克隆和有性親體效應及其調控機制
2022-09-13 08:38:56羅芳麗王沫竹董必成陳禹含于飛海
生態學報 2022年16期
羅芳麗,王沫竹,2,董必成,陳禹含,于飛海,3,*
1 北京林業大學生態與自然保護學院, 北京 100083 2 中國科學院植物研究所系統與進化植物學國家重點實驗室, 北京 100093 3 臺州學院濕地生態學與克隆生態學研究所/植物進化生態學與保護浙江省重點實驗室, 臺州 318000
1 親體效應
植物個體的表型不僅取決于該個體的基因型(遺傳信息)和所處的環境,同時也可能受其親代個體(即親體)所處環境的影響[1—4]。這種親體環境對子代個體表型的修飾效應被稱為親體效應(也稱親代效應、親系效應或親本效應),包括母體效應(也稱母代效應、母系效應或母本效應)和父體效應(paternal effect,也稱父代效應、父系效應或父本效應)。由于親體效應反映的是子代個體表型對親體環境的響應模式,因此,親體效應有時也被稱為跨代可塑性[3,5]。
生物因素(如采食、競爭、植物激素等)和非生物因素(如養分、水分、光照等)均可誘導親體效應的產生[6—7]。早在1909年,植物的親體效應就被德國學者發現[4],但這種效應最初被認為是母體的“環境噪音”,會對子代個體造成不利影響[8]。近年來的進化生態學研究發現,在很多情況下,親體效應是可遺傳的,并可誘導子代個體進行適應于母體環境的表型特征調整,從而提高子代個體在與母體環境相似的環境下的行為、生態位寬度、進化潛勢以及入侵能力等[9—11],進而影響植物種群的結構、動態、進化方向和進化速率等,因而具有重要的生態和進化意義[3,5,12—13]。親體效應在非克隆植物中已被廣泛報導[3,13—16],在克隆植物中也開始受到越來越多的關注[17—21]。
1.1 有性親體效應
母體植株可通過有性繁殖過程(如產生種子)將母體環境的影響傳遞給子代植株,并影響子代植株的表型(圖1);這種通過有性繁殖傳遞的親體效應,稱之為有性親體效應。有性親體效應中的子代是指通過兩性細胞的融合形成的受精卵發育成的新個體[22]。由于種子通常與母體植株密切相關,因此,植物的這種親體效應常常表現為母體效應[7]。自然界中所有的非克隆植物和大多數克隆植物都具有性繁殖能力[22—23]。因此,有性親體效應在非克隆植物和克隆植物中均廣泛存在[2,24]。
越來越多的研究表明,母體環境不僅可以影響子代個體的表型,而且可以影響子代個體表型對環境變化的響應。例如,母體植株所經歷的土壤養分水平可以顯著影響其種子的萌發率、子代個體的株高和葉生物量[24—25],母體植株所經歷的光照脅迫可以影響其種子的萌發率和萌發時間[1],母體植株所經歷的昆蟲采食脅迫可以影響其種子重量、莖生物量和根冠比[17,26]。此外,母體效應引起的表型變異能顯著提高子代植株對脅迫環境的適應性[8,10]。例如,母體效應在子代植株適應養分脅迫[27]、光照脅迫[28—29]、采食[26]、水分脅迫[30]、鹽分脅迫[31]以及病原體感染[32]中均起著積極的作用。母體效應還能通過影響次生代謝產物(如茉莉酸和水楊酸等)的產生來提高入侵植物對本地天敵的抵御能力,從而顯著提高在入侵地的入侵性[33]。一些研究發現,母體效應在低資源環境(如土壤養分貧瘠、干旱和低光照等)下對子代適合度的影響較高資源環境下更為顯著[13—14]。也有研究表明,母體效應會隨著作用時間的延長而逐漸減弱,或者被子代對其所在環境較強的表型響應而掩蓋[34]。
1.2 克隆親體效應
克隆植物是指具有在自然條件下自發地產生遺傳結構相同并具有潛在獨立性新單元或個體的能力或習性的植物,它包括克隆生長和克隆生殖兩種主要繁殖方式[23]。克隆生長是高等植物最常見的無性繁殖方式[35]。克隆植物通過克隆生長所產生子代個體(即子代分株,簡稱子株)的基因型與親體(即親代分株,簡稱母株,mother ramet)完全一致,且能夠獨立于親體而生存[23]。子株直接來源于其親體的營養繁殖體,其在發育早期可能已對親體環境產生了預適應,同時,親體環境也可能是其發育后期所要經歷的環境。因此,子株生長發育與其親體環境的關系密切[7,17]。
克隆植物通過無性繁殖過程(如匍匐莖/根狀莖產生新個體)將親體環境的影響傳遞給子株,并與子株的基因型和所處環境共同影響子株的表型(圖1)。通過無性繁殖傳遞的親體效應,稱之為克隆親體效應。親體環境可影響母株營養繁殖體(如莖、根和根狀莖等)的營養儲備、代謝物質以及表觀遺傳變異等,進而影響子代表型[5,7]。與有性親體效應不同,由于營養生長的親體無母體和父體之分,因此克隆親體效應也無母體效應與父體效應之分。此外,由于營養生長形成的子代往往包含多個分株,分株的數量和大小之間往往存在權衡[36]。因此,克隆親體效應在子株分株和整株水平上可能是不同的,即當該效應對分株生長具有積極效應時,并不能表明對子株整株的生長也具有促進作用,反之亦然[37]。
已有研究表明,當子株經歷與親體相似的環境時,克隆親體效應能顯著提高子株的總生物量和平均生物量[12]。例如,與來源于非干旱親體的子株相比,受干旱脅迫的親體所產生的子株表現出更強的抗旱性[5]。然而,也有研究發現克隆親體效應對子株生長的影響不顯著,這可能與該親代環境作用時間的長短、親代植株發育階段以及子代環境作用的方向和強度等有關[34]。因此,克隆親體效應對克隆植物種群,尤其對有性繁殖受限和/或缺乏遺傳變異的克隆植物種群的長期進化過程起著非常重要的作用[38—39]。盡管目前國內外已經開展了一些針對克隆植物親體效應的研究,但這些研究絕大多數是在個體水平進行的[17,21],而有關親體效應在植物種群和群落水平作用的研究仍十分缺乏(圖1)。
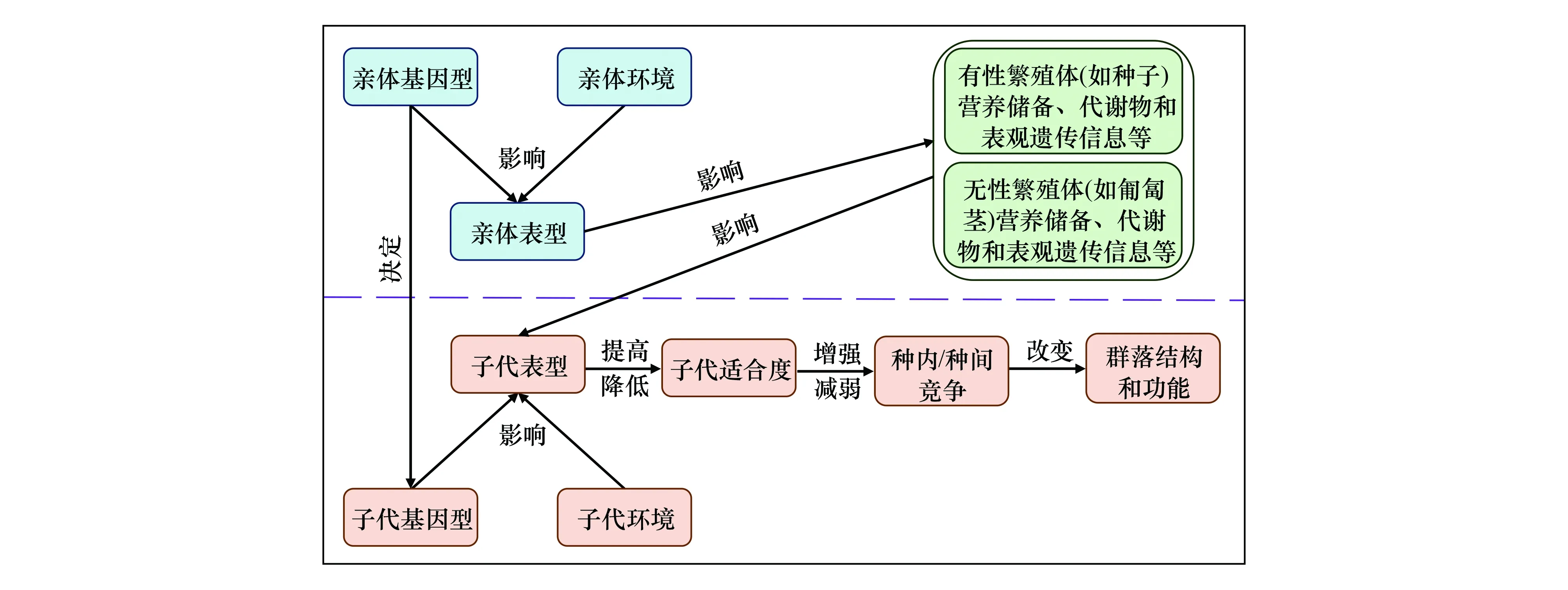
圖1 克隆和有性親體效應在植物子代個體、種群和群落水平的作用Fig.1 Roles of clonal and sexual parental effects at individual, population and community levels of offspring plants

圖2 與子代環境無關聯或有關聯的親體環境效應示意圖[37]Fig.2 Schematic diagram for parental environmental effects that are not related to offspring environments or that are related to offspring environments [37]
1.3 親體效應對后代表型和適合度的影響
Dong等[37]認為,親體效應對后代表型和適合度的影響可能包括以下四種類型(圖2)。第一種類型為親體效應不依賴于后代所處的環境, 即親體在良好環境條件下產生的后代個體總是比在不良環境條件下產生的后代個體的適合度高,或良好環境條件下親體產生的后代個體的表型對后代個體的適合度提高總是有利的[40—42]。第二種類型為親體效應在后代環境與母代環境相似時是有利的,而在與母代環境不同時是不利的[1,43]。第三種類型為親體效應只有在后代處于良好環境條件下才有利,而在后代處于不良環境條件下沒有顯著影響[10]。第四種類型為親體效應只有在后代處于不良環境條件下才有利,而在后代處于良好環境條件下沒有影響[44]。
1.3.1有性親體效應對后代表型和適合度的影響
一些研究表明,當子代個體經歷與母體相似的環境條件時,有性親體效應往往能顯著提高子代個體對這種環境的適應性,如顯著提高子代植株的個體大小、碳水化合物積累、生物量、繁殖產出和幼年植株的存活率[10,25,30],提前植物的花期[45]以及提高種間競爭能力[25]等。相反,當子代環境與母代環境不一致時,親體效應可能不顯著,甚至可能表現出負效應,如延遲繁殖期、降低繁殖產出等[14]。因此,親體效應可以對子代適合度產生積極或消極的影響,這與母代環境密切相關[14]。同時,有研究發現有性親體效應可以在代際間進行疊加,例如兩代(祖母代和母代)與一代(母代)處于干旱脅迫環境相比,其子代具有更高的營養物質含量、更快的萌發過程、更高的存活率和更大的植株個體,表明親體效應在代際間的疊加能讓子代提前適應相似的環境脅迫,從而顯著提高其適合度[30]。親體效應對子代表型和適合度的影響可能進一步影響植物的種內[31]和種間關系[14,25],進而影響植物種群和群落的結構、功能和穩定性(圖1)。
1.3.2克隆親體效應對后代表型和適合度的影響
克隆親體效應可以通過親體產生的營養繁殖體傳遞至子代,在一定條件下有助于提高克隆植物對環境變化(如資源異質性)的適應能力(圖1)。與有性親體效應一致,在克隆親體效應中子代適合度受親代和子代環境的共同影響;不同的是,克隆親體效應同時也受營養繁殖體類型、營養物質水平和代謝物質水平等的影響[7,17,46—47]。在克隆親體效應的研究中,平均適合度和總適合度為常用指標,它們分別以同一母株產生的所有分株的適合度指標(如分株數和/或生物量)的平均值或總和來衡量[37]。當親代與子代均處于高養分環境時,克隆親體效應能顯著提高克隆植物空心蓮子草(Alternantheraphiloxeroides)子代的平均適合度和總適合度[17,37,46]。同時,克隆親體效應也可顯著提高克隆植物鳳眼蓮(Eichhorniacrassipes)和空心蓮子草等的入侵性[48—49]。已有研究表明,不同克隆繁殖體類型的克隆親體效應強度不同,例如,養分處理下空心蓮子草通過母代莖形成子代的親體效應更強,而在采食脅迫下根形成子代的親體效應更顯著[17]。然而,對該物種親體效應的研究也發現其某些表觀遺傳變異可持續十個無性世代,但并非長期穩定,而是表現出中等水平的穩定性[49]。這種暫時的表觀遺傳穩定性可能是克隆植物(特別是遺傳變異受限的植物)適應未知生境的重要機制[48,50]。
1.4 親體效應對后代競爭能力的影響
1.4.1有性親體效應對后代種內和種間競爭能力的影響
母體可通過影響其種子的營養物質儲備來改變子代個體的表型可塑性,從而影響子代個體的競爭能力(圖1)。資源獲取能力強的母體通常能產生高質量的種子,使子代個體具有較強的競爭能力[2,13,51];同時,在子代個體發育早期,良好的資源條件也能提高其成株的表型性狀、生物量積累和競爭能力[40,52]。此外,母代植株還可通過表觀遺傳變異(詳見2.3)影響子代植株的表型可塑性,從而改變子代的競爭力[3,51,53—54]。相反,當種子的營養儲備少、子代早期資源狀況差或子代環境不可預測時,母體效應可能不會顯著影響甚至會抑制子代個體表型性狀,從而降低子代個體的種內和種間競爭能力[37]。由于母體環境、物種或基因型的差異,種子的營養儲備往往不同[8,53]。因此,相同或不同植株間可能會產生表型性狀的分化,改變對資源利用的有效性,從而影響植物種群或群落內的種內和種間競爭強度。
已有研究較多地關注非生物因子脅迫下有性親體效應對子代競爭能力的影響[13—14],然而,自然生態系統中往往同時存在非生物因子和生物因子脅迫的影響。比如,母代植物種內/種間競爭(即生物因子脅迫)或者母代非生物與生物因子脅迫的交互效應(如競爭 × 土壤水分)所引起的表型可塑性也能進行跨代遺傳[3,51]。當母體經歷較強的種內/種間競爭環境時,其子代個體也具有較強的競爭能力,表現在種子具有較快的萌發過程、植株具有較快的生長速率和較高的表型可塑性等[3,51]。
1.4.2克隆親體效應對后代種內和種間競爭能力的潛在影響
如果克隆親體效應對后代個體表型的改變能提高其生長和適合度[17,20,37,55],那么克隆親體效應可能增強植物后代個體的種內和種間競爭能力(圖1)。相反,如果克隆親體效應對植物后代個體表型的改變在某些情況下降低其生長和適合度[1,10,14],那么克隆親體效應將削弱后代個體的種內和種間競爭能力[25]。然而,目前國內外有關克隆親體效應對植物種內和種間關系影響的研究仍十分缺乏[14,25],已開展的少數幾例研究均針對非克隆植物,探討其有性親體效應的影響[14,25,31,53,56]。克隆親體效應可能通過改變后代尤其是群落優勢種的種內和種間競爭能力,從而影響群落的組成和多樣性。目前,有關克隆親體效應對植物種內和種間關系影響的研究還有待開展(圖1)。
1.5 親體效應對種群結構、功能和穩定性的潛在影響
植物親代個體所經歷環境的多樣性可通過親體效應使后代個體產生表型的多樣性[17,37,41,54],而這種親體效應介導的表型多樣性對植物種群的結構、功能和穩定性將可能產生重要的影響(圖1)。大量研究表明,生物多樣性的提高對生態系統功能(如生產力)和穩定性(如抗干擾能力)具有重要的促進作用,這種效應不僅體現在物種(多樣性)水平[57—58],而且也體現在種內(遺傳多樣性)水平[59—60]。物種多樣性和遺傳多樣性對生態系統功能和穩定性的促進效應主要是由于物種多樣性高的群落或遺傳多樣性高的種群包含更多對資源獲取策略不同的物種或基因型,即包含更多功能差異的表型,從而能夠更加充分的利用群落或種群中的資源(互補效應)[61],或者由于這些群落或種群包含對其功能影響較大的優勢物種或基因型,即包含具有優勢功能的表型(選擇效應)[61]。物種多樣性和遺傳多樣性介導的種間和種內表型多樣性均涉及非常高的遺傳變異,而克隆和有性親體效應介導的種內表型多樣性可以完全不涉及或僅涉及少量的遺傳變異[17,37,41]。與物種多樣性和種群遺傳多樣性的效果類似,如果親體效應介導的表型多樣性高的種群包含更多功能差異的表型或包含具有優勢功能的表型,那么通過互補效應和/或選擇效應,該種群的功能和穩定性可以得到顯著提高。然而,迄今為止,國內外尚未開展有關親體效應介導的表型多樣性對種群結構、功能和穩定性影響的研究(圖1)。
1.6 親體效應對群落結構、功能和穩定性的潛在影響
對于植物群落中的優勢物種,有性親體效應介導的后代表型多樣性也可能進一步影響植物群落的組成、功能和穩定性[62—64]。與群落中優勢種遺傳多樣性介導的表型多樣性在群落水平的效應相似,如果群落中優勢種親體效應介導的后代不同表型之間具有互補的資源利用能力,那么后代表型多樣性的增加可能提高該種群的資源利用效率和生產力,從而使群落中其它物種由于難以獲取到必需的資源而衰退[54]。在這種情況下,群落中優勢種親體效應介導的后代表型多樣性將降低群落的物種多樣性和生產力。相反,如果群落中優勢種親體效應介導的不同表型后代個體之間在競爭、促進或化感等能力上存在顯著差異,即具有功能差異,使得群落中其它物種可以與該優勢種不同表型的個體共存,那么群落優勢植物親體效應介導的表型多樣性可提高整個群落的物種多樣性和生產力[54]。然而,國內外尚缺乏有關親體效應介導的表型多樣性對群落結構、功能和穩定性影響的研究(圖1)。
2 親體效應的潛在調控機制
親體效應受繁殖體營養儲備、代謝物質水平、表觀遺傳變異等眾多機制的調控[5]。不同調控機制下,親體效應作用的歷時長短不同。與表觀遺傳機制相比,供給機制和代謝物質調控機制對后代影響的歷時相對較短[7]。在自然種群中,這些調控機制并不能嚴格地區分開,也不會相互排斥,往往是多個機制共同影響后代表型[8]。
2.1 供給機制
養分供給效應是指親代植株將碳水化合物、脂質、蛋白質和礦質營養等分配給繁殖體,通過繁殖體將營養物質提供給子代[8,51]。養分供給效應在自然界普遍存在,這在早期的研究中已被證實[4]。種子和營養繁殖體的營養儲備被認為是關鍵的從親體獲取的性狀,通常與繁殖體數量存在權衡關系[8]。營養物質的供給量取決于親體所處環境的資源狀況和親體的營養分配策略,同時也受植物遺傳因素和環境因素(如干旱、高溫、遮陰、放牧等)的影響[51]。在相同環境條件下,不同物種或同一物種的不同基因型個體的母體細胞質等也可影響種子的營養物質儲備[8]。養分供給效應不僅影響營養繁殖芽的萌發、種子的萌發和子代幼苗存活率,同時對子代生長、競爭能力和繁殖等也具有顯著的影響[20,37,65—66]。
親代較高的繁殖體分配還可能產生銀勺效應(也稱后續效應),即營養儲備較高的繁殖體形成的后代個體無論其生長環境條件如何,總是具有較高的適合度[40,67]。因此,銀勺效應能提高子代個體后期在脅迫環境下的表型可塑性、生物量積累、繁殖能力以及適合度[67—68]。近期關于克隆植物空心蓮子草親體效應的研究表明銀勺效應顯著提高了子代植株后期在低土壤養分條件下的生物量積累和營養繁殖能力[21]。由于該效應強調個體發育早期良好的環境條件對發育后期耐受性和適應性的影響,有研究指出該效應對子代的影響往往具有滯后效應,例如,子代個體發育至成株后才表現出較強的競爭力[52,69]。
子代個體的適合度也能反映親體環境的營養狀況:養分狀況良好生境的親體所產生的繁殖體具有較高的營養儲備,能為子代個體提供更多的營養物質,產生的子代適合度較高;相反,親代植株養分供給不足時,繁殖體營養儲備較少,供給效應減弱,子代個體的適合度也相應降低[5,70]。研究表明,種子的養分供給效應對于幼苗早期適應脅迫環境具有重要作用。例如,母體養分供給充足的子代植株能形成龐大的根系以適應干旱土壤,或形成高大的枝干以適應遮陰環境[71]。然而,在自然種群中,養分供給效應也存在一定的局限性。首先,養分供給效應對植物發育早期的表型可塑性影響較為顯著,隨后逐漸減弱[3,24]。其次,養分供給不足時,種子的產量和質量均不高,供給效應較弱。最后,由于較大的種子具有較強的競爭力和耐受能力,而較小的種子在種子庫中具有較長的存活率,因此,種子養分供給往往還與土壤種子庫維持存在權衡關系[8]。
2.2 代謝物質調控機制
植物繁殖體不僅能從親體獲取營養物質,同時也可獲得一些代謝物質,如蛋白質、mRNAs、防御化合物、次生代謝產物和激素等[8]。其中,蛋白質既可作為供給機制中的營養物質,也可以作為調控分子起作用。研究表明,蛋白質和mRNAs在調控種子休眠和萌發過程中起著重要的作用[72]。親體環境能改變其基因表達,繁殖體中的mRNAs能將遺傳信息進行準確轉錄并合成蛋白,有助于脅迫條件下種子的萌發[73—74]。母體環境脅迫還可以改變種子中激素的含量及胚對激素的敏感性;例如,與非遮陰條件下相比,長芒莧(Amaranthuspalmeri)母體在遮陰條件下產生的種子中脫落酸的含量提高了44%[75]。種子萌發常常受多種調控因子如激素水平、蛋白質、mRNAs等的共同作用,但這些調控因子對種子萌發和子代幼苗發育影響的相對重要性以及相關機制目前尚不明確[8,72]。
2.3 表觀遺傳機制
親體的環境條件可以誘導其產生表觀遺傳變異,并且這種表觀遺傳變異可能遺傳給后代,從而引起后代個體表型的變異,因此,表觀遺傳變異可能是親體效應的潛在機制之一[3,76]。
2.3.1表觀遺傳變異
表觀遺傳變異基于一系列可激活、降低或完全消除某些基因表達的分子過程,包括DNA甲基化、染色質結構重塑、以及小RNA分子介導的調控過程等;這些分子機制可調控基因表達,進而改變植物個體的表型[77—78]。其中,DNA甲基化廣泛存在于植物體中,它主要來源于遺傳控制、環境誘導及自發產生的表觀突變,被認為是植物親體效應的重要調控機制之一[9,79]。
DNA甲基化是由DNA甲基轉移酶(DNA methyltransferase,DNMT)催化S-腺苷甲硫氨酸(S-adenosyl-Lmethionine, SAM)作為甲基供體,將甲基基團加在CpG島上(CCGG),從而將胞嘧啶轉變為5-甲基胞嘧啶的一種反應[77,80]。DNA甲基化廣泛存在于細菌、真菌及動植物體中。植物基因組的甲基化常發生在對稱的CG或CHG位點,以及非對稱CHH位點上(H = A,C,T),并參與調控轉錄抑制[81]。首先,基因調控區域的胞嘧啶甲基化可直接干擾轉錄因子與其識別位點的結合,進而抑制基因表達[80—81]。其次,一些結合蛋白也可特異性地結合到甲基化位點上,并通過募集組蛋白去乙酰化酶形成競爭性轉錄阻遏物抑制轉錄因子的結合[80—81]。最后,DNA甲基化還可通過染色質重塑影響基因轉錄與表達[82—83]。因此,DNA甲基化可通過調控基因表達及與轉錄機制的相互作用影響植物表型。
已有研究表明,環境誘導的親代表觀遺傳變異至少在某些情況下可以直接遺傳給子代個體,并影響其對環境變化的表型響應[49,84—85]。例如,親代經歷土壤養分、水分、溫度和鹽分等處理介導的表觀遺傳變異可以顯著影響植物種子的萌發以及后代個體的生長和抵御脅迫的能力[85—86];親代經歷的光照條件誘導的DNA甲基化可以顯著影響空心蓮子草后代個體的生活史性狀[18];親代遭受采食脅迫所誘導的表觀遺傳變異也可以影響野蘿卜(Raphanusraphanistrum)有性后代個體對采食的防御能力[26]。因此,表觀遺傳變異給植物提供一種新的快速應對環境變化的途徑,使得某些性狀的快速進化成為可能[77,87]。
2.3.2遺傳變異、表觀遺傳變異與表型變異
遺傳變異是指DNA序列的變化[59,88],而表觀遺傳變異是在沒有DNA序列改變的情況下基因表達和基因功能的可遺傳變化[89—90]。表觀遺傳變異受遺傳變異的調控,但很多情況下表觀遺傳變異可以部分的、甚至完全的獨立于遺傳變異[91—92]。一些植物種群的遺傳變異與表觀遺傳變異之間存在顯著相關性,如擬南芥(Arabidopsisthaliana)、西班牙紫羅蘭(Violacazorlensis)、互花米草(Spartinaalterniflora)、短芒大麥草(Hordeumbrevisubulatum)和水稻等,表明表觀遺傳變異可由遺傳變異所驅動,且兩者共同影響植物的表型變異[93—98]。表觀遺傳變異和遺傳變異兩者間的相關性可能是由順式(cis)或反式(trans)遺傳信息促使特定位點表觀遺傳印記的獲得或保留所引起的[99]。然而,也有一些植物種群的表觀遺傳變異與遺傳變異之間無顯著相關性,如香菇草和野生馬鈴薯(Solanumruiz-lealii)種群可在遺傳變異水平較低的情況下,表現出較高的表觀遺傳變異[81,92,100—102]。表觀遺傳變異的獨立性可能是由于減數分裂或突變過程中發生了表觀遺傳信息的隨機漂變[78,103]。種群表觀遺傳變異與遺傳變異的關系對于研究種群進化具有重要意義[104];因此,更多相關的研究亟待開展。
2.3.3親體效應與表觀遺傳變異
(1) 有性親體效應的表觀遺傳機制
DNA甲基化可在有性繁殖的減數分裂過程中進行保守復制并穩定傳遞給子代[5,105]。在DNA復制過程中,特異的甲基轉移酶可識別新合成的半甲基化雙鏈,并基于鏈對稱性將甲基加在新鏈的非甲基化胞嘧啶上,從而復制甲基化模式,實現DNA甲基化的跨代遺傳[46,106]。因此,表觀遺傳可使植物“記住”所經歷的環境,預測并克服未來的環境壓力,并通過多代的可靠傳遞,選擇適應性信息[81,107]。然而,表觀遺傳信息在減數分裂過程中可能會發生DNA甲基化重置的現象,阻礙表觀遺傳信息在世代間的傳遞,從而削弱親體效應[105,108—109]。
有性親體效應下DNA甲基化具有不同程度的遺傳穩定性:短暫變異,短期可遺傳變異(兩代間遺傳),以及長期可遺傳變異(多代間遺傳)[81,110]。盡管可遺傳的表觀遺傳變異被認為是親體效應的重要調控機制,但表觀遺傳的穩定性及其與親體效應的關系尚不明確,直接驗證兩者關系的研究仍非常缺乏[5,46]。
(2) 克隆親體效應的表觀遺傳機制
對于克隆植物的親體效應,由于營養繁殖缺少減數分裂重組及分離過程中的變異發生機制,因此,受母代環境影響的表觀遺傳信息能更有效地傳遞至新產生的子代,并成為可遺傳表型變異的重要來源[46,49,111]。例如,當克隆植物香菇草的母代和子代均處于遮陰條件時,DNA甲基化變異的遺傳率可達到94.44%[102]。環境脅迫的強度及持續時間可影響克隆親體效應,在多代間重復相同的環境脅迫有利于克隆親體效應的維持[12,109]。例如,與來源于非干旱脅迫的子代相比,受干旱脅迫的母代所產生的子代表現出更強的耐旱性[5]。因此,表觀遺傳變異對克隆植物種群,特別是有性繁殖受限、缺乏遺傳變異的克隆植物種群尤為重要,提高了這些植物對環境變化的適應能力,在其長期進化過程中可能發揮著重要的作用[38—39]。然而,目前關于表觀遺傳變異在克隆親體效應中的作用尚不清楚[111]。
3 克隆植物、親體效應與表觀遺傳機制
克隆植物在自然界廣泛存在,是很多生態系統的優勢物種[23,112—113]。克隆植物特有的生活史性狀包括營養繁殖、克隆整合、克隆可塑性和克隆內分工等,這些性狀可以增加克隆植物抗逆、抗干擾的能力以及對異質性資源的利用能力[35,47]。研究表明,很多克隆植物如空心蓮子草、鳳眼蓮和虎杖(Fallopiajaponica)等種群的遺傳多樣性極低,僅由一個或幾個優勢基因型組成[23,48,50,114],但卻存在較高的表觀遺傳變異(表現為較高的DNA甲基化水平)。通過克隆親體效應,受母代環境影響的表觀遺傳信息能有效地傳遞給后代,并影響無性后代個體的表型[7,111],然而,它是否會影響無性后代個體的克隆生活史性狀和適合度目前尚不清楚。如果克隆親體效應能通過表觀遺傳變異影響后代個體的克隆生活史性狀,那么它將可以極大地提高克隆植物適應環境脅迫和環境異質性的能力(圖1)。
在自然生境中,克隆植物種群內和種群間表觀遺傳變異的大小及差異,以及表觀遺傳變異、遺傳變異、表型變異和環境變化之間的關系均是目前生態表觀遺傳學亟待解決的關鍵科學問題(圖1)。由于不同生境下的植物種群遺傳變異程度不同,自然植物種群遺傳變異和表觀遺傳變異對表型變異的貢獻程度較難區分開[91—92],然而,明確這些變異之間的關系是深入理解表觀遺傳變異影響克隆植物進化潛勢的前提和基礎[104]。在區分表觀遺傳變異和遺傳變異效應的研究中,通常選用同一種系的后代個體來控制遺傳變異,但不同個體間仍存在遺傳變異[26,115—116]。與非克隆植物相比,克隆植物遺傳變異程度相對較小,來自同一基因型基株的后代個體可以嚴格控制遺傳變異的潛在影響,因而易于區分遺傳變異與表觀遺傳變異的作用[9,102]。因而,克隆植物更有利于探究表觀遺傳變異在多種生態和進化過程中的功能意義[46,117]。
4 展望
親體效應對植物尤其是克隆植物的長期適應進化具有重要意義[38—39,47]。然而,個體水平上克隆親體效應對克隆后代特有生活史性狀的影響及其表觀遺傳機制尚不明確,種群和群落水平上克隆和有性親體效應的作用及其表觀遺傳機制的相關研究就更為缺乏。親體效應所介導的后代(尤其是優勢物種或基因型)的表型多樣性可能與物種或基因型多樣性具有相似的生態學效應[7,56,118—121]。由于不同表型之間可能具有資源利用互補、生態位分化或形成優勢表型的潛力,種群表型多樣性越高,其生態功能(如生產力積累或抗干擾能力)可能也越強。
基于前期相關基礎,研究認為仍有以下幾個重要問題需要進一步關注。在個體水平上:(1)克隆植物自發表觀突變的發生頻率及其遺傳穩定性如何?(2)遺傳變異與表觀遺傳變異調控克隆生活史性狀變異的貢獻程度如何?(3)克隆植物基因組特點、倍性水平、生活史特征、生境預測能力等是否會對表觀遺傳變異產生影響?在種群和群落水平上:(4)克隆和有性親體效應介導的表型變異是否影響植物的種內和種間關系?(5)克隆和有性親體效應介導的表型多樣性是否影響植物種群的結構和生產力?(6)克隆和有性親體效應介導的表型多樣性是否影響群落的組成和生產力?(7)植物野外種群表觀遺傳變異與其種群和群落特征之間是否存在相關關系?通過闡明克隆和有性親體效應在種群和群落水平的作用及表觀遺傳機制,以期能深入理解克隆和非克隆植物種群的適應與進化、群落的結構與動態以及生態系統的結構、功能和穩定性,進一步推動生態表觀遺傳學和植物生態學的發展。
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
中學物理·高中(2016年12期)2017-04-22 11:53:03
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
