華南五針松瀕危機制生態學研究
2022-08-31 06:08:38崔國發
生態學報 2022年15期
關鍵詞:物種
王 俊, 潘 鴻, 謝 磊, 崔國發
北京林業大學生態與自然保護學院, 北京 100083
種群分布格局不僅能反映某個種群適應生境后對空間占有的一種形式,而且在不同生境梯度下植物種群在其定植、生長的過程中所經歷的生態學過程不盡相同[1—5]。在一定的尺度范圍內,通過對種群分布格局的分析,可以對種群的水平結構進行定量描述,也可以進一步揭示植物群落特征的基礎和格局的成因,闡述種群及群落的動態變化等,從而為植物保護、就地保護或遷地保護的實施提供一定的科學依據[6]。
群落組成與結構是由決定性因素(如生態位需求和種間作用因素)、隨機因素(定居和干擾)兩個方面決定的,且群落組成種的相對多度和格局與其相對競爭能力有關[7—8]。植物受競爭影響程度取決于自身及相鄰植物大小、數量、植物利用資源的特性和空間影響區域,且種間競爭是植物間競爭的重要組成部分[9—11]。
華南五針松(Pinuskwangtungensis)又名廣東松,雖然其生態適應性較強,現代地理分布區范圍跨越大,但呈現出分布零散、數量少、海拔高等特點[12—16]。前人對華南五針松的研究集中在局部分布區的種群、群落特征和種間關系[17—21],以及華南五針松地理分布的主要限制因子,并討論了在氣候變化的情況下,華南五針松分布區的變化趨勢等[16,22]。至此,但在群落生態學上,國內外還未見有從華南五針松種群空間格局和生存壓力等方面探討其瀕危機制的研究。
當前,在全球氣溫升高、自然資源過度開發、環境污染日趨嚴重的情況下,通過在種群生態學基礎上定量計算不同海拔瀕危植物的生存壓力指數,提出植物間“競爭驅趕”假說,并從理論和實踐兩方面進行驗證,為瀕危植物的就地保護或近地保護研究提供理論基礎。華南五針松的自然演替處于衰退之中[23],研究華南五針松的瀕危原因不但為其他五針松組植物的瀕危機制提供重要素材外,也為其它物種的瀕危機制研宄提供了案例。
1 研究區概況
廣東南嶺國家級自然保護區地處廣東省北部韶關市的乳源縣、清遠市的陽山縣和連州市行政境界內,保護區面積58368.4hm2。地理坐標為112°30′—113°04′E,24°37′—24°57′N。總體上屬于花崗巖山地的中山山地,區內山高谷深,地勢峻峭,最高峰石坑嶺海拔1902m,為廣東省第一高峰,其次為紫云山海拔為1888m,小黃山最高海拔為1600m。氣候屬典型的亞熱帶溫濕氣候,因地勢較高,具山地氣候特色,年平均氣溫17.7℃,最高溫34.4℃,最低溫-3.6℃;降水量較充沛,多集中3—8月,年平均達1705mm,最高年份達2495mm,年平均相對濕度84%;土壤類型隨海拔高度不同,PH值在4—5.5,海拔900—1800之間為山地黃壤[24]。
廣東南嶺國家級自然保護區是廣東珍稀瀕危針葉樹種最集中的分布區,在乳陽林區中山山地的小黃山和紫云山,保存著1300hm2的華南五針松原始林,植株多,面積大,以華南五針松、五列木(Pentaphylaxeuryoidex)、長苞鐵杉(Tsugalongibracteata)、福建柏(Fokieniahodginsii)為優勢種的原始針闊混交林。
南嶺地質歷史悠久,是植物物種南北交流和東西交匯的重要過渡地區,是我國中亞熱帶與南亞熱帶重要的氣候分界線,在阻擋南北氣流的運行方面發揮著重要作用,是華南五針松的核心分布區,同時也是我國亞熱帶常綠闊葉林的主要分布區[25]。
2 研究方法
2.1 文獻收集法
根據《中國植被圖(1∶100萬)》和《中國植物志》等相關植物方面的書籍,查找出在南嶺地區不同海拔華南五針松所處的典型植被,并根據不同植被型中的建群種或共優種,記錄其生物學高度、胸徑和冠幅等,計算出華南五針松的平均壓迫力。
2.2 樣地調查法
于2018年4月底至5月初,在植物標本采集和踏查的基礎上,根據植物的分布和具體生境,采用典型抽樣調查法,在華南五針松生存最低海拔下限和最高海拔上限之間,沿垂直高度以60—200m為海拔梯度設置樣地,利用“種—面積曲線”法求算群落的最小樣地面積。在同一分布區內的兩個山頭,共設置樣地14個,其中1個人工林(實驗區的樹木園)于1977年栽種,每個樣地面積400m2。樣地調查詳細情況如表1。
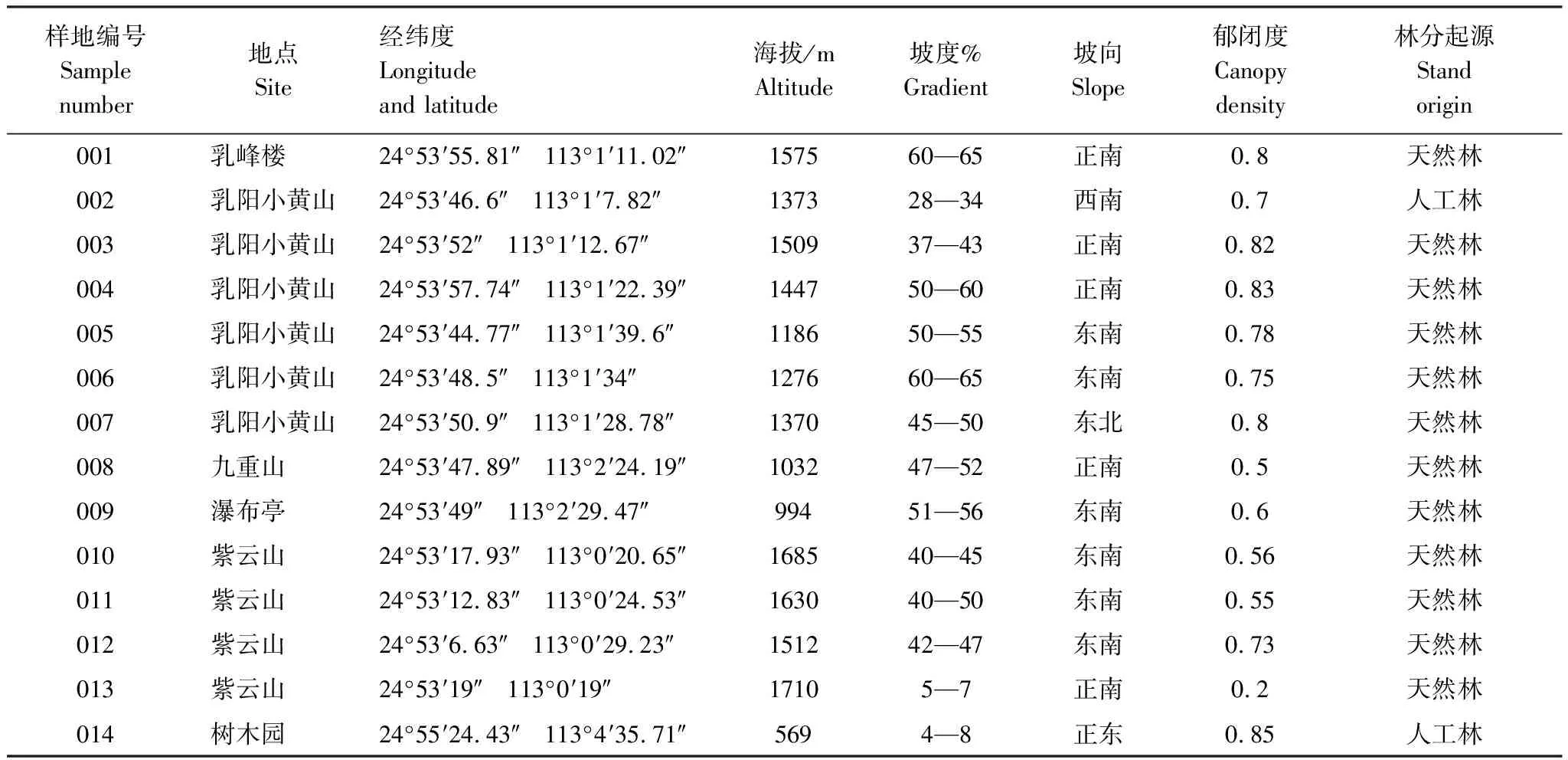
表1 調查樣地基本概況
樣地內華南五針松幼苗極小,且數量少,所以對樣方內胸徑(DBH)>4cm的喬木采用“每木調查法”,測定其胸徑、基徑、樹高、枝下高、冠幅等指標,簡單記錄灌木、草本植物的種類,同時以每個喬木樣方的兩邊為坐標軸測定每株樹木的相對坐標(X,Y),并記錄每個大樣方的生境因子,例如經緯度、海拔、坡度、坡向、坡位、郁閉度等。
2.3 計算方法
考慮到在植物群落內,植株個體之間的競爭往往取決于植株對生存環境和空間資源的爭奪,尤其表現在對光資源和水資源的競爭。在水資源不成為限制因子時,植株樹高越高、冠幅越大對光資源的競爭力越強,從而對其他植株的影響程度也就越大。胸徑相等的不同樹種間植株個體的樹高和冠幅大小往往存在很大差異,再者,樹冠大小相同而處在不同方位的植株個體對珍稀瀕危樹種個體造成的生存壓力也不同。
鑒于此,采用一種珍稀瀕危樹種個體生存壓力計算方法[11,26—27],能夠客觀、定量地反映珍稀瀕危樹種個體在植物群落中的生存壓力和平均壓迫力大小,并在此方法上進行了改進。
計算方法包括以下步驟:
(1)競爭木的確定:當某樹種的植株個體樹高即H大于等于其與珍稀瀕危樹種個體距離即L時,確定此植株為競爭木;
(2)計算珍稀瀕危樹種個體與競爭木的樹冠大小V,樹冠近似看作為橢圓柱,用橢圓柱體積量化樹冠大小;
(3)計算珍稀瀕危樹種個體承受的來自某一株競爭木的單株實際生存壓力指數PIij,理論壓迫力P;
式中,PIij為單株實際生存壓力指數,即珍稀瀕危樹種第i株個體承受第j株競爭木的生存壓力,PIij≥0;當競爭木與珍稀瀕危樹種個體的距離Lij等于其樹高Hij時,該競爭木無競爭效應,PIij為最小值0;當競爭木與珍稀瀕危樹種個體距離越近,樹冠越大,所處方位越接近珍稀瀕危樹種個體的正南方向時,PIij越大,當PIij≥1時,認為珍稀瀕危樹種第i株個體承受第j株競爭木的生存壓力較大;P為理論壓迫力;
Vij為第j株競爭木的樹冠大小,單位為m3;
Vi為珍稀瀕危樹種第i株個體的樹冠大小,單位為m3;
Di和Dij分別為第i株珍稀瀕危樹種和第j株競爭木的胸徑;
(4)計算珍稀瀕危樹種個體承受的來自某個競爭樹種即t的生存壓力指數PIit;
式中,PIit為單個樹種生存壓力指數,即珍稀瀕危樹種第i株個體承受的來自競爭樹種t的生存壓力;m為競爭樹種t的競爭木的株數;
(5)計算珍稀瀕危樹種個體承受的來自所有競爭樹種的生存壓力指數PIi:
式中,PIi為單個樹種總生存壓力指數,即珍稀瀕危樹種第i株個體承受的來自所有競爭樹種的生存壓力;n為競爭樹種總數。
(6)計算珍稀瀕危樹種承受的來自所有競爭樹種的平均生存壓力指數PI:
式中,PI為瀕危樹種平均生存壓力指數,即珍稀瀕危樹種承受的來自所有競爭樹種t的平均生存壓力;k為珍稀瀕危樹種總數。
3 結果與分析
3.1 沿海拔梯度上種群空間格局
種群的空間格局的計算方法很多,采用目前較為常用的具有相同本質或較大相似性的8種方法(方差/均值比率、t值檢驗、負二項參數、擴散型指數、Cassie指標、叢生指標、平均擁擠度和聚塊性指標等)綜合分析華南五針松種群的分布格局[6,28—31],計算結果見表2。
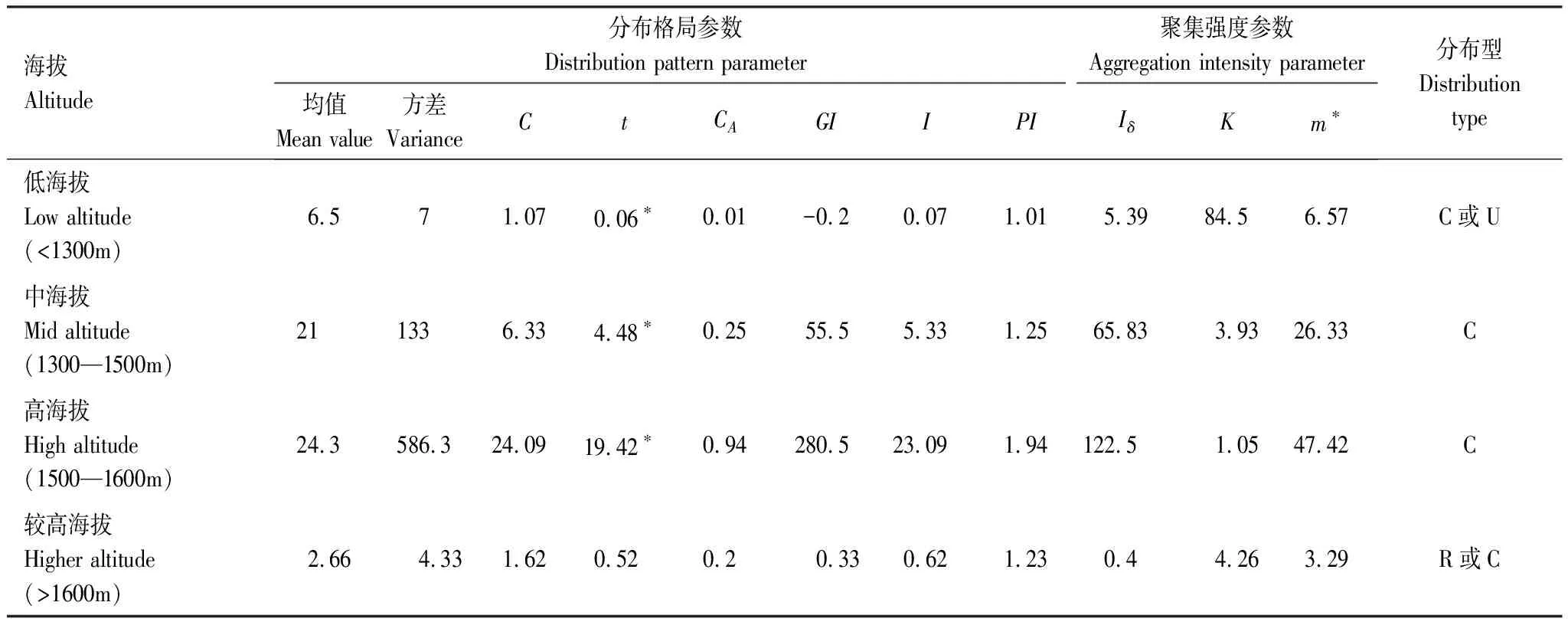
表2 不同海拔梯度上華南五針松種群空間格局
由表2可知,在不同海拔梯度上C、Iδ和PI均大于1,I和CA均大于0,而GI小于0。以上6個指標表明種群在中、高海拔處為聚集分布,低海拔為聚集或均勻,且C通過了t檢驗,進而驗證沿海拔梯上種群的分布格局與預期假設呈顯著差異。
隨海拔的升高,Iδ和m*的值逐漸增大,同時K值逐漸減少,由此表明,隨海拔梯度的增加(1600m除外),種群聚集強度增加,種群的擁擠程度也相應增加,種間和種內競爭激烈,競爭壓力增大;當海拔在1600m以上時,K值又變大,表明聚集程度變緩,生存壓力減小。
3.2 “競爭驅趕”假說的提出與驗證
植物為了其更好生存需最大限度的加大對養分、能源和空間等的爭奪,當兩個或者多個植物所要求的資源,空間等發生重疊時就發生種內和種間競爭,樹高冠密的個體就會占據優勢,欺負其他植物,驅使他們逃離到生存壓力小的地方或死亡,這就是“競爭驅趕”假說。
本次調查中,在華南五針松生存的最低海拔下限和最高海拔上限之間,沿小黃山和紫云山共設置13塊樣地,共測175棵華南五針松。從圖1中可以得出:沿海拔梯度的上升,華南五針松種群平均生存壓力指數整體上呈遞減趨勢(R2=0.31,P<0.05),海拔較高的區域生存壓力指數較小,利于華南五針松種群的生存。
在中高海拔1509m處物種豐富,種群密度大,伴生種的平均樹高和冠幅均高于華南五針松,種群競爭激烈,造成華南五針松種群平均生存壓力指數較大,在擬合回歸中出現突變點。
南嶺屬亞熱帶季風濕潤氣候,其水平地帶性植被為中亞熱帶南部亞地帶常綠闊葉林。由表3得出,不同海拔區段呈現出不同的植被型,華南五針松適生于中上海拔的針闊混交林內,這跟實際調查的華南五針松分布海拔是一致的;《中國植物志》中記錄華南五針松高達30 m,胸徑1.5m,分布海拔為700—1600 m,而在實際調查中最低海拔為994 m,最高海拔接近1710 m,這種分布海拔區域的變化極有可能是由于闊葉樹種的存在,且樹高冠密,平均壓迫力較大,從而驅使針葉樹種有一定程度上的提升。
不論從理論(表3)和實際調查(圖1)中發現,華南五針松是有一個暫時穩定的群落環境,但隨著群落內植物種內和種間競爭驅趕,全球氣溫升高,且華南五針松種群喜溫涼濕潤,而高海拔生存壓力較小,受到的阻力就小,種群更加傾向于高海拔分布,但當種群分布上移到達山頂時,會出現溫度急劇降低,風大霧多,土層淺薄等現象,若干年后華南五針松則無處存活被迫退出群落,導致種群的滅亡。
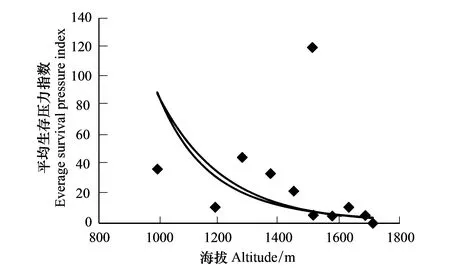
圖1 不同海拔梯度上華南五針松種群平均生存壓力指數 Fig.1 The everage survival pressure index of the population of Pinus kwangtungensis along different elevation gradients
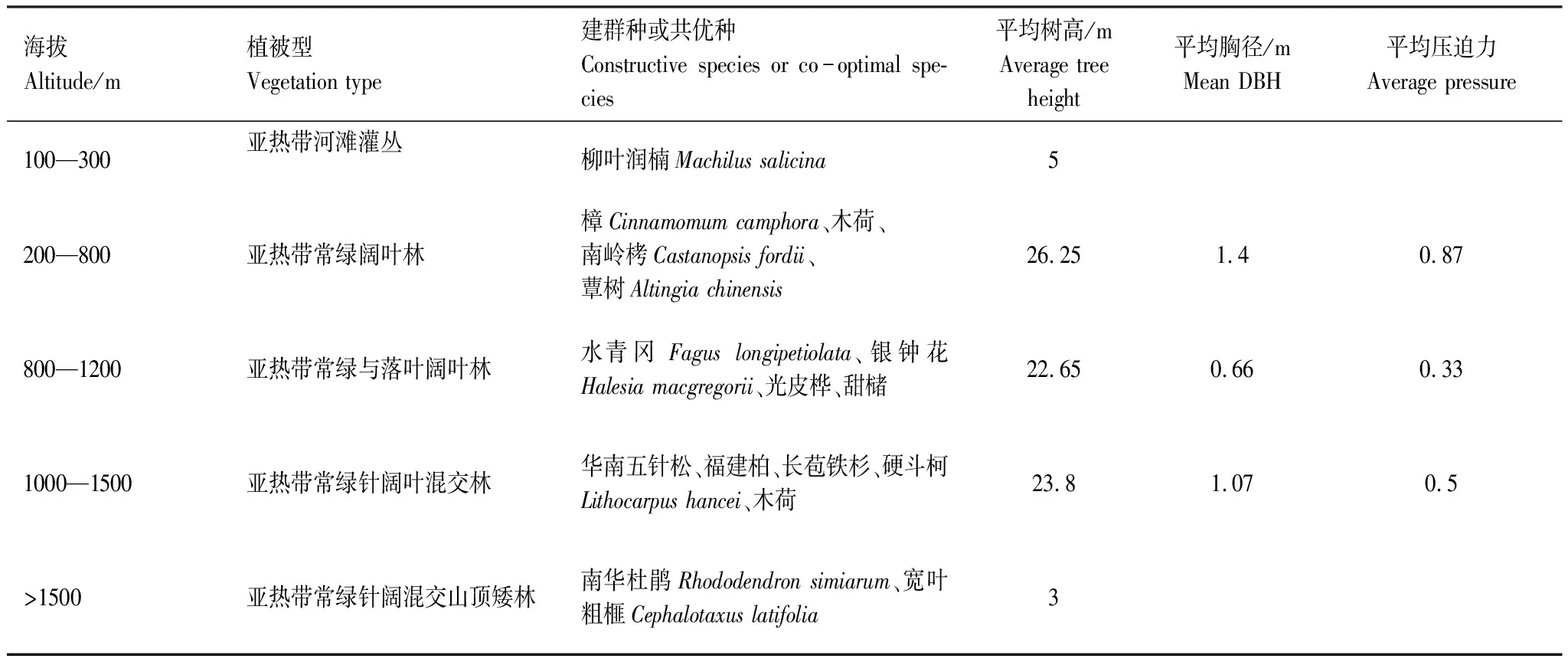
表3 不同海拔華南五針松在不同植被帶上的理論壓迫力
3.3 同海拔生存壓力
表4、圖2和圖3表明:在土壤質地、坡向和氣候基本相似的情況下,在海拔1509m處,華南五針松種群平均生存壓力指數為120,而在海拔1512m處的平均生存壓力指數是2.6,前者是后者的46倍。在1509m處,華南五針松成片存在,密度大,物種多樣性豐富,平均樹高9.33m,平均胸徑和冠幅分別為12.73cm和36m3,而競爭木像馬蹄荷(Exbucklandiapopulnea)、木荷(Schimasuperba)、南方鐵杉(Tsugachinensis)、五列木等樹種的平均樹高為12m,平均冠幅是100m3,種內和種間相互競爭,且以種內競爭為主,伴生種的主要競爭順序為木荷>甜櫧(Castanopsiseyrei)>五列木>馬蹄荷(Exbucklandiapopulnea)>馬尾松(Pinusmassoniana)>長苞鐵杉;在1512m處,華南五針松種群的群落結構較簡單,只有少量的南方鐵杉和長苞鐵杉等樹種,平均樹高不高于4.5m,以種內競爭為主,其他競爭樹種對其競爭生存壓力較小。
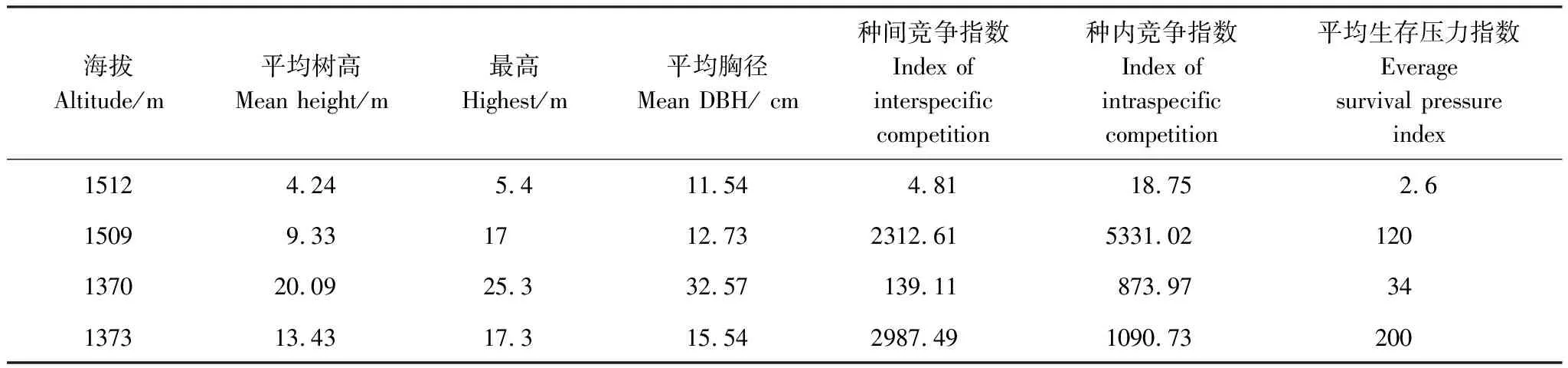
表4 同海拔華南五針松群落特征
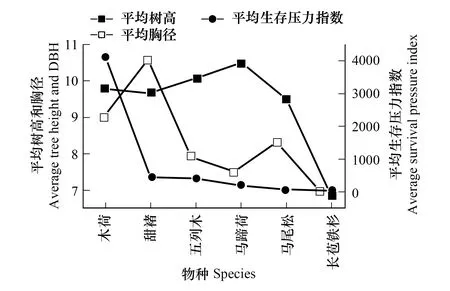
圖2 海拔1509m華南五針松群落特征及種間生存壓力 Fig.2 The characteristic of the community of Pinus kwangtungensis and the survival pressure of interspecific at 1509m
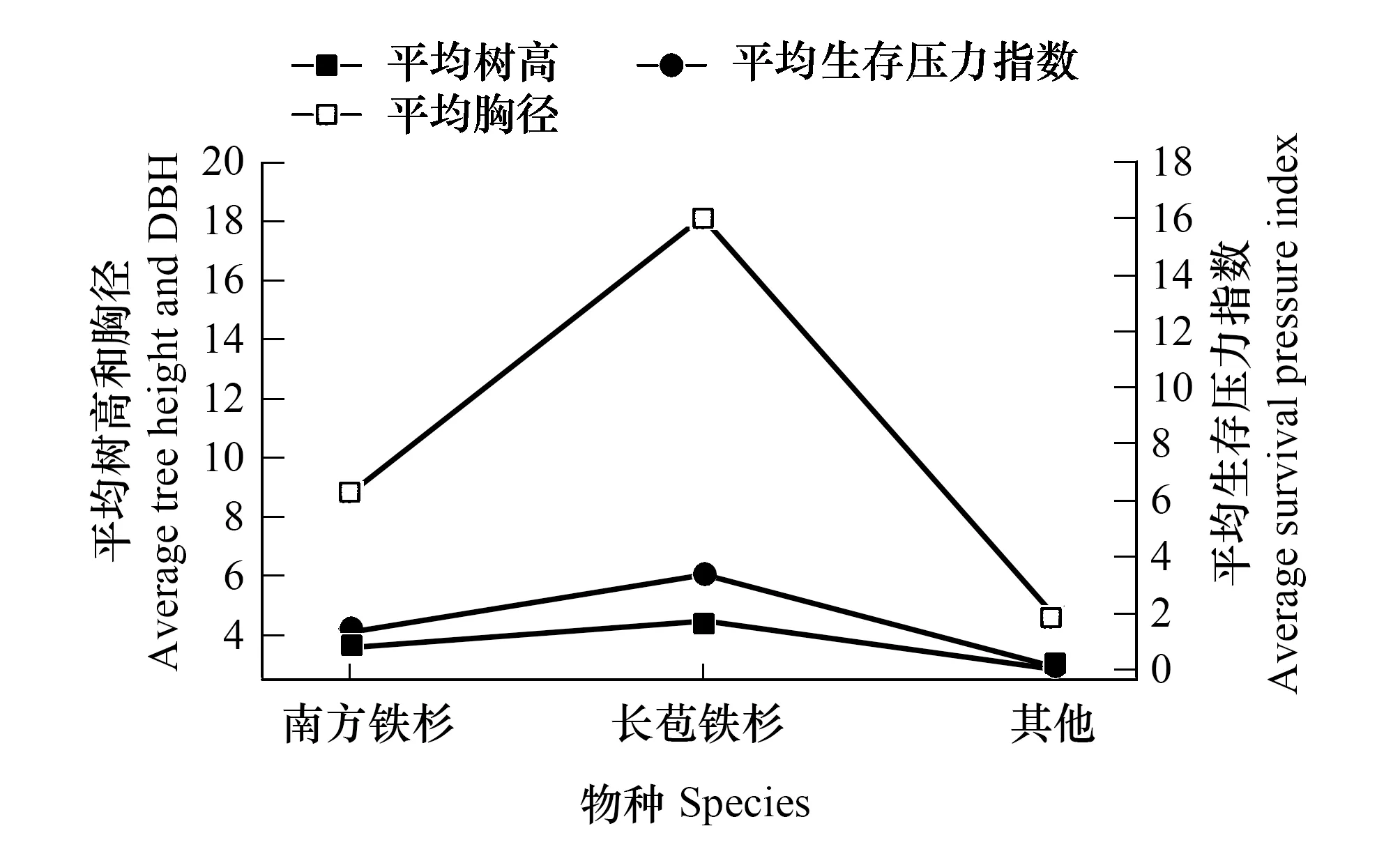
圖3 海拔1512m華南五針松群落特征及種間生存壓力 Fig.3 The characteristic of the community of Pinus kwangtungensis and the survival pressure of interspecific at 1512m
由表4、圖4和圖5得出:在土壤質地、坡向和氣候基本相似的情況下,在海拔1370m,華南五針松種群平均生存壓力指數為34,而在海拔1373m處的生存壓力指數為200,兩者相差約160。這主要是由于在在海拔1370m處,樣地為半陰坡,華南五針松天然生存,平均樹高20.09m,以種內競爭為主;雖為優勢種,但主要受青岡(Cyclobalanopsisglauca)、木荷、甜櫧等物種的競爭,競爭順序為青岡>木荷>甜櫧>五列木;而在海拔1373m處,樣地為半陽坡,屬華南五針松人工林,種群競爭壓力大,以種間競爭為主,伴生種的競爭順序為吳茱萸(Evodiarutaecarpa)>擬赤楊(Alniphyllumfortunei)>紅楠(Machilusthunbergii)>光皮樺(Betulaluminifera)>仁昌木蓮(Manglietiachingii)>乳源木蓮(Manglietiayuyuanensis),群落結構不合理造成競爭壓力巨大。
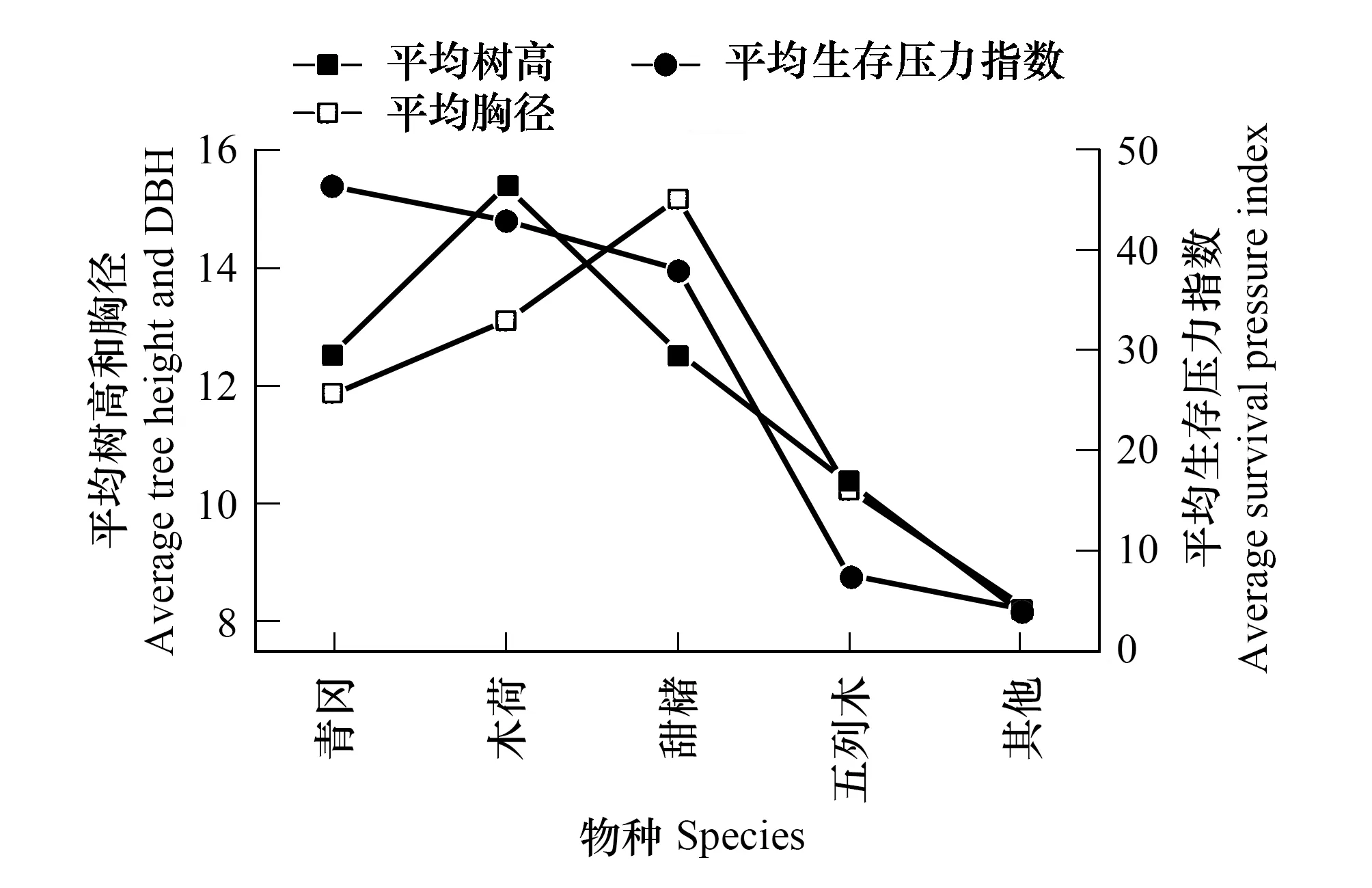
圖4 海拔1370m華南五針松群落特征及種間生存壓力 Fig.4 The characteristic of the community of Pinus kwangtungensis and the survival pressure of interspecific at 1370m
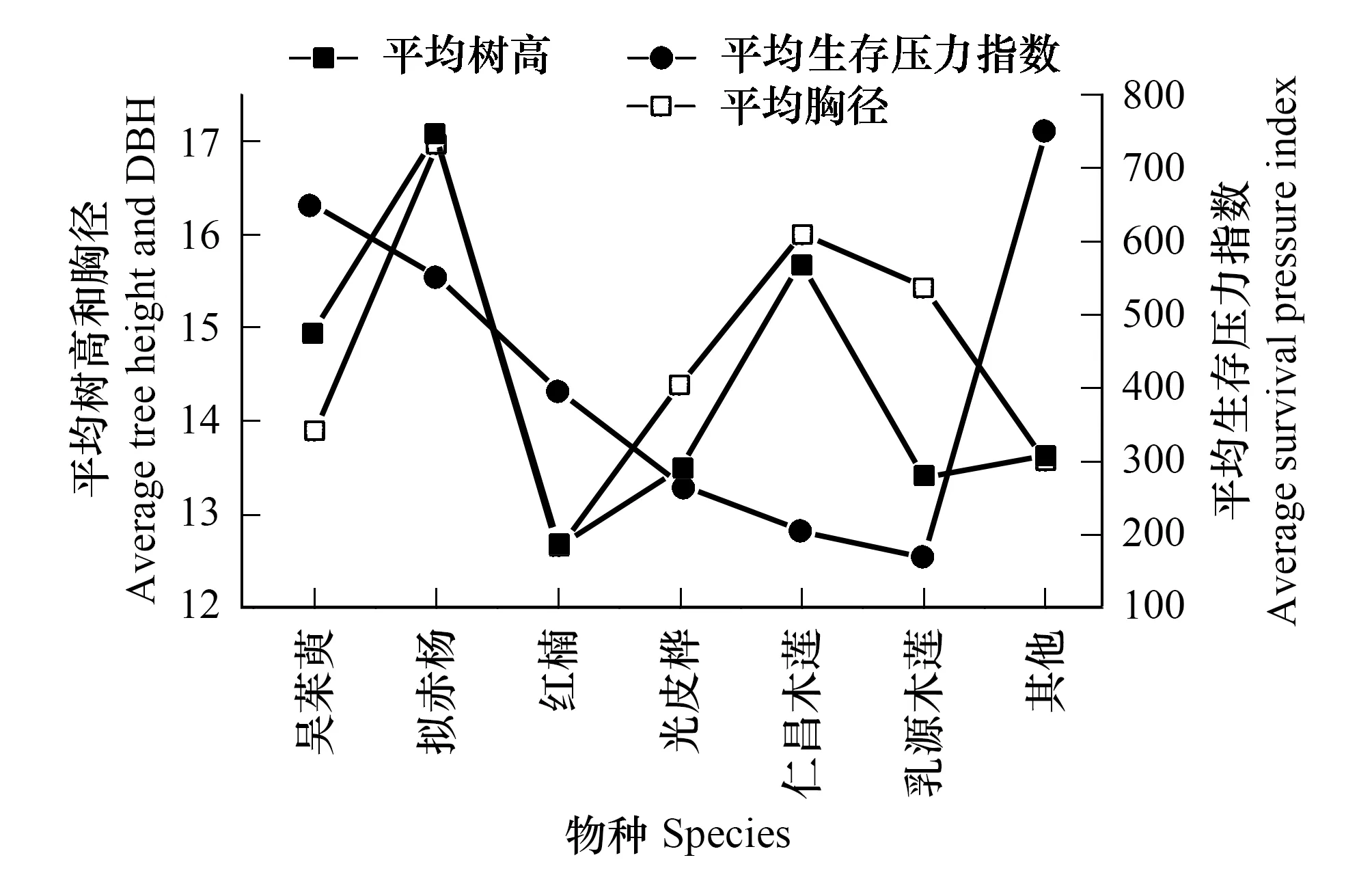
圖5 海拔1370m華南五針松群落特征及種間生存壓力 Fig.5 The characteristic of the community of Pinus kwangtungensis and the survival pressure of interspecific at 1373m
3.4 人工林的生存壓力指數
在低海拔569m處,保護區于1977年育苗種植14棵華南五針松,經過近40年的時間,現今只有4棵活立木,且種群平均生存壓力指數為50,競爭壓力大,若不加以嚴格保護,及時對競爭樹種進行整枝或擇伐處理,預計4棵華南五針松將會在10—20年時間里成為瀕死木。
4 討論
種群空間格局隨尺度的變化而不同,不同的空間格局特征由不同的因素所致,而本文只是在一定區域采用樣方分析法研究種群空間格局,且隨尺度的不斷變大,生物因素和非生物因素的作用加強,種群空間格局會相應的改變,下一步將從生態位的分化理論及對不同生境的偏好程度來分析華南五針松種群的適應性、物種共存和不同尺度下的分布格局。
植物瀕危或滅亡是由內在和外在等各種因素共同作用的結果,且物種間的空間分離減弱種間競爭,阻止或減緩競爭優勢物種對競爭劣勢物種的競爭排除作用,這有利于維持物種共存和物種多樣性維持[32]。華南五針松雖在主林層占據顯著地位,但在演替層和更新層中的優勢遠不及其他優勢種[19],隨著群落內種間、種內競爭的激烈,華南五針松種群在中高海拔處生存壓力最大,而華南五針松種群在沿海拔梯度上升過程中平均生存壓力指數整體上呈遞減趨勢,海拔較高的區域利于華南五針松種群的生存,物種會傾向于向高海拔遷移,降低其他優勢物種的競爭強度,這與在溫度升高下,華南五針松分布的最低海拔將不斷上升,南亞熱帶和北熱帶地區的華南五針松已經上移到山頂的結論[16]和群落種間競爭驅趕致使群落正向演替是華木蓮瀕危的一個重要原因[33]有相似之處。而本研究只是在一定區域內通過沿海拔梯度上計算種群生存壓力,提出“競爭驅趕”假說,并進行了驗證,但隨著時間的推移,華南五針松種群的數量會不會減少以及分布區的上限和下限會不會縮短都將進一步需要驗證。
華南五針松作為一種比較原始和古老的物種,幼苗難以成活,種群更新困難,對溫度、濕度比較敏感。由于氣候變暖將導致物種適宜生境的喪失和破碎化,已造成歐洲南部山脈地區大量物種的數量在過去10年里正在逐步減少,在地中海地區14個物種向山頂移動,分布區逐漸縮小,生存壓力大和不適應,從而引起高山植物滅絕或被其他物種取代[34—36],及在溫度對華南五針松地理分布起主導作用,其對高溫條件的要求極為嚴格,喜溫涼濕潤的環境下[16],隨著氣候帶北移,利用土壤、濕度、物候和降水等因素,結合區域氣候等模型,探討華南五針松分布區是否會北移及向高海拔遷移,以及關鍵致危因子和瀕危理論機制還需要進一步的研究。
造成物種瀕危的因素主要是內因或者外因,抑或是兩者的結合。“競爭驅趕”假說是在種群和群落基礎上推斷物種瀕危的一個方面,屬第一次提出,是對外因的一個重要補充。其生存壓力指數的計算綜合考慮了樹高、樹冠、距離、胸徑等生態因子,優于諸如鄰體干擾和Hegyi單木競爭等模型的計算方法,能夠更客觀、真實、定量反映珍稀瀕危樹種個體及種群在群落中的生存狀況,為物種瀕危的推斷提供了一個全新模式。
5 結論
華南五針松種群在中、高海拔處為聚集分布,低海拔為聚集或均勻分布,且隨海拔的升高,Iδ和m*的值逐漸增大,同時K值逐漸減少,種群聚集強度增加,當達到一定高度時,聚集強度降低,種內和種間競爭減緩,生存壓力變小,驗證“競爭驅趕”假說的存在。
在南嶺國家級自然保護區內,華南五針松種群天然分布的垂直最低海拔為1000m左右,最高海拔上限1710m左右,在理論分布上有所抬升,且隨海拔升高,華南五針松平均生存壓力變小,海拔較高的區域利于華南五針松種群的生存,但當物種繼續向高海拔遷移,這些物種將由于分布區逐漸縮小、生存壓力大和不適應等因素造成最終滅絕,因此本研究認為物種之間存在“競爭驅趕”,這可能是導致華南五針松瀕危的原因之一,為華南五針松植物瀕危機制的理論研究提供一個示例。
海拔高度和溫度影響植物的群落結構,在立地條件相似的情況下,由于華南五針松復雜的群落結構及激烈的種內、種間競爭,造成同海拔的華南五針松種群呈現不同的生存壓力及華南五針松人工林群落中種群生存壓力指數大于天然林群落。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
