利用Ecopath模型評價鰱鳙放養對千島湖生態系統的影響
2022-09-13 08:46:08鄭一琛常劍波
生態學報 2022年16期
鄧 悅,鄭一琛,常劍波,3,*
1 武漢大學水利水電學院, 武漢 430072 2 武漢大學水生態研究所, 武漢 430072 3 武漢大學水資源與水電工程科學國家重點實驗室, 武漢 430072
千島湖是錢塘江的重要水源,對保障錢塘江中下游的水環境質量以及周邊上千萬人口的飲用水安全起著非常重要的作用。然而隨著地區經濟的加速發展,千島湖的污染負荷迅速增長,于1998年和1999年爆發了兩次大面積的水華。自此后千島湖實施了以鰱、鱅放養和兇猛魚類控制為主要措施的“保水漁業”,目的是為通過下行效應遏制水華現象。但近年來千島湖局部水域仍不時爆發藍藻水華,可見生態系統在人工干預下的結構化進程尚未被真正了解。
Ecopath模型最早由Polovina在1984年提出[1],隨后結合Ulanowicz的能量分析生態學理論,發展成為一種生態系統能量流動的定量分析方法[2]。它基于物質平衡原理,可以對生態系統的食物網結構、能量流動過程和流動特征等進行量化綜合分析[3—4],目前已經成為水域生態系統研究的重要工具,廣泛應用于生態系統的食物網結構及能流特征研究[3,5—7],生態修復和漁業的影響評價[8—13]以及區域生態系統對比分析中[14—19]。本研究依據2008—2010年間在千島湖進行的生態和漁業資源調查數據,構建了千島湖生態系統2010年的Ecopath模型。通過分析2004—2016年間生態系統的基本結構和總體特征變化趨勢,總結在“保水漁業”的人工干預下的生態系統結構化過程,評價長期投放鰱、鳙魚對生態系統造成的影響。本研究工作從生態學原理上分析修復措施給淡水水體帶來的影響,是一次有益的探索和嘗試,可為后期類似水體生態修復評價提供數據支撐。
1 材料和方法
1.1 研究區域概況
千島湖位于浙江西部與安徽南部交界的淳安縣境內,地處29°11′—30°02′N,118°34′—119°15′E,是20世紀50年代為修建新安江水電站筑壩攔蓄新安江上游而形成的淡水湖泊。千島湖水域面積約為570 km2,東西長約60 km,南北寬約50 km,正常庫容量為178.4×108m3,最深處達97 m,平均水深34 m,多年平均水溫15.8℃。千島湖是飲用水重要水源地,其漁業也十分發達,是淳安縣的支柱產業。
1.2 Ecopath模型
1.2.1劃分功能組
功能組是指具有共同棲息地、相似食性、相似尺寸、相似生活史特征的物種集合。為使模型更具可比性,在本研究中,除依據各生物種類的食性、個體尺寸及棲息地特點等特性外,還參考了2004年的劃分標準,將千島湖生態系統劃分為17個功能組。由于千島湖生態系統種群結構的變動,2004年、2016年和2010年的功能組組成有所不同(表1)[12,20]。

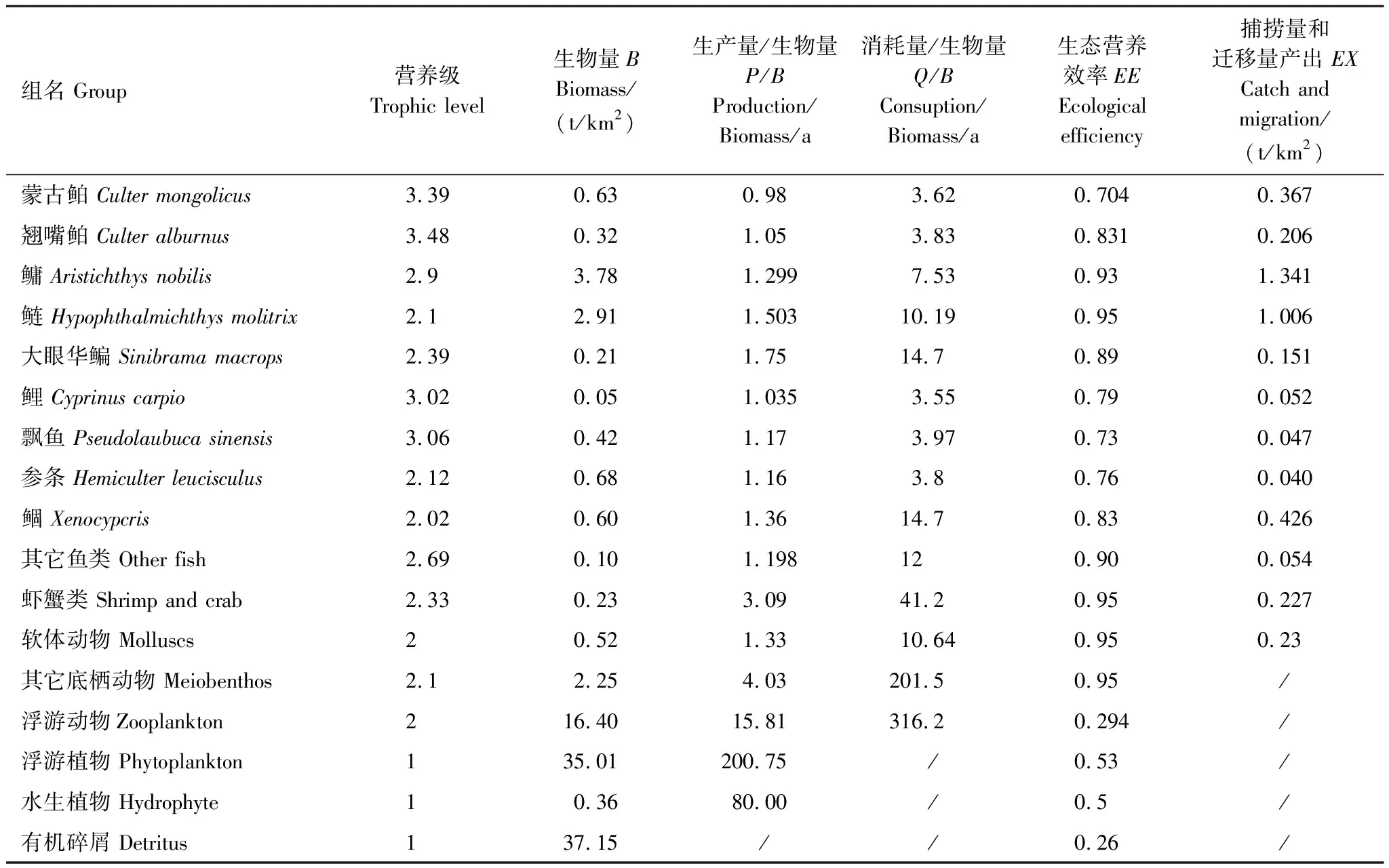
表1 千島湖生態系統3個年份的功能組
1.2.2生態學參數
各功能組的各類生態學參數的估計如下:
(1)生物量(B)
生物量在Ecopath模型中選用生物濕重(t/km2)的能量形式來表示。部分魚類的生物量數據由捕撈量數據計算而出,具體的計算方法為:
B=Y/F=Y/(Z-M)
(1)
式中,Y表示漁獲量,F為捕食死亡率,Z為總死亡系數,Z=P/B,M為自然死亡系數,M的數值由 Pauly. D在1980年提出的經驗公式估算出[21]:
logM=-0.2107-0.0824logW∞+0.6757logK+0.4627logT
(2)
logM=-0.0066-0.279logL∞+0.6543logK+0.4634logT
(3)
式中,L∞為漸進體長(cm),W∞為漸進體重(g),K為生長系數,T為該水域年平均水溫(℃),取15.8℃;
研究中其它底棲生物、浮游動物、浮游植物的生物量參考在2008—2010年的實測數據[22—24]。部分功能組的生物量因為無已知的生態調查數據,故采用預設的生態營養轉換效率EE算出。參照其它許多生態系統模型通行的方法[25], 將蝦蟹類、軟體動物、底棲藻類的EE值分別假定為 0.95、0.95和0.5。碎屑的生物量計算采用與初級生產碳相關的相關經驗公式[26]。
(2)生產量/生物量(P/B)
P/B系數指年生產量和年平均生物量的比值。魚類和蝦蟹類的P/B系數參考千島湖和竺山湖的相關文獻[12,27];軟體動物和其它底棲生物的P/B系數參考太湖的相關研究[28];浮游動植物以及底棲藻類的P/B系數引自千島湖相關文獻[12]。
(3)消耗量/生物量(Q/B)
Q/B系數為單位時間某種生物的消耗量與生物量的比值。研究中魚類的Q/B由Pauly的經驗公式計算[21]:
log(Q/B)=7.964-0.204logW∞-1.965T′+0.083A+0.532h+0.398d
(4)
T′=1000/(T+273.15)
(5)
A=H2/a
(6)
式中,W∞是漸進體重(g),T為該水域的表層水溫(℃),A為尾鰭形狀參數(一般魚類取1.32),h為布爾型變量(草食魚類取1,其它類取0),d為布爾型變量(碎屑食性的魚類取1,其它類取為0),H為魚類的尾鰭高度(cm),a則為尾鰭面積(cm2)。所需要的參數數據通過查詢漁業數據庫網站(http://www.fishbase.org),并參考相關的文獻[12,28—29]。
蝦類、軟體動物、其它底棲動物和浮游動物的Q/B由Q/B=(P/B)/(P/Q) 間接計算而來, 它們的P/Q的取值參照一些公認的資料[30],分別為0.075、0.125、0.02和0.05。
(4)捕撈量和遷移量產出(EX)
參考千島湖魚類資源數據中做的漁業捕撈系統調查得到,統計中額外考慮鰱、鳙魚每年的投放量,作遷入量計。
(5)生態營養效率(EE)
蝦、軟體動物、底棲藻類的EE值分別假定為 0.95、0.95和0.5[25],部分魚類功能組參考相關文獻來取值[12,31]。
(6)食物矩陣(DC)
魚類的食性組成參考千島湖和太湖的相關文獻[27,31];對包含幾種動物的功能組,食物組成則根據權重算出。浮游動物、軟體動物、水蚯蚓和蝦蟹類的食性參照目前的食性研究結果[26,32]。
1.2.3模型調試
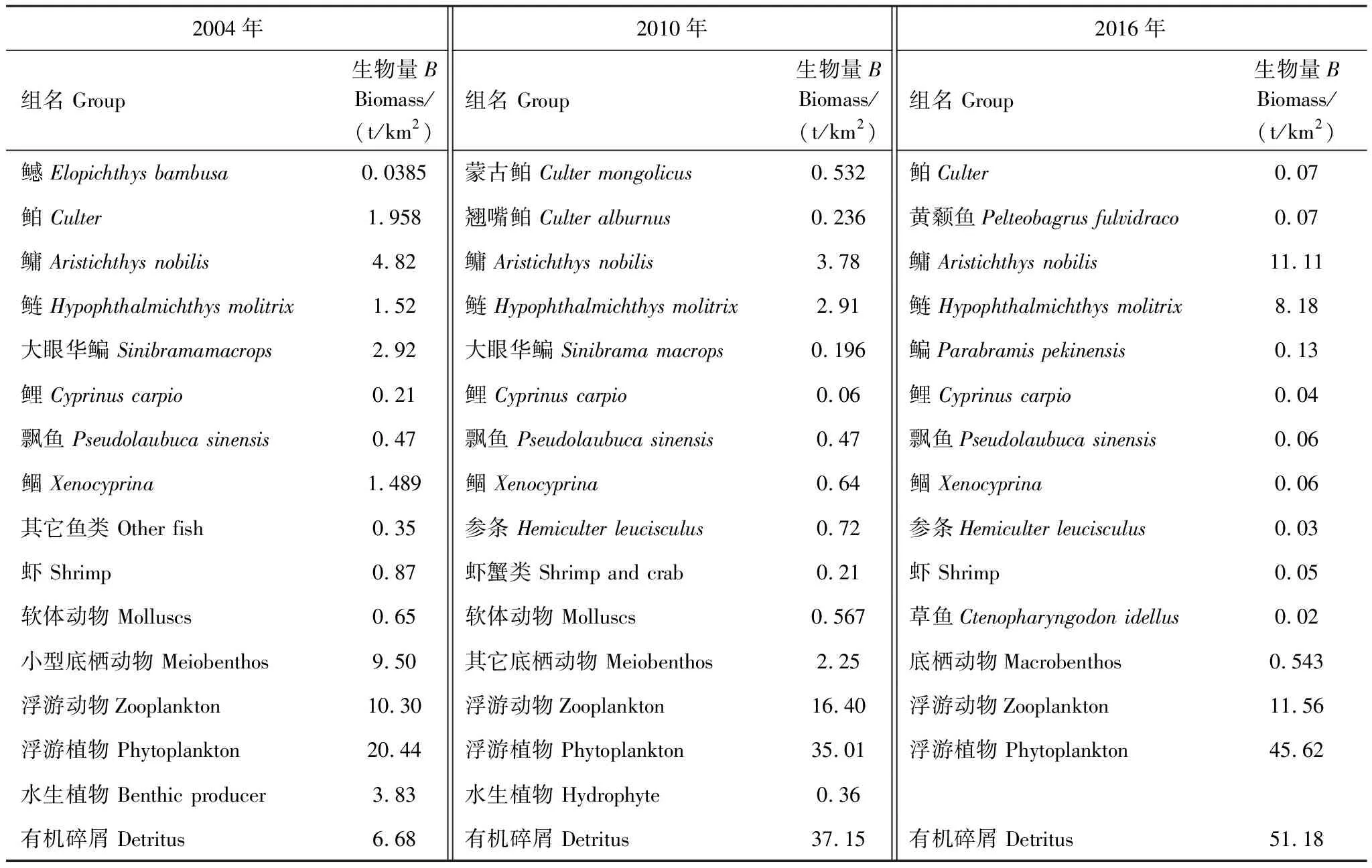
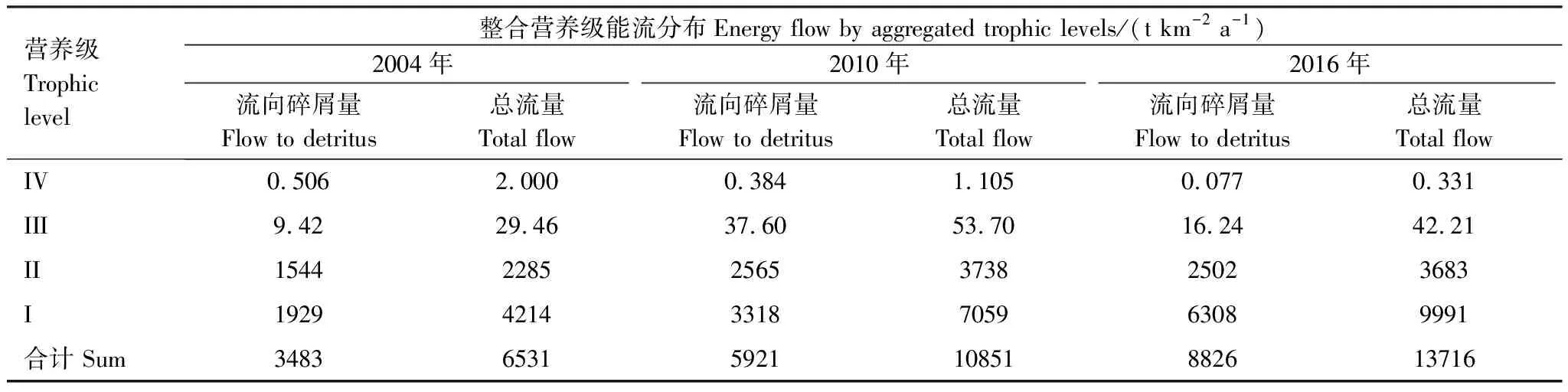
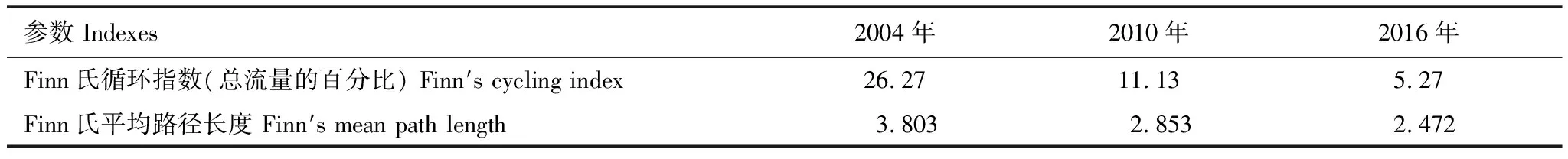
基本參數輸入模型后,需要保證各功能組的生態營養轉換效率EE值都要調整到在0和1之間。因為超過1的EE值是不正常的,任一個物種被捕食和被捕撈的量不可能大于其自身的能量產出[33—34]。這時需要通過調節部分參數,使模型中各個功能組的輸入和輸出達到平衡。模型在首次運行時出現有的功能組的EE值大于1,通過調整P/B或者DC等來使0 參數輸入模型后,進行模型的平衡與計算,輸出千島湖生態系統Ecopath模型的運行結果如表2所示。 將其中一些重要的參數與劉其根[12]構建的2004年模型結果以及于佳等[20]的2016年模型結果進行比較。生物量列表如下表3,由于千島湖生態系統種群結構的變動以及劃分功能組的不同,此處選取一些具有代表性的功能組作對比。 各功能組生物量變化可以經由表3看出。鰱、鳙屬于人工放養魚類,其生物量大小主要取決于放養和捕撈情況。其它魚類里,鳡魚的生物量經由1999和2000年的除鳡行動在2004年已經大幅減少,到2010年幾乎完全消失。鲌魚、飄魚、參條和鲴魚等魚在2004年的生物量相較于保水漁業實施前有短期的提升[12]。但經過六年后卻都大幅度減少,到2016年仍呈現繼續減少的趨勢。除此之外,底棲動物的生物量也大幅減少,浮游植物和有機碎屑的生物量卻成倍增長。 表2 千島湖2010年生態系統模型功能組參數和輸出結果 表3 3個年份的各功能組生物量對照 為了更直觀的表示出食物網關系,將不同功能組的營養流合并為數個營養級,稱為整合營養級[35]。2010年的能量流動過程可以形象的用下圖1的林氏錐分析法圖來表示。在忽略了生物量和生產量等都非常低的營養級之后,千島湖生態系統2004年、2010年和2016年的整合營養級為4個(表4)。其中低營養級的能流在整體系統中占比較大,越往頂級越小,成典型的金字塔型。2004年營養級Ⅰ、Ⅱ的總流量分別為4214 t km-2a-1和2285 t km-2a-1,占全部營養級能流量的64.5%和35%;2010年營養級Ⅰ、Ⅱ的總流量分別為7057 t km-2a-1和3737 t km-2a-1,占總能流量的65.0%和34.4%;2016年營養級Ⅰ、Ⅱ的總流量分別為9991 t km-2a-1和3683 t km-2a-1,占總能流量的72.8%和26.9%。在流向碎屑的總量上,2004年為3483 t km-2a-1,2010年為5921 t km-2a-1,2016年為8826 t km-2a-1,分別占總能流的53.3%、54.6%和64.3%。 圖1 千島湖2010年生態系統各營養級之間的能量流動/(t km-2 a-1)Fig.1 Energy flows between trophic levels of Qiandao Lake ecosystem in 2010 表4 千島湖3個年份生態系統整合營養級能流分布 從表5可見,2010年的生態系統在生產力和營養物總流通量上較2004年有較大程度的增加,2016年則又在2010年的基礎上提升明顯。2004年系統的總生產力為4440.0 t km-2a-1,到2010年升高為7341.1 t km-2a-1,增加了65 %,2016年則為10244 t km-2a-1,為原先的2.3倍;總凈初生產力為初級生產者產出的產量總和,2010年和2016年分別為7059 t km-2a-1和9991 t km-2a-1,增幅分別為67%和137%;在總消耗和總呼吸值上,2004年到2010年和2016年的數值逐漸變大;系統的總流通量通常可以表征生態系統的規模,2004年的值為16041.0 t km-2a-1,2010年和2016年則分別為20112.6 t km-2a-1和24698.2 t km-2a-1,呈現持續增大的趨勢。 表5 千島湖生態系統3個年份的總體特征 表5中還陳列出一些可以反映生態系統成熟度特征的參數。系統總初級生產量/總呼吸量(TPP/TR)在三年間的值分別為3.73、4.35和6.51,遠大于1且逐漸升高。連接指數是該生態系統食物網的實際連接數與可能連接數的比率;系統雜食指數被定義為所有消費者的平均雜食指數[34],這兩個指數可以在一定程度上反映出生態系統內部食物網聯系的復雜程度。千島湖生態系統3個年份的連接指數(CI)和雜食性指數(SOI)相差不大,在雜食性指數上略有增大的趨勢。 Finn氏循環指數和 Finn氏平均路徑長度也與系統成熟度、復原力和穩定性密切相關[34]。如表6所示,千島湖生態系統2004年、2010年和2016年的Finn氏循環指數(FCI)分別為26.27%、11.13%和5.27%,Finn氏平均路徑長度(FMPL)為3.803、2.853和2.472,兩項指標的數值都大幅減小。 表6 千島湖生態系統3個年份的網絡分析指數 從上述數據中可以總結出2010年千島湖生態系統在營養級、能流狀況等方面的特征。2010年的千島湖中除人工投放的鰱、鳙魚之外,大多數魚類和底棲動物的生物量都較低,浮游植物和有機碎屑的生物量卻較大。整合營養級的營養能流分布成典型的金字塔型,且高、低營養級的能量轉化效率相差很大,總體來說低于林德曼提出的生態金字塔1/10效率,以及Christensen等[36]在1993年總結多個生態系統模型提出的生態系統平衡傳輸效率(9.2%)。高強度捕撈干擾和能量傳遞阻塞的現象較為明顯。 生態系統的成熟度是相對于系統幼態化的一個概念,一個成熟度高的生態系統會更加穩定、更能抵抗外部沖擊。Ecopath模型從Odum提出的24個用以表征生態系統結構和功能特征的指標中提取了一些比較重要的參數進行量化表示[37]。如系統總初級生產量/總呼吸量在系統的早期發展階段會大于1,因為生產量會超過呼吸作用進行原始積累,越成熟的系統中,該比率應該越接近1[34]。千島湖生態系統2010年的TPP/TR為4.35,大于其在2004年的值,與1999年和2000年的值(分別為2.07和1.99)相比也更大[38],反映出其成熟度的降低。與國內外其它水庫相比,2010年千島湖的TPP/TR高于印度的Wyra水庫(其在實施漁業管理措施后降低至1.053)[39]和三峽水庫的1.899[40],低于巴西Itaipu水庫的6.3[41],處于不夠成熟的發展階段。除此之外,千島湖生態系統的CI和SOI值分別為0.219和0.100,相比于三峽水庫(0.371和0.205)[40],太湖(0.21和0.04)[28]和印度的Bakreswar水庫(0.18和0.11)[42]等湖泊,表現出其在食物網結構和飲食組成的多樣性方面仍缺乏復雜性。同時,2010年生態系統的FCI和FMPL分別為10.62%和2.843,與同類型其它水庫相比較低。FCI代表一個生態系統的循環量,與整個系統的穩定性、恢復力和成熟度相關。它的值較高說明一個系統更加穩定和成熟[37],較低則表示生態系統較為脆弱,對營養輸入的改變比較敏感[43]。模型計算出的浮游植物的高生物量在一定程度上證實了這一統計結果。由此可見,千島湖2010年的生態系統穩定性較差,比較容易受到外界因素的干擾或破壞。 本研究通過Ecopath模型的對比分析,探明了千島湖生態系統自實施了保水漁業后,在營養級狀況、總體特征上的發展演變趨勢。千島湖漁獲物的平均營養級呈現一直降低的趨勢,低營養級漁獲物數量增加,這種現象出現的原因可能是:自實施非經典生物操縱以來,頂級捕食者被大量清除,營養級較低的鰱、鳙魚則被大量補充;且隨著系統的發展,頂級消費者的群落組成和生物量都發生了明顯的變化,特別是一些營養級較高的肉食性魚類如鲌魚生物量的減少,可能也是使得平均捕撈營養級下降的原因。從整合營養級來看,從2004年至2016年千島湖系統低營養級占比在逐漸擴大,營養流中流向碎屑的部分也越來越大。營養級Ⅰ、Ⅱ的能量利用都不夠充分,近年來能量傳遞阻塞的現象更加明顯。且在持續性的人為干預下,系統內的魚群種類結構和生物量均發生較大變動。鳡魚被大量清除后,到2010年已經近乎完全消失。在保水漁業實施后的短期內,可能由于頂端捕食壓力的減少和餌料的增加等原因,2004年的鲌類、鲴類、大眼華鳊、參條等魚的生物量較2000年之前有一定上升[12]。但長時間的人工放養和捕撈使千島湖的漁業資源結構發生變化, 食物網趨于簡單,發展至10年之后,除了主要由人工放養控制的鰱、鳙魚之外,魚類資源量整體下降。然而,浮游植物和碎屑生物量卻有比較明顯的增加,這一方面可能是魚類生物量下降的原因,另一方面是由于周圍面源污染和水產養殖的剩余餌料在深水湖泊中的累積。 在能流特征上,系統發展后期和保水漁業實施第一年前后差別較大。劉其根等[38]在分析2000年的生態系統時發現,相比于1999年,系統的規模減小,但成熟度和穩定程度均有提升,藍藻水華等也得到了一定程度的克制。但不可忽視的是,頂級捕食者和較高營養級魚類生物量的變動勢必會使整個系統的能流格局發生改變。TPP/TR的值逐漸增大,說明系統處于不成熟的發展階段,且有成熟度下降的趨勢。Finn氏循環指數(FCI)和Finn氏平均路徑長度(FMPL)每過六年就幾乎削減一半,表明在長期的鰱、鳙魚放養累積下,生態系統的營養交互關系減弱,再循環率較低,復原力降低。系統退化到不成熟的擁有較高生產力的發育期。 綜合上述指標分析,保水漁業實施初期系統規模瞬減,隨后又自我調控,不斷發展積蓄能量。但可能由于系統營養結構和外源輸入的變化,初級生產者產出的能量未能得到有效的利用,很大一部分能量流向碎屑,魚類資源量整體下降,可能會造成湖體營養物質過剩,影響生態系統的健康穩定發展。在保水漁業等修復工作實施后的十幾年間,生態系統的營養級結構發生了一定程度的惡化。千島湖作為一個由水庫發展而來的淡水水體,捕食與被捕食的關系本身就較為簡單,生態系統更容易被外界的擾動影響,故而應更加注意控制人類活動和人為手段。 “保水漁業”采用非經典生物操縱的思路,圍網放養濾食性的鰱魚和鳙魚,直接攝食浮游植物,以期減少藍藻水華。但鰱、鳙魚主要抑制的是枝角類和大型浮游植物,間接促進橈足類和小型藻類種群增長,浮游植物的豐度增加。故而可能會出現藍藻水華得到抑制,但湖域富營養化程度加重的情況。低營養級再通過上行效應影響系統中的其它組分,系統整體的穩定性降低。同時,浮游生物食性的魚類還會加快氮、磷的循環速率,擴大初級生產力。上述分析也與文章第二部分中浮游植物生物量的逐漸上升的趨勢,以及初級生產力規模的逐漸擴大相對應。 諸多國內外學者也曾探索生物治理措施對水生態系統造成的影響,研究濾食性魚類對藍藻水華和富營養化進程的作用。在不同水域、不同實驗條件和不同研究方案下得出的結論不盡相同。王銀平等通過原位培養濾食性魚類鰱、鳙攝食微囊藻后的排泄物,利用葉綠素熒光技術研究鰱、鳙濾食對藻類光能及生長活性的影響,結果證明鰱、鳙單次濾食未對微囊藻造成生理上的致命損傷,反而可能會藻類超補償生長[44]。而美國學者Mueller等[45]在斑點叉尾鮰養殖池塘中的實驗,以及一些國內學者在東湖[46]、滆湖[47]的實驗則說明了鰱、鳙控藻的可行性。因此,需要在長期、規范的實驗下明確放養魚類和系統其它組分之間的聯系,準確把握不同實驗結果背后的生態學原理,才能更好地驗證非經典生物操縱理論的有效性。 鰱鳙魚放養的生態修復方式使千島湖的食物網結構發生了較大的變動。對淡水生態系統進行生態修復工作后,系統的結構化是逐漸推進的。人為的修復工作不止會影響直接作用的某幾個物種,還會通過系統內部的聯系傳遞到其它部分,從而影響整體。放養鰱鳙魚可能會使藍藻“水華”在短期內得到抑制,但經過一段時間的發展后,系統可能會呈現魚類資源量下降、能量轉換效率降低、成熟度和穩定度下降的狀態。因此,在進行某項修復工作之前,需要從生態學原理上系統地考慮問題,從避免過多的人為擾動出發,盡量還原生態系統本身的恢復力。2 結果與分析
2.1 生物量和營養級結構

2.2 系統能量流動特征

3 討論
3.1 生態系統總體特征
3.2 生態系統變化趨勢
3.3 鰱鳙魚放養影響
4 結論
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
工業設計(2022年8期)2022-09-09 07:43:20
今日農業(2021年11期)2021-08-13 08:53:34
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
家庭影院技術(2017年9期)2017-09-26 03:41:45
