哀牢山常綠闊葉林常見兼性附生植物的化學計量特征
2022-08-31 06:02:34張婷婷劉文耀
生態學報 2022年15期
關鍵詞:植物
張婷婷,劉文耀,胡 濤
1 中國科學院西雙版納熱帶植物園熱帶森林生態學重點實驗室,昆明 650223 2 中國科學院大學,北京 100049
附生植物是山地森林生態系統中重要的結構性組分之一,在維系生態系統生物多樣性、生態功能及其過程發揮著重要的作用[1—2],兼性附生植物是一類重要而特殊的植物類群,它能同時在樹皮、巖石表面和林地土壤中生長,是生長于不同基質上的同一物種的不同生態型[3]。同種植物不同生態型分布方式的差異可能反映了植物對環境變化的特殊適應策略,導致其生態化學計量特征與養分利用策略有所不同[3]。研究生態過程中生物與環境間能量以及多種化學元素交互作用是生態化學計量學的主要研究內容[4]。碳(C)、氮(N)、磷(P)是植物生長發育所需的主要元素,也是生物體有機體的組成基礎。C、N和P這3種元素相互作用,共同調節植物的生長發育[4—5],其元素比值可以用來指示植物的養分利用率以及養分限制狀況[5—6]。植物葉片的N∶P比通常用來判斷植物的養分受限情況,具有較高的N∶P比的植物易受P限制,反之則受N限制[5, 7]。植物葉片中C∶N與C∶P可在一定程度上反映其養分利用效率,當C∶N和C∶P比值較高時,表明植物的養分利用效率較高,反之則較低[6, 8]。生態化學計內穩性表征生物有機體在變化的環境中維持其自身元素組成相對穩定的能力[4, 9],其強弱與生物有機體的生態策略和適應性有關[10]。有關研究表明:植物通過調節不同器官間的化學計量內穩性來適應環境的變化[10]。在內蒙古草原,不同植物葉片和根之間的內穩性呈負相關關系,其中羊草(Leymuschinensis)葉片內穩性指數最大,而其根中內穩性為最弱,灰綠藜(Chenopodiumglaucum)葉片內穩性最弱,根中內穩性最強[10]。Schreeg等認為,灌木植物的新葉內穩性較強,而莖和根等對環境變化更敏感,即其內穩性較弱[11]。Wu等人研究發現,在西雙版納地區不同生境下蘭科植物葉片表現出較強的化學計量可塑性,以此來應對生境轉變[3]。目前對于森林生態系統中兼性附生植物的化學計量特征,以及它們如何通過調節其化學計量內穩性來應對生境變化的了解甚少。
地處云貴高原、橫斷山和青藏高原南緣三大地理區域結合部的哀牢山國家級保護區, 是生物多樣性較豐富及植物區系地理成分薈萃之地,該保護區內分布著我國目前保存最完整、面積最大的亞熱帶山地濕性常綠闊葉林,林內附生植物非常豐富[12],其中長葉粗筒苣苔(Briggsialongifolia)是哀牢山山地森林中一種常見的兼性附生植物[13],它分布于樹干、樹根際部及林地土壤不同基質中,表現出獨特的適應能力。但目前關于兼性附生植物這一特殊植物類群生態化學計量特征與養分利用策略鮮有研究報道。長葉粗筒苣苔屬于苦苣苔科(Gesneriaceae)物種,該科植物主要分布于亞洲和非洲熱帶及亞熱帶地區,我國苦苣苔科植物的生物多樣性十分豐富,其中滇黔桂及其鄰近地區是我國苦苣苔亞科植物分布和分化的中心,苦苣苔科很多種類是藥用植物,一些種類具有較高的園藝觀賞價值,有相當一部分為瀕危植物[14]。苦苣苔科植物除少數類群適應力比較強外,絕大多數是完全依賴于特殊的優良小生境來維持生存[15],且同種不同居群間存在形態變異[16]。因此,苦苣苔科植物具有很高的研究價值[15, 17]。本研究以哀牢山山地濕性常綠闊葉林中常見的兼性附生植物長葉粗筒苣苔為對象,通過研究其地生植株與林冠不同高度上附生植株不同器官化學計量的差異,分析兼性附生植物化學計量的垂直空間分布特征及其與生長基質的關系,探討不同生境下兼性附生植物的養分利用與生態適應策略,為深入研究山地森林生態系統生物多樣性與養分循環,以及山地森林資源的保護與管理提供科學依據。
1 材料與方法
1.1 研究區概況
哀牢山位于云南省中部,為云嶺向南的延伸,是云貴高原和橫斷山脈的分界線。該地區受到西南季風的影響,具有干、濕季分明的特點。哀牢山國家自然保護區至今分布著我國目前保存最完整、面積最大的亞熱帶原生山地濕性常綠闊葉林[13, 18]。該研究位于其核心區徐家壩地區(23.35°—24.44°N, 100.54°—101.30°E),根據中國科學院哀牢山生態站長期氣象觀測結果顯示,區年均降水量1947 mm, 年均蒸發量1192 mm, 干季(11月—次年4月)、雨季(5月—10月)分明, 雨季降水量約占全年降水量的85%;年平均相對濕度85%, 年平均氣溫11.3 ℃,月平均最高氣溫為15.8 ℃, 月平均最低氣溫為4.5 ℃[19—20]。
徐家壩地區山地濕性常綠闊葉林分布于海拔 2400—2600 m 間,其群落上層的優勢喬木主要包括木果柯(Lithocarpusxylocarpus)、變色錐(Castanopsiswattii)等[18]。該區森林附生植物物種豐富,根據前期的調查結果,附生地衣有183種,附生苔蘚植物176種,附生蕨類117種和附生種子植物113種[21]。兼性附生植物是一類重要而特殊的附生植物,其中長葉粗筒苣苔(Briggsialongifolia)是該區森林中常見的兼性附生種子植物[13],它是苦苣苔科粗筒苣苔屬多年生無莖草本,多附生于木果柯(L.xylocarpus)、變色錐(C.wattii)等高大的喬木樹干及樹木根際處,也有部分植株生長在林地土壤中,具有明顯的垂直分布特征,是研究同一物種在不同生境下化學計量特征與養分利用策略的理想材料。
1.2 樣品采集
已有的研究表明,林冠腐殖質與林地土壤的養分條件差異較大,且林冠層與林地的小氣候也存在一定差異[22]。而環境因子的變化,尤其是土壤中養分的變化會引起植物各組織內元素組成的改變[23]。其次,高大喬木的根部,即附生、地生生境交界處的土壤養分等環境因子可能與林地以及林冠生境存在一定差異,因此,為了研究同一物種在不同生境下的化學計量特征與養分利用策略,依據長葉粗筒苣生長及其空間分布特性,我們對林地、宿主喬木的根部、樹干等不同生境下的植株進行調查采樣。
2018年4月對哀牢山徐家壩地區山地濕性常綠闊葉林內長葉粗筒苣苔的分布進行調查和采樣。在生態站通往愛山亭、蟾蜍洼以及馬鹿壩的道路旁,因林冠較為開闊,長葉粗筒苣苔能同時生長在喬木樹干及根部、以及宿主根部附近林地土壤中。在附生植物研究中,單株樹木通常被描述為附生植物的最小生境單元[24]。因此,本研究將林內樹干、樹干基部以及樹根周圍林地土壤均生長有長葉粗筒苣苔的單株宿主喬木作為一個樣樹。本研究在上述區域設置20 m × 2000 m的樣帶進行樣品采集,選擇7株胸徑>30 cm、附生有長葉粗筒苣苔的優勢宿主喬木木果柯、變色錐作為附生植株調查與樣品采集的宿主樣樹,共有7個重復。對每株樣樹根部周圍的地生植株,以及樹干基部和1.3 m樹高處的附生植株通過整株收獲法進行采樣,同時也采集3種生境下長葉粗筒苣苔生長基質(林地土壤、樹干基部土和樹干1.3 m處樹皮附著的腐殖質)的樣品。將各植株的葉片、根系和生長基質樣品分別裝入樣品袋,帶回實驗室。而后對所收集的植物樣品用自來水和蒸餾水沖洗,裝入信封進行烘干處理。
1.3 樣品處理及測定
將野外采集的植物及基質樣品,置于烘箱內在65 ℃條件下烘干48 h至恒重。烘干后的生長基質放入研缽研磨,過60目篩,裝入自封袋;烘干的植物樣品用植物粉碎機粉碎至粉末狀態,裝袋編號以待進一步分析。處理好的生長基質和植物樣品C、N含量用碳氮分析儀(Vario MAX CN, Elementar Analysensysteme GmbH, Hanau, Germany)測定;P含量采用電感耦合等離子體原子發射光譜儀進行測定(iCAP6300, Thermo Fisher Scientific, Waltham, USA)。
1.4 數據處理
數據主要通過SPSS 19.0進行統計分析,插圖通過SigmaPlot 12.5繪制。數據分析前均進行正態性和方差齊性檢驗。不同生境下的生長基質以及植物的化學計量組成、不同生境下植物的化學計量內穩性的差異,以及植物不同元素的內穩性差異的比較均采用單因素方差分析法(one-way ANOVA)和最小顯著差異法(LSD) (α=0.05)。用成對t檢驗比較各生境中長葉粗筒苣苔不同器官間化學計量組成以及化學計量內穩性的差異,用雙因素方差分析(two-way ANOVA)分析生境、器官以及生境和器官的交互作用對植物化學計量特征的影響。
關于植物化學計量內穩性的強弱,本研究采用Sterner和Elser提出的化學計量內穩性計算模型[4]:y=cx1/H進行計算,其中x為生長基質的元素(N、P)含量以及化學計量比(N∶P),y為植物不同器官的元素(N、P)含量以及化學計量比(N∶P),c為常數,H為內穩性指數。依據所測得的植物和不同生長基質的N、P元素含量及化學計量比,分別計算出各元素及元素比率的內穩性指數。
2 結果與分析
2.1 不同生境下長葉粗筒苣苔生長基質的C、N、P含量及化學計量比
根據測試結果表明,生長基質中C、N、P元素含量在不同生境間的差異不顯著,其分布范圍分別為292.1—379.73 g/kg、15.16—20.23 g/kg、1—1.17 g/kg,其中附生植株生長基質中的全C、全N和全P含量要高于地生植株生長的林地土壤(圖1)。在林地、樹干基部及樹干1.3 m處3種生境的基質中,C∶N、C∶P和N∶P比值存在一定差異,但其差異均未達到顯著水平。不同生境基質的C∶N∶P比也有所不同,其中林地土壤的C∶N∶P比為294∶15∶1,樹干基部土及樹干1.3 m處樹皮腐殖質的C∶N∶P比分別為313∶17∶1和348∶19∶1。
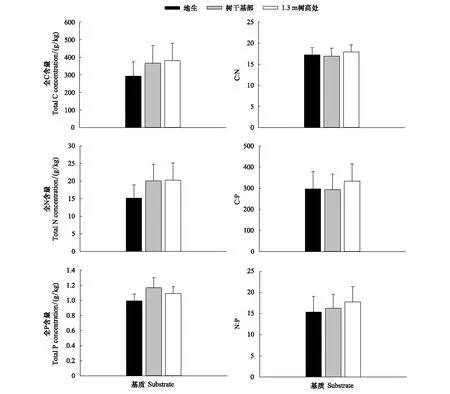
圖1 不同生境下長葉粗筒苣苔生長基質C、N、P化學計量特征Fig.1 Stoichiometric characteristics of C, N, and P in substrate of Briggsia longifolia among different habitats
2.2 不同生境下長葉粗筒苣苔葉片和根系的C、N、P含量及化學計量比
雙因素方差分析結果表明,生境變化對長葉粗筒苣苔的P含量以及N∶P比值具有顯著影響,器官對植物的N含量、C∶N以及N∶P比值具有顯著影響,而生境和器官的交互作用對植物的元素含量及化學計量比的影響均不顯著(表1)。
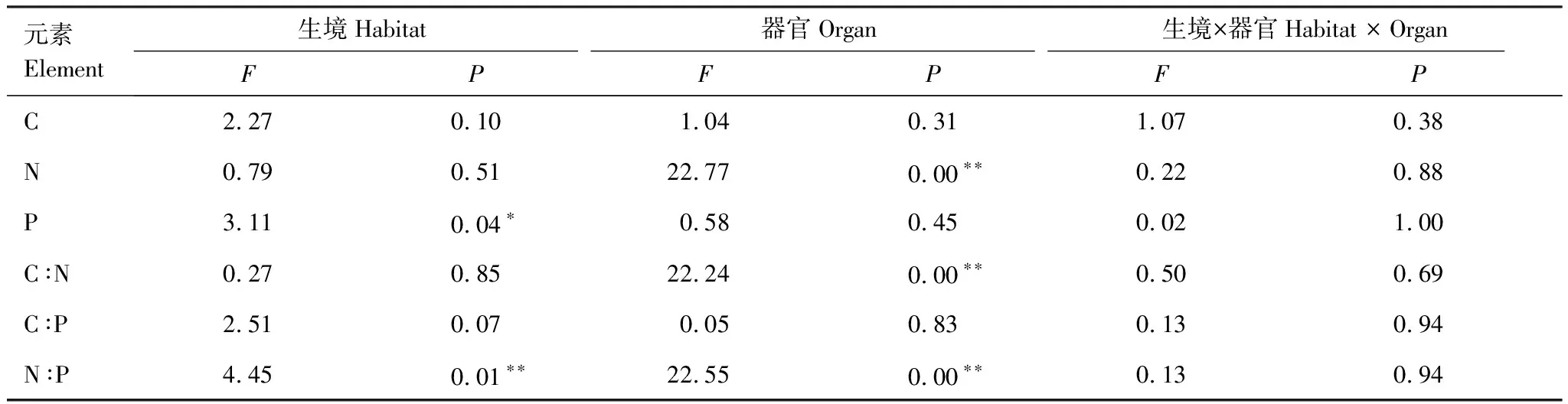
表1 生境、器官以及其交互作用對長葉粗筒苣苔的化學計量特征的影響
長葉粗筒苣苔葉片和根中C、N、P元素含量以及C∶N、C∶P及N∶P比值在不同生境間的差異均未達到顯著水平(圖2)。不同生境下長葉粗筒苣苔葉片的C、N、P元素含量范圍分別為440.86—448.50 g/kg、14.24—14.98 g/kg、1.33—1.83 g/kg,根系中C、N、P元素含量范圍分別為427.42—461.50 g/kg、18.04—20.37 g/kg、1.26—1.75 g/kg。地生植株葉片中的C∶N、C∶P及N∶P、根系中的C∶P和N∶P均高于樹干基部處和1.3 m樹高處的附生植株。地生植株葉片及根系的C∶N∶P比均高于附生植株,其中葉片的C∶N∶P比高于根系。在3種生境下長葉粗筒苣苔葉片的N含量、N∶P比值均顯著低于根系 (P<0.05),C∶N則顯著高于根系 (P<0.05)。
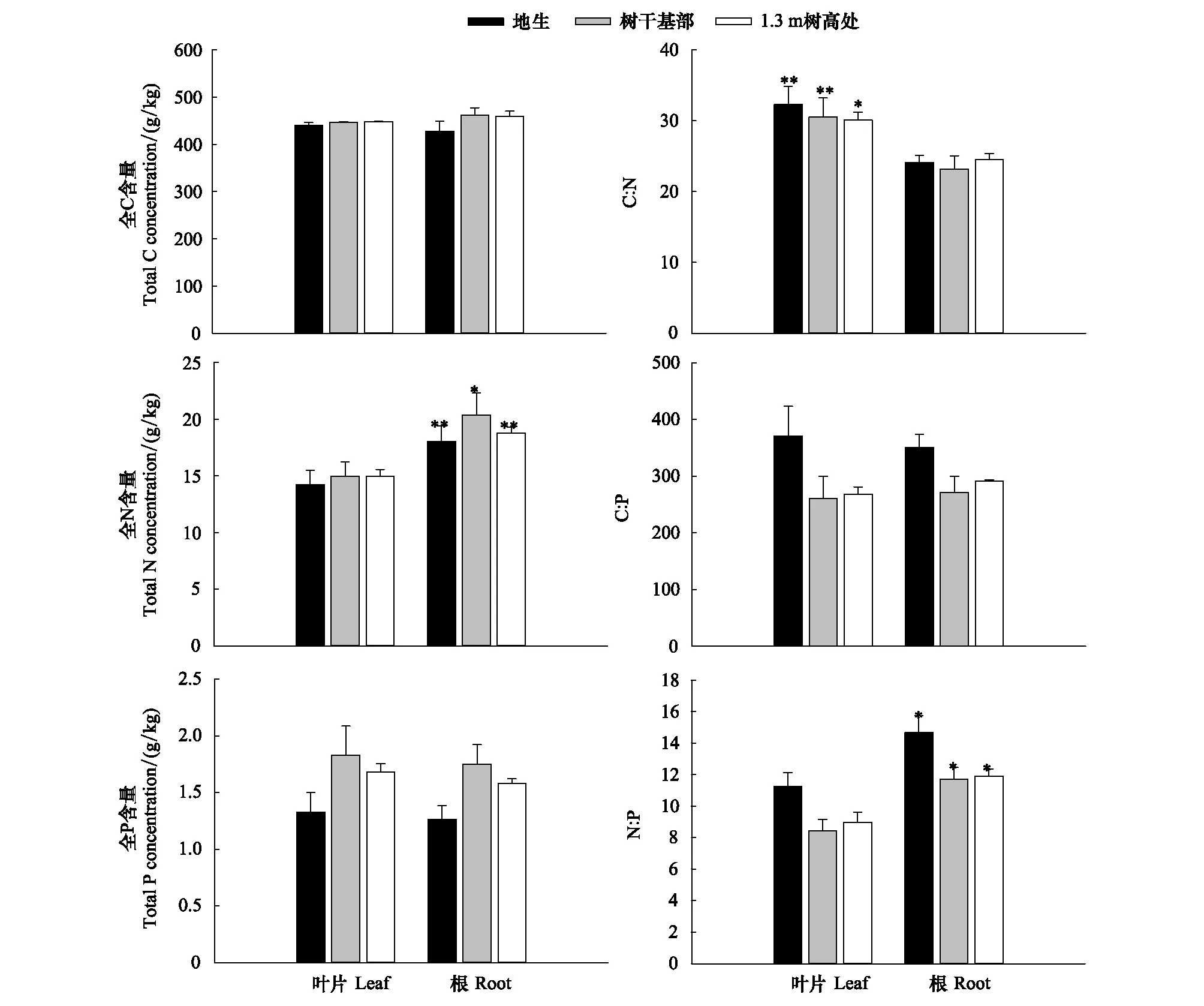
圖2 不同生境下長葉粗筒苣苔葉片和根系的C、N、P化學計量特征Fig.2 Stoichiometric characteristics of C, N, and P in leaf and root of Briggsia longifolia in different habitats “*”和“**”指示同一生境中葉片和根間差異的顯著性水平(P<0.05,P<0.01)
2.3 長葉粗筒苣苔葉片和根系的化學計量內穩性特征
長葉粗筒苣苔葉片N含量以及N∶P比值的化學計量內穩性指數均大于根系,但兩器官間的內穩性指數的差異未達到顯著水平(表2)。長葉粗筒苣苔葉片與根系中化學計量內穩性指數的大小排序均表現為:HN∶P>HN>HP。根據化學計量內穩性強弱的界定范圍(H> 4:穩態型;2 表2 長葉粗筒苣苔葉片和根系C、N、P化學計量內穩型模型統計與內穩性指數(H) 本研究結果表明,長葉粗筒苣苔葉片的C、N和P的平均含量分別為445.81 g/kg、15 g/kg和1.59 g/kg,其中C含量低于全球 492 種陸生植物的葉片C平均含量(464 g/kg),也低于本研究區哀牢山森林群落優勢喬木幼苗C含量[26],但高于中國草原生態系統213種植物的C平均含量(438 g/kg)[27]。 N含量低于全球陸生植物和中國陸生植物,而P含量則較陸生植物高[28—29],這表明此研究區內長葉粗筒苣苔的生長可能受N元素限制。 植物的葉片N∶P值在一定程度可用來判斷其生長環境中的N、P元素的受限狀況[5]。有學者認為當植物葉片N∶P<14 時,植物的生長主要受N元素限制;當N∶P>16 時,植物的生長主要受P元素限制;當 14 植物葉片的C∶N和C∶P比值的大小在一定程度上也能反映出其對N、P元素利用效率的高低,較高的葉面C∶N養分比可以增加單位養分的C儲量,即較高的C∶N和C∶P比代表植物對N和P的高度利用[6]。如,Pang等認為針葉樹的C∶N和C∶P比值較闊葉樹高說明針葉樹具有較高的N和P養分利用效率[31]。在附生生境中,養分供應有限且不穩定,水分變化劇烈,當附生生境養分充足時,附生植株通過對養分奢侈的吸收,并將多余的養分貯存在組織內,以此應對附生生境中的養分短缺和養分供應波動[24, 32]。而林地土壤養分供應較為充足,土壤含水量變化較為平緩,地生植株養分利用相對保守,受養分和水分脅迫的影響相對較附生植株要小。在本研究中長葉粗筒苣苔的附生植株葉片P含量高于地生植株,較高的P含量有利于緩解干旱脅迫對植物的生長造成的傷害[32—33]。附生植株葉片的C∶N和C∶P比均低于地生植株,表明附生植株對N、P元素利用效率較低。 由于植物不同器官的結構與組成以及功能的不同,植物不同器官的元素含量存在一定差異[34—35]。葉片作為多種生理功能(如光合作用、蒸騰作用和呼吸作用)的行使器官,需要較多的N和P來完成各種生化過程[36]。對3種濕地植物的研究結果表明,根系的N、P含量低于地上部分[36]。青藏高原窄葉鮮卑花地上部分的N、P元素含量高于根系[37]。本研究中,不同生境下長葉粗筒苣苔葉片與根系之間C、N和P元素含量及元素計量比的差異基本一致,雙因素方差分析的結果也表明生境和器官的交互作用對植物的化學計量特征的影響不顯著,這說明生境變化對植物葉片與根系之間的元素分配的影響較小[38]。長葉粗筒苣苔地生與附生植株葉片的N含量和N∶P均顯著低于根系,而C∶N則顯著高于根系。葉片中較低的N含量與植物的營養物質在非生長季向地下部分轉移有關,而較高的C∶N則可能由其較低的N含量導致,植物中C元素含量較為穩定,C∶N的變化主要受N元素影響[39]。此外,本研究區內長葉粗筒苣苔的生長受P元素限制不明顯,葉片和根系中P元素含量的差異不顯著,葉片中較低的N含量導致其N∶P顯著低于根。 不同功能群通過生理生態過程調整其自身對不同元素的吸收、運輸、分配、利用和釋放,以不同的生態適應策略來適應其各自的環境[40];而且同種植物的不同器官通過對環境變化的適應而表現出不同的內穩性特征[10]。因此,植物不同功能群及不同器官的化學計量內穩性特征可能存在一定差異。在草原生態系統,優勢物種羊草地上部分的N、P以及N∶P的內穩性指數均高于地下部分,其通過內穩性較低的地下部分貯存養分以保證地上部分的養分供給[10]。濕地植物根系的內穩性指數HN與葉片差異不顯著,但顯著高于莖[41]。哀牢山森林群落優勢喬木幼苗葉片和根系的N元素的內穩性指數HN分別為4.71、2.58[30]。在本研究中,長葉粗筒苣苔葉片N元素內穩指數HN為7.25,大于其根系HN(為5. 85),兩者均高于同一森林群落內優勢喬木幼苗。長葉粗筒苣苔較高的化學計量內穩性指數,且葉片內穩定指數大于根系,表明其對多變的環境適應性更強[42—43],它們主要通過調節根系的元素組成來維持葉片代謝活動的相對穩定,以此來適應生境的轉變[10]。 N、P在植物體的生長發育過程中行使不同的功能,導致其含量以及內穩性特征具有一定的差異[5]。生物體內含量較高的元素的內穩性較含量低的元素高,如N的內穩性高于P[44]。內蒙古草原維管植物的N元素的內穩性均高于P元素[45];濕地草本植物N元素的內穩性也較P高[41]。本研究中,長葉粗筒苣苔的N、P元素內穩性指數也表現出類似的規律[41, 44],表明植物對N元素的調控能力較強。根據養分限制性假說,即植物對易受限制的元素的控制能力較強,兼性附生植物對組織內N元素較強的控制能力也說明其生長更易受N元素限制[41]。此外,長葉粗筒苣苔葉片和根系的N∶P的內穩性HN∶P均強于N、P元素,這可能與植物中N、P元素含量具有較強的協同性[46],因而其元素比值的變異性較小[10]有關。 兼性附生植物長葉粗筒苣苔既能生長在樹皮上,又能根植于林地土壤中,具有獨特的生態適應能力。與地生植株相比,附生植株葉片的N、P含量較高,而C∶N、C∶P和N∶P比值均較低,說明其養分利用率較低;葉片的N、P以及N∶P比的內穩性指數高于根系。長葉粗筒苣苔通過調整其在不同生境間的養分吸收與利用策略,以此來適應生境的變化。因此,探討兼性附生植物在不同生境下的化學計量特征及其生態適應策略,豐富了植物生態化學計量學研究的理論和方法,也為深入研究森林生態系統生物多樣性形成及其維持機制、以及山地森林資源的保護與管理提供了科學依據。 致謝:感謝哀牢山生態站提供的幫助,感謝西雙版納熱帶植物園昆明分部地球化學實驗室在樣品元素含量分析中提供的幫助。
3 討論
3.1 不同生境下兼性附生植物的生態化學計量特征
3.2 兼性附生植物化學計量的內穩性特征
4 結論
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
