乙烯利對苦瓜幼苗內源激素代謝及相關調控基因表達的影響
2022-09-01 08:24:54吳漢東李躍飛
西北植物學報 2022年7期
焦 陽,馬 英,張 振,吳漢東,李躍飛
(錦州醫科大學 食品與健康學院,遼寧錦州 121000)
植物生長調節劑是一類與植物激素類似的化學物質,在植物生長、發育等多個生理過程中表現顯著的調控作用,具有安全、作用面廣、用量少、見效快等特點,是近代生理學和農業科學上的一項重大研究進展。以吲哚丁酸和萘乙酸為標志,天然、人工合成的植物生長調節物質已經陸續應用到農業生產中[1]。乙烯利是20世紀90年代人工合成的一種化學物質,其主要成分為2-氯乙基磷酸,通過對植株進行噴施等處理,能夠釋放出氣態乙烯發揮功能。外源乙烯(ethylene)在植物生長發育期間,如果實成熟、葉片脫落、植株矮化生長、花芽分化等過程均可產生顯著的調節作用[2]。同時,乙烯作為一種信號因子可以誘發植物內源激素代謝過程發生變化[2],如孫亮等[3]對百合鱗莖組織進行外源乙烯的處理,發現對其組織內的脫落酸(ABA)、赤霉素(GA)、生長素(IAA)的含量均產生了顯著影響。這種現象在施用外源ABA、GA對葡萄和刺五加的處理過程中同樣得到了證實,表明內源激素的合成與外源調節物質之間存在著明顯的互作效應[4-5]。
外源生長調節物質作用于植物后,會引發基因表達水平的改變,進而影響相關內源激素的代謝過程。乙烯生物合成途徑中兩個重要的限速酶1-氨基環丙基-1-羧酸(ACC)合成酶(ACS)、氧化酶(ACO)的表達水平與乙烯產量間密切相關,在對西瓜[6]、中國南瓜[7]、番茄[8]、樹莓[9]、菠蘿[10]等作物進行外源乙烯利處理后,兩種酶的表達豐度均出現了顯著提高,大大提升了乙烯的產率,表明乙烯利可以有效刺激作物內源乙烯的合成。同時,有研究表明,外源乙烯利對厚皮甜瓜幼苗葉片中IAA和GA的合成具有抑制作用,對ABA的合成具有促進作用[11];乙烯利對轉基因油菜角果和種子發育階段的內源激素GA和IAA的生成均具有抑制作用,推測乙烯與內源植物激素相互作用,調節油菜籽角果和種子的發育和生長[12];西瓜幼苗在噴施乙烯利后,體內IAA、ABA、GA的含量均低于對照[13];黃瓜幼苗經乙烯利處理后,莖尖組織中的IAA、ZT含量上升,而GA3則表現為降低,有利于雌性花芽的分化[14]。
苦瓜為葫蘆科作物,外源乙烯可以調控苦瓜幼苗的生長發育以及花芽分化[15]。當前的相關報道更多側重于對植株農藝性狀以及內源激素含量的分析,雖鑒定了個別調控基因,但從整體分子調控網絡的角度,闡述乙烯利對內源激素代謝過程的研究仍較為缺乏。因此,本研究對苦瓜幼苗進行乙烯利處理,在分析內源激素含量的基礎上,結合高通量測序(High-throughput sequencing)技術,從基因轉錄水平挖掘相關信息,獲得苦瓜應對外源乙烯誘導的內源激素代謝靶點,初步明確相關調控基因的作用特點,進而為生產上調控瓜類作物的生長發育奠定理論基礎。
1 材料和方法
1.1 試驗材料
本試驗以苦瓜常規品種“綠爵士”為試材,在遼寧省農業科學院(41.7898°N,123.4206°E)日光溫室內播種,常規管理后出苗。在幼苗3~4葉期挑選生長勢一致的植株,將其分為對照組(CK)與處理組(ET)兩部分,分別對葉片噴施蒸餾水和160 μL/L 的40% (v/v)乙烯利溶液,共噴施2次,2次間隔3 d。在末次處理后的第48小時采樣用于轉錄組測序分析(圖1),末次處理后第7天采樣用于植物激素含量的測定。每一處理的樣品隨機選取10株幼苗,采集嫩葉和莖尖組織進行混合,經液氮速凍后置于-80 ℃超低溫冰箱凍存備用,實驗共設置3次生物學重復。
1.2 測定指標及方法
1.2.1 1-氨基環丙基-1-羧酸(ACC)含量的測定稱量約0.2 g植物樣品,于液氮中研磨后轉移至50 mL離心管中,加入5 mL去離子水,在超聲清洗儀中水浴30 min。然后于4 ℃下離心(10 000 r·min-1)5 min后取上清液,將上清液pH調至4.0。加入20 mL氯仿振蕩混勻,4 ℃下離心(10 000 r·min-1)5 min,取上清液過MCX小柱,并以2 mL甲醇,1 mL去離子水,5 mL氨水(1 mol·L-1)淋洗柱子。過0.22 μm濾膜,用于高效液相色譜串聯質譜(HPLC-MS/MS)檢測。HPLC分析條件:色譜柱(100 mm×2.1 mm×1.7 μm),A相流動相為水/甲醇(98/2,V/V)(添加0.05%甲酸和5 mmol·L-1乙酸銨),B相流動相為乙腈,流速為0.3 mL·min-1。MS/MS條件:氣簾氣15 psi,霧化溫度400 ℃,噴霧電壓4 500 V,霧化器壓力65 psi。
1.2.2 ACS活性測定稱量約2.0 g植物樣品,于液氮中研磨后轉移至預冷的15 mL離心管中,加入3 mL的Tricine提取液震蕩。4 ℃下離心(10 000 r·min-1)3 min,取2.4 mL上清液加入到預冷的15 mL離心管中。過GE Sephadex去鹽柱去除樣品中原始ACC,以3 mL的Tricine柱洗液洗脫柱子,收集粗酶液。取1.5 mL粗酶液加入150 μL反應buffer及150 μL的SAM溶液作為反應底物,25 ℃下溫浴2 h,加入200 μL的HgCl2溶液終止反應。過0.22 μm濾膜,用于高效液相色譜串聯質譜(HPLC-MS/MS)檢測,以新生成的ACC來計算ACS的活性。HPLC分析條件:色譜柱(100 mm×2.1 mm×1.7 μm),A相流動相為水/甲醇(98/2,V/V)(添加0.05%甲酸和5 mmol·L-1乙酸銨),B相流動相為乙腈,流速為0.3 mL·min-1。MS/MS條件:氣簾氣15 psi,霧化溫度400 ℃,噴霧電壓4 500 V,霧化器壓力65 psi。
1.2.3 內源激素IAA、GA3、tZR含量的測定準確約1.0 g植物樣品,于液氮中研磨后加入10 mL異丙醇/鹽酸提取緩沖液,4 ℃震蕩30 min,再加入20 mL二氯甲烷震蕩30 min。于4 ℃下離心(13 000 r·min-1)5 min后取下層有機相。避光下氮氣吹干有機相,以400 μL甲醇(添加0.1%甲酸)溶解。過0.22 μm濾膜,用于高效液相色譜串聯質譜(HPLC-MS/MS)檢測。HPLC分析條件:色譜柱(150 mm×2.1 mm×2.7 μm),A相流動相為甲醇(添加0.1%甲酸),B相流動相為水(添加0.1%甲酸),梯度洗脫流速為0~1 min時20%A,1~9 min時80%A,10~10.1 min時20%A,10.1~15 min時20%A。MS/MS條件:氣簾氣15 psi,霧化溫度400 ℃,噴霧電壓4 500 V,霧化器壓力65 psi。
1.2.4 轉錄組高通量測序使用Trizol法提取6個試驗樣本的總RNA,使用Bioanalyzer 2100和RNA 1000 Nano LabChip Kit分析總RNA的產量和純度,獲得質量合格的樣本用于文庫的構建。使用Oligo(dT)寡聚磁珠富集純化RNA,將其片段化后反轉錄為cDNA,創建最終的cDNA文庫,在Illumina Hiseq-4000高通量測序平臺上機檢測。
1.2.5 生物信息學分析對原始轉錄本(raw reads)去除接頭序列以及低質量堿基信息,獲得有效轉錄本(clean reads)。采用從頭組裝的方式,使用Trinity 2.4.0進行轉錄本的整合[16]。將組裝后的Unigenes通過DIAMOND比對到Non-redundant(Nr)、Gene Ontology (GO)、Swiss-Prot、Kyoto Encyclopedia of Genes and Genomes (KEGG)以及eggNOG數據庫[17]。通過TPM值計算Unigenes的表達量,以“FDR < 0.05”和“log2FC ≥ 1”為閾值,篩選兩個文庫間的差異表達基因(DEGs)。利用GOseq和KOBAS對差異表達基因進行GO和KEGG通路富集分析。
1.2.6 差異表達基因的qRT-PCR分析參考轉錄組中的TPM值,選擇8個差異表達基因,利用Primer Premier6.0軟件設計引物,進行熒光定量PCR分析(表1)。提取樣本總RNA,檢驗RNA產量及純度,對質量合格的RNA進行反轉錄。根據第1鏈合成試劑盒(TaKaRa,日本)中的反應體系和條件合成cDNA,以β-Actin基因作為內參基因,在Bio-Rad IQ5 Real Time PCR系統(Bio-Rad,美國)上進行實時定量檢測,采用20 μL反應體系,按2-ΔΔCt算法計算各差異基因的相對表達量[18]。
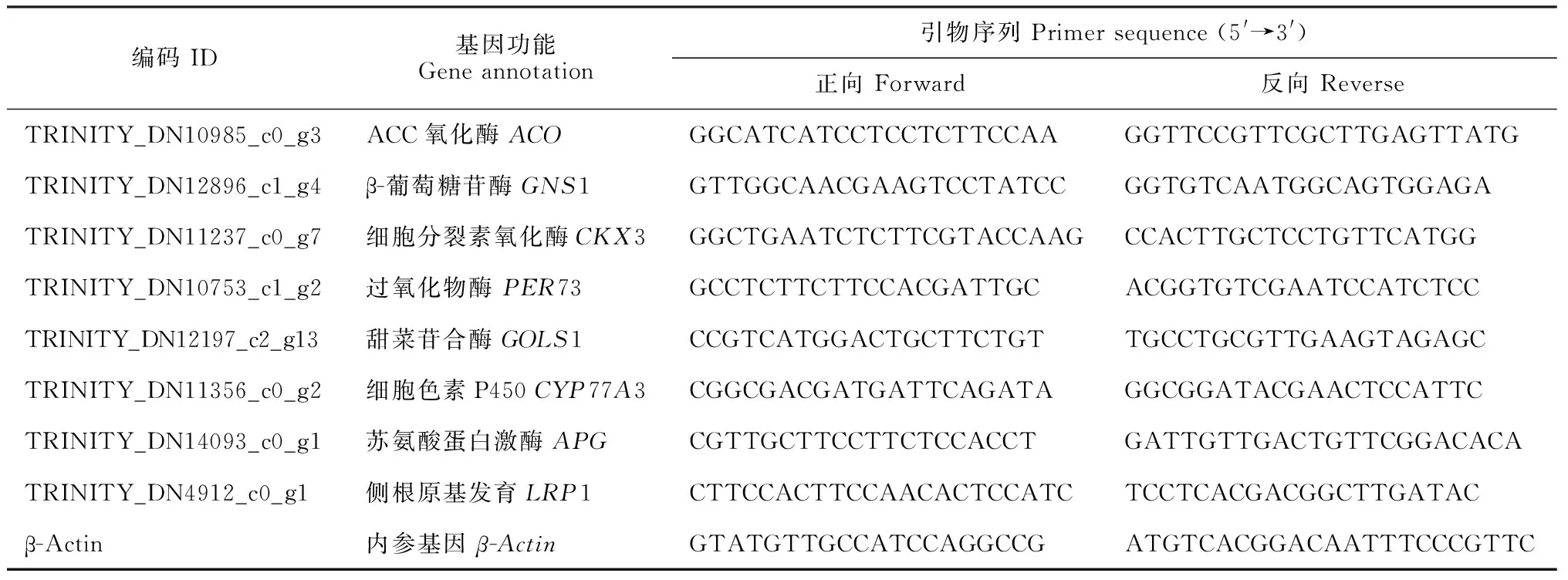
表1 qRT-PCR分析引物序列Table 1 Primers used for qRT-PCR analysis
2 結果與分析
2.1 外源乙烯利對苦瓜幼苗內源激素含量和ACS活性的影響
苦瓜幼苗經過葉面噴施乙烯利后,內源激素的代謝受到影響,內源激素含量發生了顯著的變化(表2)。其中,與對照(CK)相比,外源乙烯利處理苦瓜幼苗嫩葉和莖尖的1-氨基環丙基-1-羧酸(ACC)含量與ACS的活性均表現為顯著上升的趨勢,分別提升了23.2%和19.7%。由于ACS是ACC生物合成的限速酶,而且ACC是乙烯生物合成的直接前體,所以二者含量的升高有利于內源乙烯的合成。同時,外源乙烯利處理苦瓜幼苗的兩種植物激素IAA和tZR含量也比相應對照顯著上升,分別提高了57.5%和42.0%。與之相反,處理組幼苗GA3的含量卻比對照出現了顯著的下降,降幅達到了88.7%。上述結果表明,外源乙烯利處理能夠顯著誘發苦瓜幼苗激素水平的變化,同時誘導乙烯合成前體物質ACC含量和ACC生物合成限速酶ACS活性的顯著提高,從而促進內源乙烯的合成。

表2 外源乙烯利處理下苦瓜幼苗內源激素含量和ACS活性的變化Table 2 Endogenous hormonal contents and ACS activity of the bitter melon seedlings under exogenous ethephon treatment(ET)
2.2 外源乙烯利處理苦瓜幼苗轉錄組測序分析
通過對6個cDNA文庫進行轉錄組測序分析,共產生原始數據49.66 Gb。其中clean reads數目介于53 195 286~55 924 028之間,測序Q30值大于95%,GC含量大于45%,表明測序質量良好(表3)。對轉錄本進行組裝后,最終鑒定到了46 026個基因,平均長度494 bp。經過與各數據庫進行比對分析,在Nr、Pfam、Swiss-Prot、eggNOG、GO和KEGG數據庫中分別有59.37%、45.74%、41.88%、57.1%、51.86%和25.64%的基因得到了注釋。BLASTx比對結果顯示,同源序列命中比率最高的幾個物種分別為甜瓜(11385)、黃瓜(10580)、花燭屬紅掌(478)、葡萄(456)和水稻(446)。
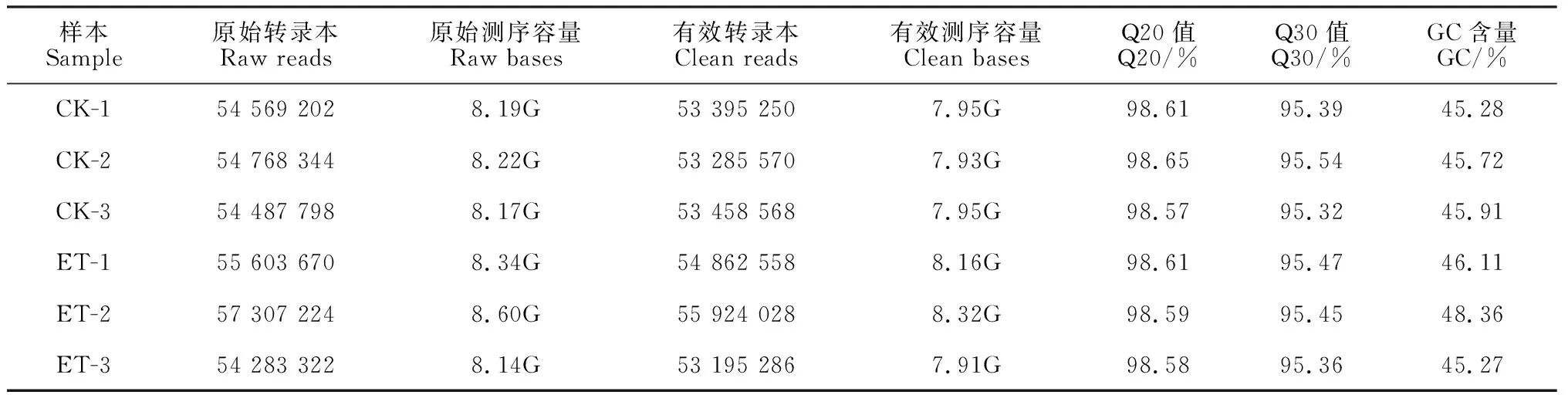
表3 cDNA文庫RNA-seq 測序結果Table 3 Summary of sequencing results based on RNA-seq from six cDNA libraries
進一步通過差異表達量分析,共獲得了1234個差異表達基因(DEGs),其中679個為上調表達,555為下調表達。GO富集分析顯示,共有980個DEGs得到了有效富集,包括具有“分子功能”的545個,“細胞原件”的179個,“生物代謝過程”的909個。此外,共有570個DEGs在KEGG庫中得到了有效富集,這些基因可以整合到118條代謝通路當中。其中包括植物-病原互作途徑(63DEGs)、植物激素信號轉導(41DEGs)、苯丙素的生物合成(39DEGs)、淀粉及糖代謝(35DEGs)、氨基糖和核苷酸糖的代謝(31DEGs)(圖2)。
2.3 外源乙烯利處理苦瓜幼苗參與內源激素合成的差異基因
為了進一步明確乙烯利誘導了哪些功能基因,進而影響內源激素的生物合成,基于數據庫比對信息,我們篩選了與激素代謝相關的差異表達基因(表4)。其中,在乙烯生物合成途徑中,共鑒定到了6個顯著上調表達的DEGs,包括1個ACS基因、1個ACO基因、3個HMT3基因,這些基因的表達趨勢與激素含量的變化情況一致。對于生長素合成途徑,1個ALDH基因和1個TAA1基因出現了顯著下調,另一個基因UGT74B1表現為顯著的上調。由于生長素合成途徑較為復雜,通常包含4條支路,因此上述基因的表達可能暗示著代謝支路發生了變化。與細胞分裂素代謝相關的基因表達水平也出現了改變,2個與其合成相關的CYP735A基因表達受抑,負責將細胞分裂素轉化為其他代謝產物的2個CKX基因則大幅度上調表達。對于GA的代謝途徑,只鑒定到1個參與其生物合成的上調表達基因KAO1,此外,通過對差異基因進行篩選,發現2個抑制其活性的CYP736A12-like基因高倍數上調表達。
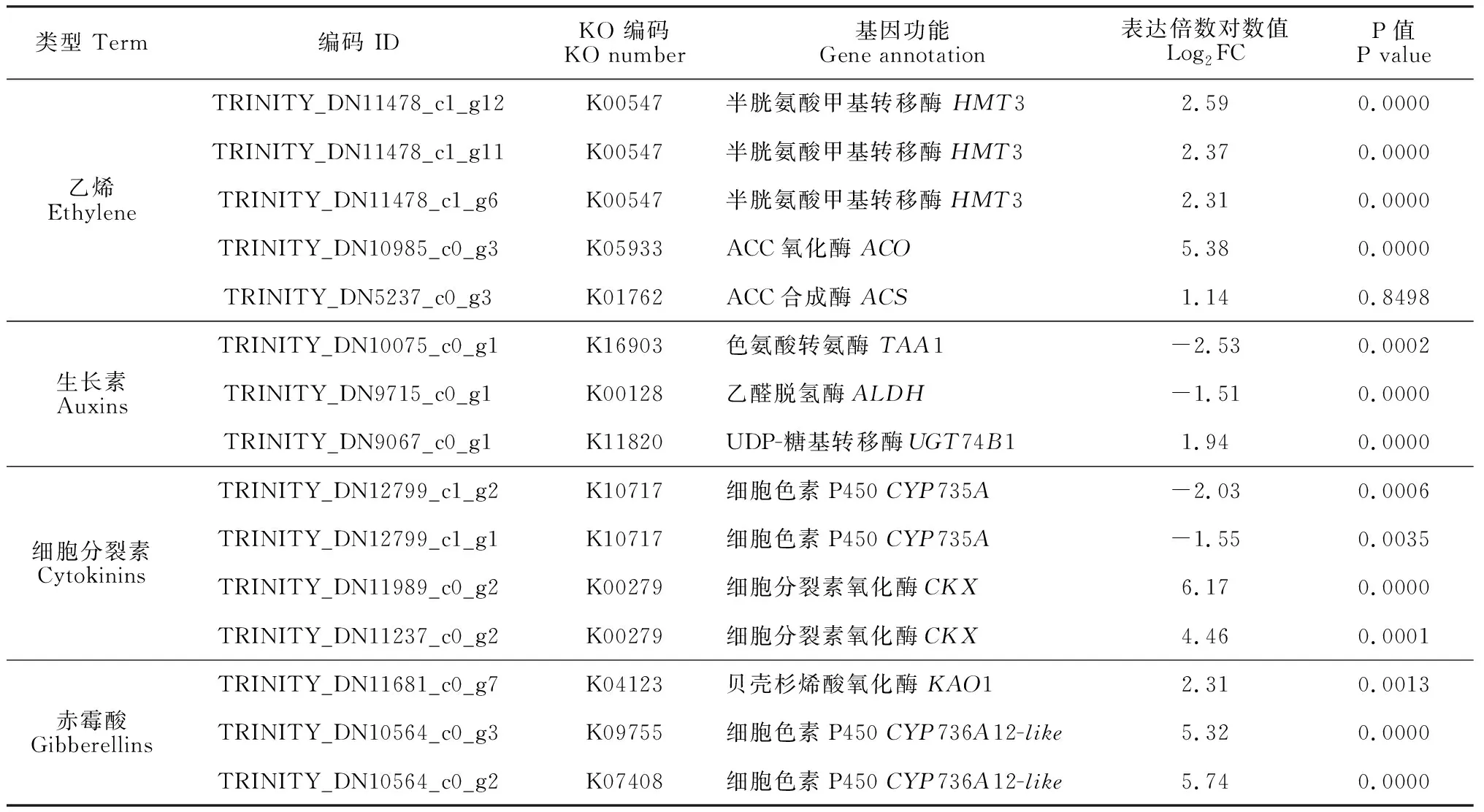
表4 與內源激素代謝相關的差異表達基因Table 4 Differentially expressed genes involved in hormonal metabolism
2.4 外源乙烯利處理苦瓜幼苗參與激素信號轉導的差異基因
內源激素通常以信號轉導的方式誘導下游一系列基因進行表達。通過對KEGG通路數據庫中map04075進行分析發現,在乙烯的信號轉導路徑當中,共有1個EIN3基因和11個ERF基因顯著上調表達。在生長素通路當中,3個負調控基因AUX/IAA的表達活性下降,1個ARF基因、1個GH3基因和3個SAUR基因表現為活性的上升。對于細胞分裂素通路來說,只獲得了1個上調表達的負調控基因A-ARR。在GA信號通路中,3個植株矮化基因GID1、1個PIF3基因表達活性顯著升高(表5)。上述參與激素應答的基因表達豐度的改變,表明乙烯利處理不僅影響內源激素的生物合成,同時也間接影響了下游基因的相應功能。
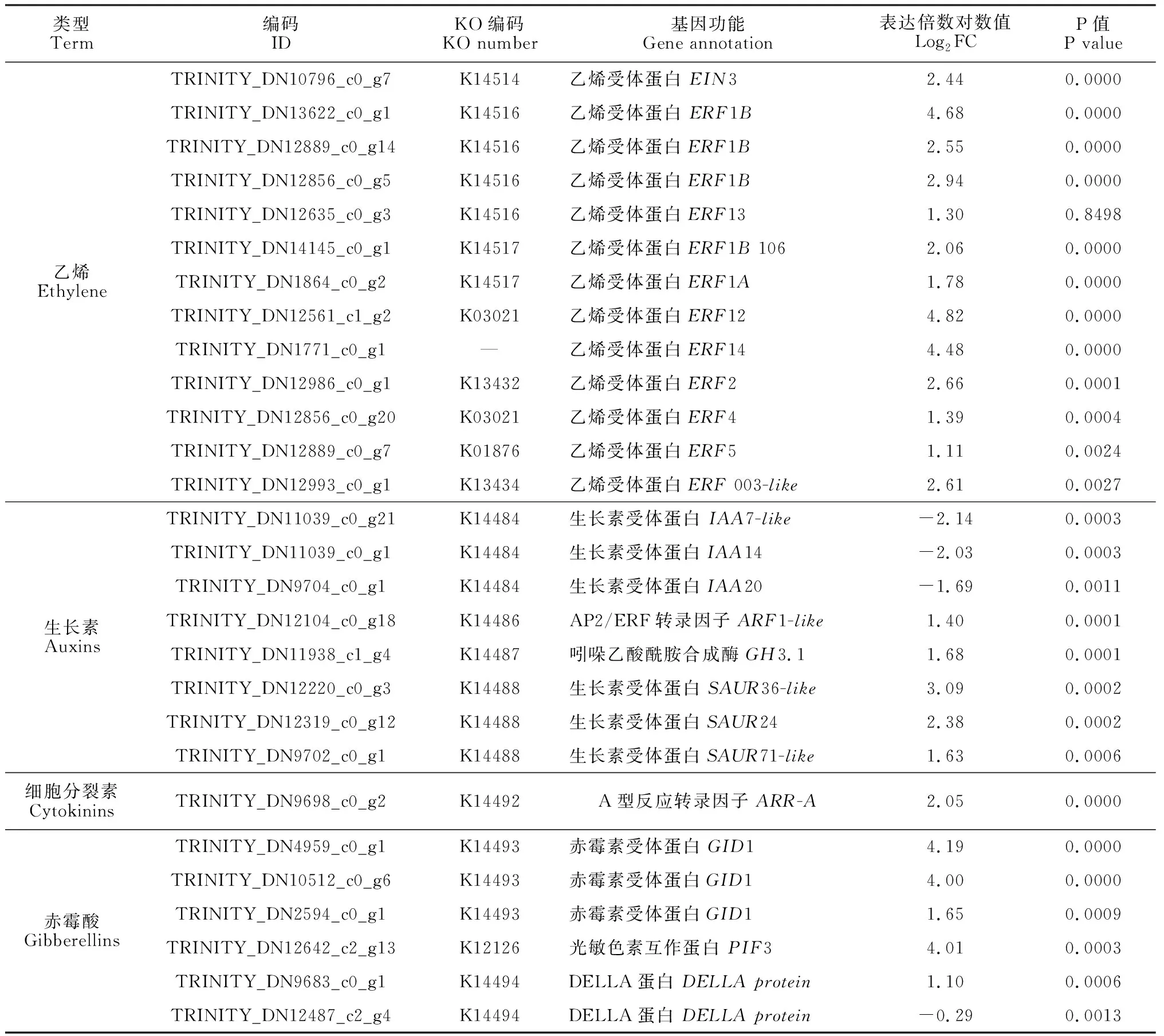
表5 與內源激素信號轉導相關的差異表達基因Table 5 Differentially expressed genes involved in phytohormone signal transduction
2.5 外源乙烯利處理苦瓜幼苗差異表達基因的qRT-PCR分析
為了驗證轉錄組測序的準確性,挑選了8個基因進行熒光定量表達分析,主要包括β-葡萄糖苷酶基因、細胞分裂素代謝調控基因CKX3、乙烯合成基因ACO1、細胞色素類基因CYP77A3等。結果顯示,上述8個基因的表達趨勢與轉錄組log2FC值變化趨勢一致,其中4個為上調表達,4個下調表達,表明本次轉錄組獲得的數據信息較為可靠(圖3)。
3 討 論
植物的生長發育受多種因素的調控,包括內因和外因的影響。外部因素主要體現為生長環境,包括溫度、光照、水分、土壤、養分等多個條件,組織器官的發育需要合適的環境條件。內部因素則主要以植物激素的形式扮演著極為重要的“調節器”作用,在極低的濃度下即可誘發植物產生顯著的生理反應,并表現為相應的農藝性狀改變。本研究中,外源乙烯利誘發了苦瓜幼苗幾種激素含量發生顯著變化。作為乙烯生物合成的直接前體物質及其生物合成限速酶,ACC的含量與ACS的活性均顯著升高。在番茄和大麥等作物幼苗中,施用乙烯利或者ACC均可以升高內源乙烯的含量,這與本研究結果一致[9,19]。生長素可以促進植株器官組織的發育,本研究中IAA的含量也表現出顯著的升高趨勢,陳學好等[14]的研究也表明,黃瓜幼苗經乙烯利處理后組織內IAA的含量出現了升高。Swarrp等[20]以擬南芥幼苗為試材,經外源乙烯處理后,IAA含量同樣表現為升高的趨勢。但也有一些作物在經過乙烯利處理后IAA的含量出現了下降,我們推測可能是由于實驗對象的不同或者取樣時期的差異所導致。細胞分裂素主要通過細胞數目的增殖和莖尖組織的快速分化調節植株的生長發育,其中兩種類型iP-type 和tZ-type通常被認為具有顯著生理活性[21],本研究中苦瓜幼苗經乙烯利處理后tZR的含量出現了顯著升高,表明外源乙烯與細胞分裂素之間存在著正向的互作關系。GA在植物的發育過程中主要調控植株的高度與種子的萌發,本研究中GA3的含量在乙烯利的作用下出現了顯著的下降,這與大多數研究結果相符[11-12]。孫義甫等[22]在早前的研究中曾指出,乙烯利與GA間存在著明顯的抗拮作用,適當濃度的GA溶液噴施黃瓜幼苗可以有效逆轉乙烯利對幼苗形態的影響。
內源激素含量的改變為進一步開展相關研究提供了條件,結合激素代謝的變化趨勢,我們對乙烯利處理后的苦瓜幼苗進行了Illumina測序等后續研究。以往關于苦瓜的研究報道,對于乙烯利的作用模式基本局限于生理水平的研究,并沒有深入探討其對分子調控網絡的影響,因此,從分子層面認識苦瓜應答外界乙烯刺激的研究具有重要理論意義。測序結果顯示,與激素代謝相關的功能基因出現了差異表達。首先在內源乙烯的代謝過程中,S-腺苷-L-蛋氨酸(SAM)是其生物合成途徑的重要起始物質,同時也是合成ACC的前體物質,本研究發現了3個表達水平顯著升高的HMT3基因,該類基因可以有效促進SAM的生物合成[23]。此外,ACO和ACS是乙烯合成途徑中的重要限速酶,二者的表達豐度在處理組中均出現了較大的提升,尤其是ACO1基因的表達倍數很高。上述結果表明,內源乙烯含量的升高主要源于SAM的累積以及ACO和ACS基因的正向調控。
對于植株的生長發育,生長素與細胞分裂素主要通過細胞體積的增大與細胞數目的增殖兩種方式進行調控。生長素的生物合成包括IAOx、TRA、IAM和IPA四條途徑[24]。本研究中2個與IAOx途徑相關的基因ALDH和TAA1出現了下調表達,而與IPA途徑有關的基因UGT74B1卻出現了上調表達。聯想到之前實驗結果中IAA含量的增加,推測在外源乙烯的處理下苦瓜幼苗IAA的合成主要集中于IPA途徑,且合成水平有較大的增強。對于細胞分裂素的變化,理化分析與轉錄組測序并不一致,HPLC-MS/MS檢測結果顯示tZR的含量比對照組有顯著升高,但tZR合成的限速酶基因CYP735A表達水平卻出現了下降。同時,促進tZR轉化為其他代謝產物的CKX基因卻表現為顯著的上調表達。對于上述研究結果,我們分析可能與tZR較特殊的合成過程有關。已有研究證實tZR主要在植物的根中合成,然后經由木質部運輸至地上葉片中[21]。在對地上植物組織進行乙烯利處理后,可能誘發了某種反饋機制,當葉片中與tZR生物合成相關的基因表達受抑時,根部tZR的合成代謝出現強化并更加集中地轉移至葉片中,來彌補由于轉錄受抑帶來的不利影響,從而使葉片部位保持較高的tZR含量。
赤霉素屬于四環二萜類化合物,迄今為止已發現一百多種類型。其中主要以GA1、GA3、GA4、GA7等結構類型具有顯著的生理活性,尤其以GA3的研究和應用最為廣泛[25]。外源乙烯對內源GA的影響較多表現為抗拮作用,本研究也發現苦瓜幼苗中GA3含量在乙烯利處理后出現了顯著下降,降幅達到了88.7%。但測序結果顯示,控制GA3合成的相關功能基因并沒有出現顯著的下調表達。值得注意的是,在激素信號轉導途徑中,多達11個ERF家族轉錄因子出現了表達水平的升高,其中有多個表現出極高的表達倍數;Dubois等[26]和Plett等[27]的研究發現,ERFs基因的上調表達可以誘發生理活性型GAs含量下降,進而導致植株表現出明顯的矮化生長現象。另外,我們在統計高倍數表達的差異基因時,發現了2個與GAs代謝間接相關的P450CYP736A12基因,前人研究表明在擬南芥幼苗中超表達這類基因可以使生理活性型GAs的含量出現下降,從而引發植株莖節的短縮[28]。鑒于此,我們推測本研究中外源乙烯利雖然沒有在轉錄水平影響GA3的生物合成,但轉錄后調控機制(Post-translational regulation)很可能以諸如ERF等轉錄因子介導的方式,使活性GA3出現分解,進而導致其含量的下降。
除上述參與內源激素合成的基因受到影響之外,激素信號轉導的相關基因也出現了表達水平的改變。植株在經歷乙烯利處理后,內源激素誘導信號通路中一系列調節因子差異表達,使植株發育過程中的細胞體積、數目,莖桿長度,以及側根發育等表觀性狀出現變化。這些調節因子中不乏有類似ERFs的基因,還能以反饋調控的方式影響內源激素(如GA3)的代謝水平。總體來講,本研究從分子代謝水平上獲得了大量外源乙烯利對苦瓜作物的作用靶點,并探討了基因可能存在的調控功能。將來可以從中挑選某些感興趣的基因進一步開展功能驗證等后續工作,從而更加深刻地揭示乙烯類物質對瓜類作物的調節機制。
