杭州午潮山天然次生林木本幼苗組成及其時空分布格局
2022-08-31 06:08:30岳春雷李賀鵬
生態學報 2022年15期
關鍵詞:物種
朱 弘,楊 樂,岳春雷,李賀鵬
浙江省林業科學研究院,杭州 310023
森林的更新和演替是一個復雜的生態學過程,也是森林生態領域長期關注的熱點問題之一[1]。其中,木本植物實生幼苗天然更新是森林植被恢復與演替的主要方式,木本幼苗通過定植、生長、擴散,對未來森林群落的物種構成、種群結構、分布格局與演替進程帶來深遠的影響[2]。森林更新受樹種生理特性、地理環境、自然干擾、人為干擾等因素及其相互作用的影響,這些生物的和非生物的因素隨時間和空間而不斷變化,構成了森林的空間異質性和時間異質性,使森林更新具有時空變化的特點[3]。
杭州午潮山開發利用歷史悠久,近代由于過度森林砍伐和不合理的土地利用活動,導致區域內原始植被遭到了嚴重破壞;1960年后,浙江省林業科學研究院將其作為科研基地加以保護,經過長期的封山育林,演替形成了以木荷(Schimasuperba)、青岡(Cyclobalanopsisglauca)、苦櫧(Castanopsissclerophylla)等為建群種的浙北典型天然次生林[4]。近年來,有關午潮山森林的群落結構、物種組成、功能性狀等方面已經取得了一定的進展[5—6],然而相關研究主要針對喬木大樹,對其林下幼苗的關注度明顯不足,特別是在大樣地的實踐中,往往被選擇性忽視,從而限制了對該區域森林生態系統的全面認識。
幼苗階段被認為是森林演替過程中最重要和最敏感的階段,其種類構成、數量和存活動態等方面的變化也最為明顯[7]。基于幼苗層物種的空間分布規律和自然更新的探究,有助于預測森林演替方向和進程[8—10]。鑒于此,本研究重點以杭州午潮山天然次生林天然更新幼苗為研究對象,關注以下3方面問題:1)林下木本幼苗物種組成和數量特征;2)主要優勢樹種幼苗基徑結構;3)優勢木本幼苗在不同發育階段的空間分布規律和小尺度空間效應。研究結果旨在為為豐富北亞熱帶森林物種多樣性構建與群落演替機制資料,也為加快該區域的植被恢復重建和森林經營管理提供依據。
1 材料與方法
1.1 研究區概況
研究區地處杭州西郊,屬天目山余脈,平均海拔264 m,主峰504 m。氣候帶屬亞熱帶季風氣候,其中年平均氣溫15.4℃,極端最高溫39℃,極端最低溫-9.4℃,年均降水量為1 992.5 mm,年無霜期240 d,年平均相對濕度81%;森林植被類型涵蓋常綠闊葉林、落葉闊葉林、針闊混交林、針葉林、竹葉林和灌木林6大類型,森林覆蓋率達98.2%。土壤類型為紅壤[6,11]。
1.2 樣地設置和幼苗調查
2021年3—4月,在前期林分踏查的基礎上,從幼苗分布較為豐富的典型溝谷地(30.19°N,120.00°E,海拔237 m)設置一個與底部溪流方向大致平行的矩形樣帶,再在樣帶內采用相鄰格子法劃分出6個5 m × 5 m的小樣方,調查面積共計150 m2。以樣方左下角為坐標原點建立直角坐標系,調查和記錄樣方內胸徑(Diameter at Breast Height,DBH)<1 cm的林下所有幼苗的物種名稱和空間二維坐標(x,y),并利用卷尺和數顯游標卡尺(桂林廣陸數字測控有限公司)測量每個個體的株高和基徑(Stem base diameter, SBD)。
1.3 數據處理
1.3.1幼苗數量特征
幼苗密度=幼苗總株數/樣方面積(m2)
(1)
幼苗豐富度=物種數/樣方面積(m2)
(2)
幼苗重要值(IV)=(相對多度+相對頻度)/2
(3)
式中,相對多度(RA)=(某個種的株數/所有幼苗總株數) × 100%,頻度(F)=某物種出現的樣地數/樣地總數× 100%,相對頻度(RF)=(某一物種的頻度/所有物種的頻度之和) × 100%[12—13]
1.3.2優勢木本幼苗的確定和不同徑級結構的劃分
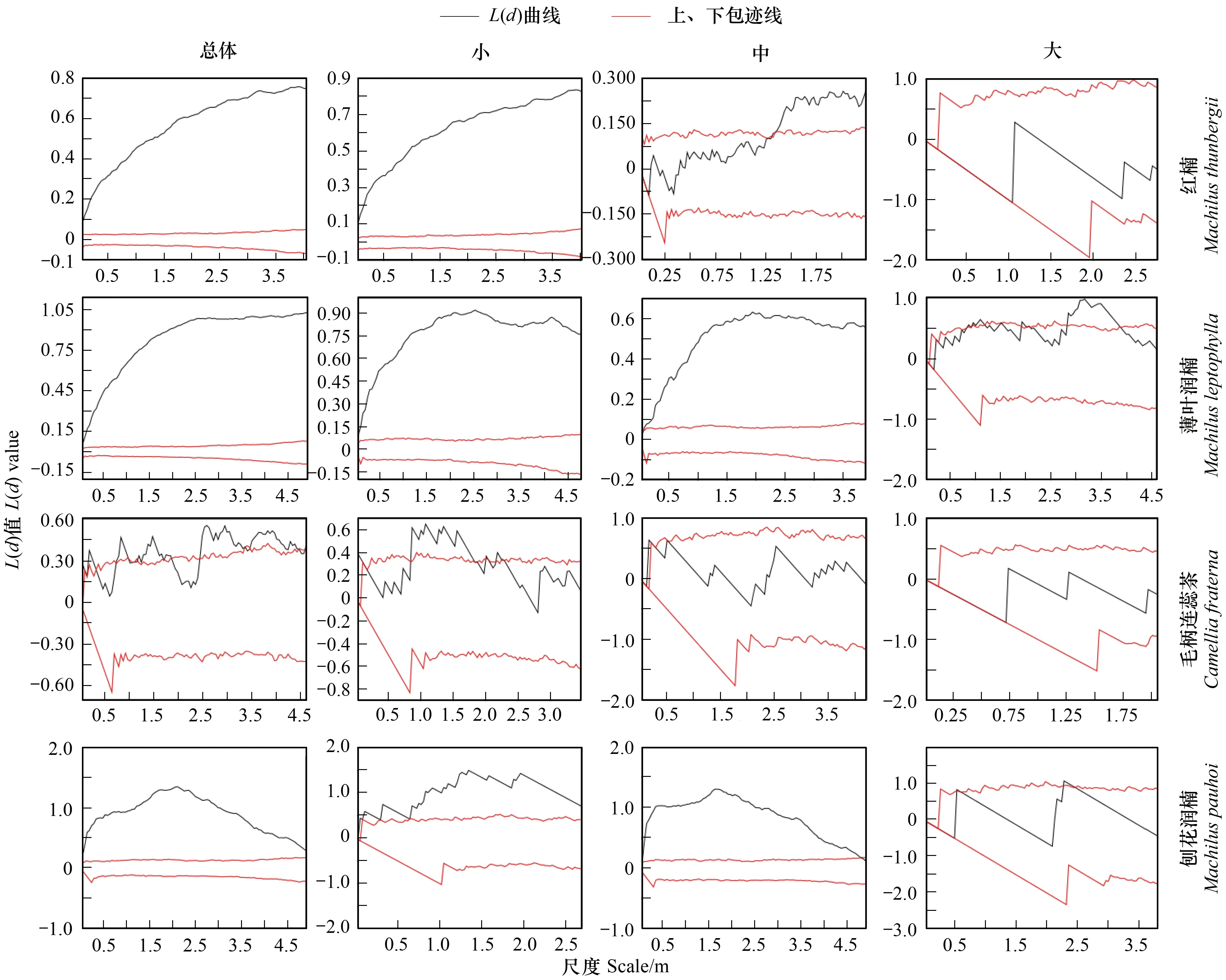
按重要值大小進行排序(表1),篩選IV≥0.3的物種作為優勢木本幼苗作為后續研究對象。依據“空間替代年齡”的原理和方法,即將基徑從小到大的順序視為時間順序關系,首先繪基徑制頻率直方圖(Frequency histogram),將頻率分布較大轉折點作為齡級劃分的標準[14],同時結合樣地內幼苗個體數及自身特性的實際情況,最終將不同優勢樹種基徑(BD)劃分為小徑級(Small-size classes)、中徑級(Medium-size classes)和大徑級(Large-size classes)3個徑級,視為不同生長發育階段。其中紅楠(Machilusthunbergii)3個徑級依次為:1.32 mm≤BD≤5.55 mm、5.55 mm 1.3.3最近鄰分析 利用最近鄰分析(Nearest Neighbor Analysis, NNA)法分析不同物種在不同徑級下的空間分布格局。它最早于1954年由密歇根大學的Clark和Evans提出[15],是一種經典的點格局分析方法。其原理是將隨機分布模式下的預期最近鄰距離與實際觀測得到的最近點的平均距離進行比值,用得到的最近鄰指數(Nearest neighbor index, NNI)來作出種群的分布類型[14]。公式如下: (4) 式中,A是研究區的總面積,di是i第棵樹與其最近樹之間的距離,n為每類樹的總株數。當NNI接近1時,呈隨機分布;NNI<1時,呈聚集分布;NNI>1時,呈離散分布。為了更好地反映實測平均距離和預期平均距離的偏離程度,利用正態分布檢驗得出Z值及其95%置信水平。Z值為負且值越小,個體越趨于聚集分布;Z值正值且越大則顯著偏向均勻分布;Z值位于二者之間則為隨機分布。 1.3.4Ripley′s K函數分析 由于種群空間格局特征具有很強的尺度依賴性,因此進一步利用Ripley′sK函數對不同物種在不同距離尺度下的空間分布格局進一步分析。該方法自1977年由Ripley[16]首次提出,已經在國內外廣泛使用。公式如下: (5) 式中,d為空間尺度,A為研究樣地面積,n為樣地內某一種植物的個體總數,uij為個體i到個體j之間的距離,Wij為邊界效應校正系數。當uij≤d時,Ir(uij)=1;當uij>d時,Ir(uij)=0;為保持方差穩定,對K(d)作開平方的線性變換如下: (6) 在隨機分布下,L(d)在所有空間尺度下均等于0;當L(d)>0,表示幼苗個體服從聚集分布;當L(d)<0,表示幼苗個體服從均勻分布。 將實測格局與選定的零模型(null models)通過Monte-Carlo法繪制空間格局,擬合上、下兩條包跡線,重復驗證99次,確定95%的置信水平。用實際數據計算不同尺度的L(d)值,若L(d)在包跡線以內,則呈隨機分布;若L(d)值在包跡線以上,則呈顯著聚集分布;若L(d)在包跡線以下,則呈顯著均勻分布;上述兩種點格局分析法的運行和出圖均在PAST version 4.03統計軟件包(http://folk.uio.no/ohammer/past/)中實現。 研究樣地內共記錄到811株實生幼苗,隸屬12科20屬27種(表1),其中樟科(Lauraceae)(5)、殼斗科(Fagaceae)(4)、山茶科(Theaceae)(3)和山礬科(Symplocaceae)(3)為排名前4的科;前4個優勢種(IV≥0.3)為紅楠、薄葉潤楠、毛柄連蕊茶、刨花潤楠累計個體總數為691株,占樣地內所有幼苗總株數的85.20%;優勢種各級散布模式和聚集程度明顯不同(圖1);樣地總體幼苗密度和幼苗豐富度分別為5.4株/m2和0.18種/m2。 表1 午潮山林下幼苗物種組成及數量特征 圖1 午潮山150 m2樣地內4個優勢木本幼苗不同基徑結構的空間分布圖Fig.1 Spatial distribution of four dominant tree seedlings at different growth stages in Wuchao Mountain with 150 m2 plot 從主要優勢木本幼苗基徑分布的小提琴圖(圖2),可以看到不同樹種基徑分布有一定的差異:紅楠和薄葉潤楠總體基徑呈典型的正偏態分布,差別在于紅楠小徑級個體數量優勢明顯占其總株數的83.44%,預示著其種群結構屬于快速增長型;而薄葉潤楠小徑級與中徑級的比例接近1∶1,兩端存在更多的離散值,種群結構屬于穩定增長型;毛柄連蕊茶總體呈現略微正偏分布的雙峰型,各徑級數量分布較為分散,小基徑占其總株數的51.72%,且起始值較大,種群結構屬于緩慢增長型;刨花潤楠總體基徑近似正態分布,中徑級占其總株數的77.02%,種群結構屬于穩定型。 圖2 午潮山林下代表性木本幼苗的基徑分布的小提琴圖 Fig.2 Violin plots of stem base diameter (SBD) classes distribution of understory seedling for typical tree species in Wuchao Mountain 利用最近鄰分析分別對4個優勢種3個徑級的空間分布格局進行空間分布格局的判定,結果(表2)表明:紅楠總體和小徑級的NNI指數<1,Z值均為負值,在空間上呈顯著聚集分布;中徑級和大徑級的NNI指數>1,Z值均為正值,在空間上呈顯著離散分布。薄葉潤楠總體和小徑級的NNI指數<1,Z值均為負值,在空間上呈顯著聚集分布;中徑級的NNI指數接近1,Z值為負值,未能拒絕在P<0.05的水平上隨機零假設,在空間上呈不顯著隨機分布;大徑級NNI指數>1,Z值均為正值,在空間上呈顯著離散分布。毛柄連蕊茶總體的NNI指數接近1,Z值為正值,未能拒絕在P<0.05的水平上隨機零假設,在空間上呈不顯著隨機分布;小、中、和大徑級的NNI指數均>1,Z值均為正值,在空間上呈顯著離散分布。刨花潤楠總體、小徑級和中徑級的NNI指數均<1,Z值均為負值,在空間上呈顯著聚集分布;大徑級的NNI指數>1,Z值均為正值,在空間上呈顯著離散分布。 表2 午潮山4個優勢木本幼苗在不同生長階段的最近鄰分析結果 Ripley′sK函數分析結果(圖3)顯示,紅楠總體和小徑級在0—4 m范圍內均呈聚集分布;中徑級在0—1.25 m范圍內呈隨機分布,在>1.25 m呈聚集分布;大徑級在0—2.5 m范圍內均呈隨機分布。薄葉潤楠總體和小徑級在0—4.9 m范圍內均呈現強烈聚集分布;中徑級在0—3.9 m范圍內呈隨機分布;大徑級在0—2.8 m和3.8—4.5 m范圍內呈隨機分布,在2.8—3.8 m范圍內呈聚集分布。毛柄連蕊茶總體在0—4.5 m范圍內呈集中和隨機的波動模式;小徑級在0.8—2.2 m范圍內呈集中分布,伴有少量隨機分布;中徑級和大徑級分布在0—4 m和0—2 m范圍呈隨機分布。刨花潤楠總體和中徑級在0—5 m范圍內呈集中分布,當d=2 m左右,L(d)=1.23,聚集強度分別達到最大;小徑級在0—2.7 m范圍內呈集中分布;大徑級基本在0—3.9 m范圍內呈隨機分布。 圖3 午潮山樣地內4個優勢木本幼苗不同徑發育階段的Ripley′s K函數分析Fig.3 The Ripley′s K analysis of four dominant tree seedlings at different growth stages in Wuchao Mountain 研究森林群落組成、結構隨演替的變化規律,對于揭示森林群落構建與物種共存機制具有重要意義[17—20]。基于小尺度的樣方調查表明,午潮山喬木幼苗樹種組成較為豐富,群落整體分布密度較高,表明林下自然更新良好。但不同種間的個體數量分配差異較大,表明不同種的繁殖能力與環境適應是不同的。徑級結構在一定程度上反映植物種群的年齡結構特征和生長發育狀況,成為預測種群和群落的動態發展趨勢的一種有效方法。在所有調查的木本幼苗中,紅楠和薄葉潤楠是重要值排序前兩位的樹種,也是中至北亞熱帶的優勢種[21],由于儲備較多的小徑級幼苗個體,屬于增長型種群,或將成為下一階段午潮山群落演替的建群樹種。毛柄連蕊茶是亞熱帶山區常見伴生種,通常以常綠灌木或小喬木的形式生長,也是早春重要的蜜源植物之一[22]。依靠其顯著的基徑和高度,重要值排名維持至第3,雖然個體數量不多,但生長良好,推測未來仍將作為林下伴生樹種與其它物種長期混交共存。刨花潤楠的重要值排名第4,該種是亞熱帶重要的山地鄉土樹種,亦是集經濟用材價值和園林觀賞價值為一體的珍貴特色樹種[23],其幼苗各徑級呈近正態分布的樣式反映出該種生長發育對本區域有著很好的適應性。總之,午潮山木本幼苗是午潮山森林資源的重要組成部分,上述優勢物種通過占據不同水平與垂直空間,采取不同的生活史對策,在群落中得以共存,并維持群落的穩定性。 種群分布格局的變化反映了植物生存策略、適應機制,進而影響群落的發展趨勢[24—27]。利用最近鄰分析可以分析全局尺度下樹種的空間分布格局,而基于Ripley′sK函數分析能夠進一步量化研究區域內植物個體在不同空間尺度上的分布格局,從而揭示植物種群的生態學特性和形成機制[28]。在本研究中,紅楠、薄葉潤楠和刨花潤楠3個潤楠屬在整體和小徑級個體均呈聚集分布,隨著徑級和空間尺度的增大,聚集強度達到最強后逐漸降低,常呈隨機或均勻分布。說明不同樹種在不同發育階段、不同齡級也會形成不同的空間格局,這通常是自身生物學特性和環境因素共同作用的結果。可能的原因如下:首先,3種植物均隸屬于樟科潤楠屬植物,幼苗期耐陰蔽,喜濕潤肥沃土壤,研究地處低海拔溝谷,水熱充沛,較厚的林下凋落物層有利于種子冬季儲存及打破生理休眠,從而獲得較高的種子萌發率,促進幼苗的集中大量萌發和生長;其次,核果球形,推測擴散傳播以重力傳播和水流傳播為主,因而野外觀察到大部分種子集中散落在母樹周圍及沿河流溝谷兩側;此外,3種植物低齡級幼苗個體較小,處在森林群落的下層,對光照、水分、土壤等環境因子的競爭能力較弱,聚集分布的幼苗在母樹遮蔽下可以形成幼苗群,能夠形成有利于其生長發育的微環境,從而提高個體競爭能力及成活率。但是當種群年齡和空間尺度逐漸增大,種子有效傳播能力和繁殖能力不斷減弱,種內種間對資源和空間的需求不斷激增,加上病蟲害、草食動物取食用等侵擾加劇帶來的負密度制約效應影響[29],導致大量個體死亡,種群密度下降,聚集強度隨之減弱,種群由聚集分布向隨機分布或離散分布轉變[30]。此外,還可以看到,紅楠與薄葉潤楠的分布格局較為接近,而與刨花潤楠相差較大,推測前兩者具有更相似的生物學特征,如均耐陰喜濕、更大的比葉面積、幼枝輪生等特征,相似的生態位導致生境選擇的趨同,從而導致兩者常常相伴而生。 相較而言,毛柄連蕊茶的個體數量較少、種群密度較低,分布格局由整體上的不顯著隨機分布向顯著離散分布轉變,這一現象的產生,一方面,受限于自身較慢的生長速率,導致其在幼苗層群落中的競爭力有所弱化,林中大量草本和藤本生長占據地表空間,使得許多的毛柄連蕊茶小徑級幼苗容易被掩蓋,影響生長發育。另一方面,由于常年處于林冠下層,郁閉度較高,在授粉期光照不足情況下,毛柄連蕊茶植株的開花結實率有下降的可能。此外,所在生境還頻繁受到人為干預,如當地林場工人的砍伐、挖筍、采茶等活動導致小苗很難正常生長,最終影響了該物種種群結構的正常演替。 綜合上述研究結果,建議在森林經營過程中:1)可對小尺度范圍內集中分布和長勢不良(如病蟲害或者頂芽受損)的紅楠和薄葉潤楠幼苗進行合理間伐,降低小徑級階段自疏的比例,提高中、高徑級的比例。2)對林下郁閉度較高、枯死木、病株、攀援藤本(如南五味子Kadsuralongipedunculata、雞矢藤Paederiascandens等)所在區域進行及時清理,有利于林窗的快速形成,從而提高林下木本幼苗(如毛柄連蕊茶)的更新率和成活率。3)對殼斗科、山茶科等亞熱帶常綠闊葉樹種(如表2中重要值排序稍靠后的青岡、苦櫧、木荷等樹種)幼苗進行適當撫育或補植工作,優化種群結構,加速群落向頂級群落的演替過程,最終促進整體森林群落結構的正向更新,提高群落的穩定性與物種多樣性。2 結果與分析
2.1 林下幼苗組成與數量特征
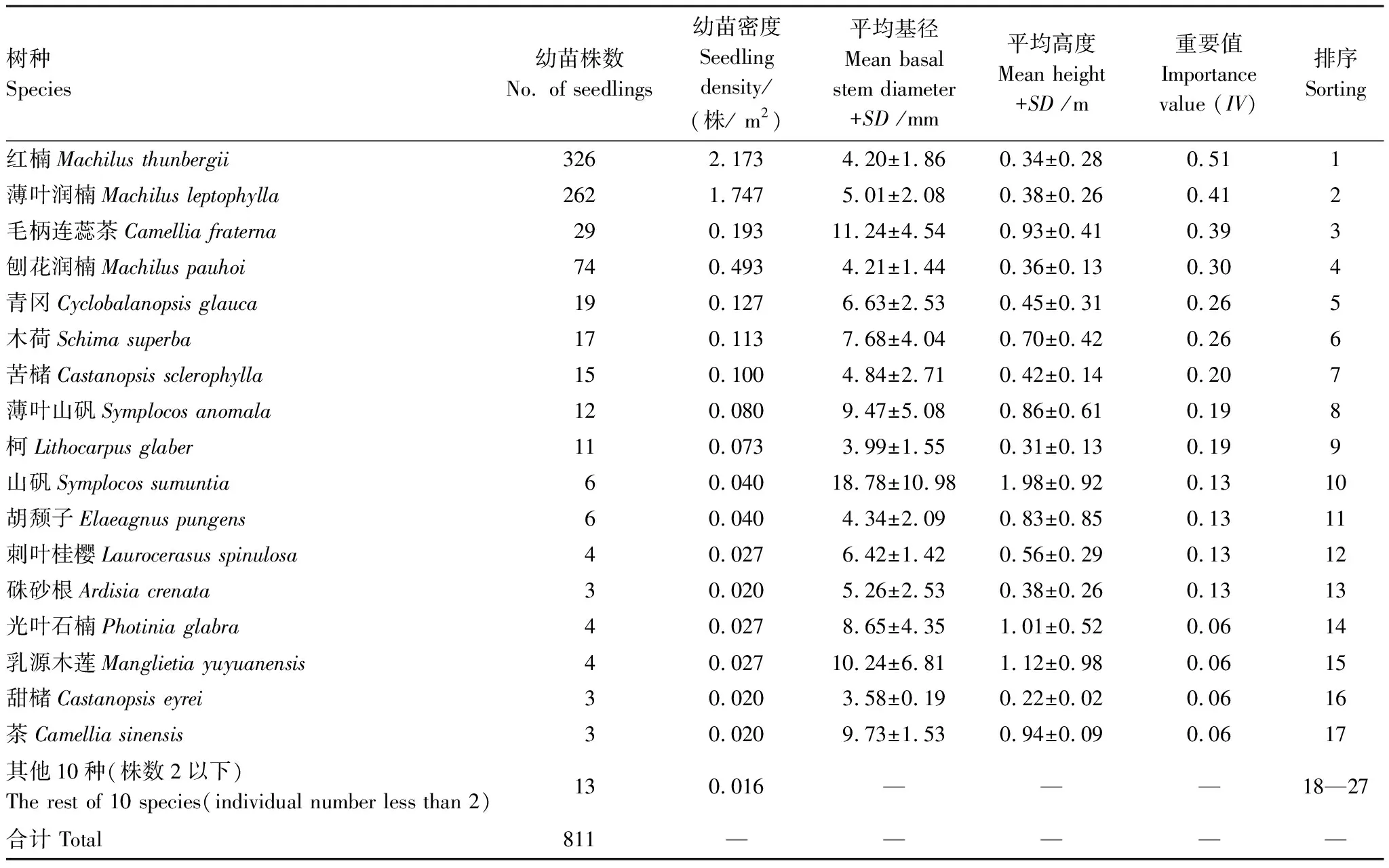
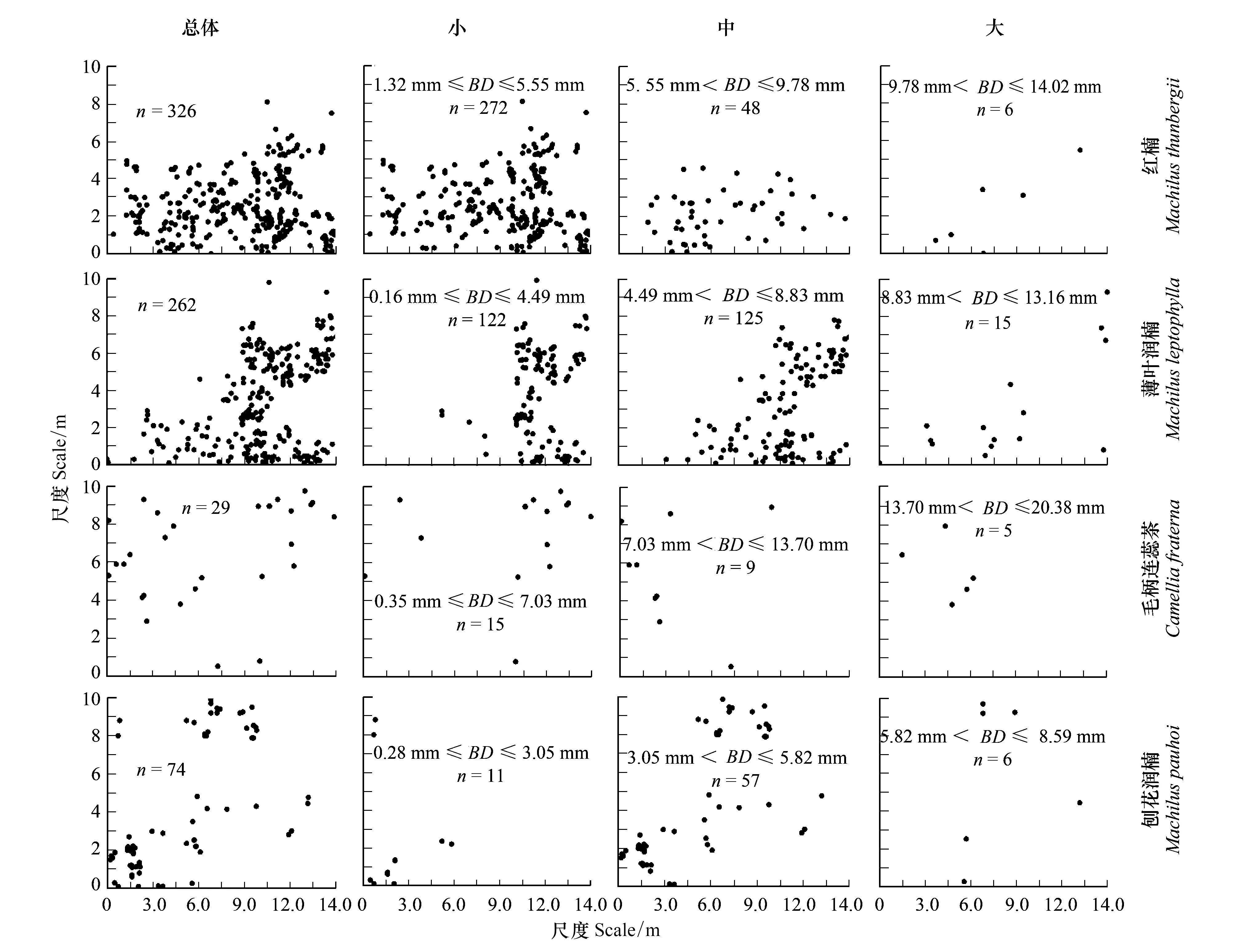
2.2 優勢樹種幼苗的基徑結構
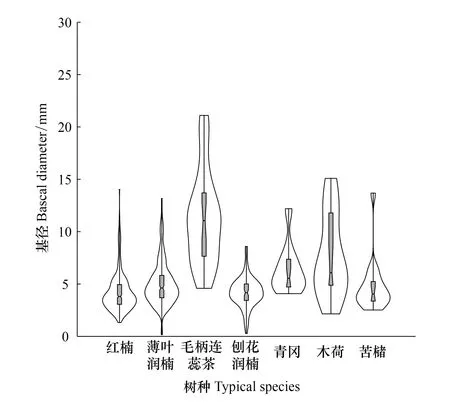
2.3 優勢種幼苗不同徑級個體的空間分布格局
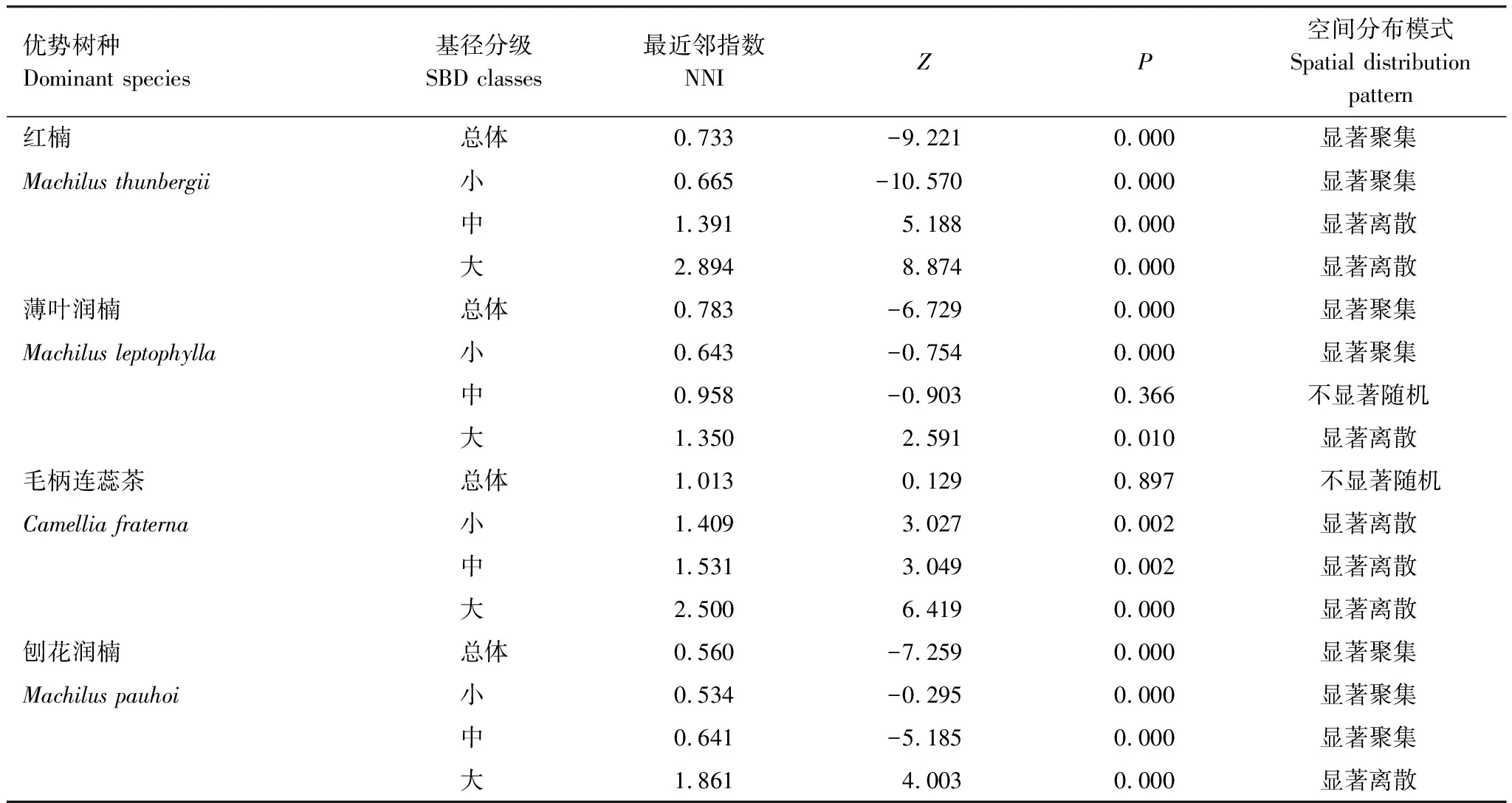
2.4 優勢種幼苗不同徑級個體多尺度特征分析
3 討論
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
