模式(Ti=0.5)過渡的發(fā)展趨勢,未來可以延長D30等處理的觀測時間,進一步探究單葉薔薇幼苗根系在水分充足的條件下有無發(fā)生叉狀分支的可能性。雖然CK、D70組幼苗在分支數(shù)量、根尖數(shù)量上整體保持緩慢增長,但與第一次采樣相比兩組的分形維數(shù)均出現(xiàn)下降,說明后期的單葉薔薇幼苗根系分支能力相對減弱,這可能是受長期干旱導(dǎo)致的結(jié)果,其植株枝葉受脅迫表現(xiàn)更為明顯(圖7)。
3.4 單葉薔薇幼苗表型可塑性對潛水埋深的響應(yīng)
可塑性在自然界中廣泛存在,按性狀類型可分為形態(tài)可塑性、生理可塑性、行為可塑性、生活史可塑性等,是植物適應(yīng)環(huán)境變化的最直觀的策略之一[53]。表型可塑性是指同一基因型的物種應(yīng)對異質(zhì)生境而改變表型性狀的能力[54]。表型可塑性強的植物往往具有更強的適應(yīng)性和更寬的生態(tài)幅,占據(jù)更大范圍的生境,從而成為廣幅種[55]。楊寧等[56]研究發(fā)現(xiàn),林下、中林窗、大林窗等4種不同林冠環(huán)境中,林緣曠地的箭竹(Sinarundinariabasihursuta)地下莖、粗根、細(xì)根生物量百分比均最高,分株形態(tài)差異顯著;葉子奇[57]對干旱區(qū)河漫灘胡楊根系生長與形態(tài)適應(yīng)對策進行了研究,發(fā)現(xiàn)當(dāng)年生胡楊幼苗根系構(gòu)件(主根、側(cè)根、末端根)即可對干旱脅迫表現(xiàn)出不同程度的響應(yīng),尤其是在重度干旱脅迫下,幼苗根冠比和根構(gòu)型變化相當(dāng)顯著。面對70 cm潛水埋深和對照組帶來的長久干旱時,單葉薔薇幼苗僅在地上干物質(zhì)、根系干物質(zhì)、根尖數(shù)量和分支數(shù)量4個指標(biāo)上有較大的可塑性(0.70—0.80);相反,最強的表型可塑性發(fā)生在D30組株高、扎根深度、地上干物質(zhì)、根系干物質(zhì)、總根長、根表面積、根體積、根尖數(shù)量、分支數(shù)量9個指標(biāo)上(0.793—0.955),這說明單葉薔薇幼苗根系在第一年內(nèi)就可發(fā)生功能分化,形態(tài)可塑性、分配可塑性極為顯著。占據(jù)多樣的生境或最大限度地達到表型與生存環(huán)境的一致性,可以有效緩沖環(huán)境對植物生長和生殖產(chǎn)生的消極影響,從而可以避免成為生態(tài)角度上的脆弱小種群,避免造成遺傳瓶頸[58]。
4 結(jié)論
潛水埋深可以顯著影響單葉薔薇幼苗的生長發(fā)育,幼苗可通過改變根系形態(tài)、增加扎根深度、提高根系生物量等根系構(gòu)型特征來應(yīng)對中短期的干旱脅迫,深層潛水和CK處理下的幼苗還通過擴大根冠比來適應(yīng)更長久的干旱環(huán)境。從資源分配的角度來看,單葉薔薇幼苗為減少碳消耗而采用相對簡單化、內(nèi)部競爭較小的人字形分支結(jié)構(gòu),短而細(xì)的密集橫向分支(0—2 mm細(xì)根為主)成為根系擴大土壤資源獲取效率的活性位點,從而保障植株生長發(fā)育的物質(zhì)供需平衡,這是單葉薔薇對環(huán)境脅迫的適應(yīng)性策略。適度的干旱,如50—70 cm的潛水埋深,可以促進單葉薔薇幼苗扎根深度,而單葉薔薇幼苗在較淺的潛水埋深(30—40 cm)環(huán)境中能快速解除干旱,更好的進行地上器官的生長發(fā)育,具體表現(xiàn)為在在株高、總根長、根表面積、根體積、根尖數(shù)量、分支數(shù)量、根組織密度、分形維數(shù)和地上、地下干物質(zhì)10個指標(biāo)上與CK組差異顯著,這表明單葉薔薇幼苗對異質(zhì)性環(huán)境具有極高的適應(yīng)能力,表型可塑性強。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
