鳳眼蓮入侵程度對金魚藻和黑藻生長及種間關系的影響
2022-09-13 08:40:34蔡竟芳高海燕沈一欒阿斯哈李紅麗
生態學報 2022年16期
張 羽,蔡竟芳,萬 云,高海燕,孫 凱,沈一欒,阿斯哈,李紅麗 ,*
1 北京林業大學生態與自然保護學院,北京 100083 2 南京大學生命科學院,南京 210073 3 鄂爾多斯生態環境職業學院,鄂爾多斯 017010
生物入侵已成為破壞濕地生態系統日益嚴重的生態問題,是繼生境破壞和破碎化之外對生物多樣性有嚴重影響的又一威脅因素[1],外來入侵種能夠在短時間內占據廣泛的空間快速繁殖,引起本地物種種群特性的變化,造成本地物種的消失甚至滅絕,破壞生態系統穩定性[2]。外來入侵種往往會導致生境破碎化,通過競爭或者直接占據本地種生態位從而排擠本地種,或者與本地種競爭資源,使本地物種的種類和數量減少,物種多樣性降低,一些種群的遺傳多樣性降低,甚至導致本土物種瀕危或滅絕[3—4]。鳳眼蓮(Eichhorniacrassipes)被稱為水域的“癌細胞”,擴展蔓延速度快,可以在很短時間內渡過適應期,依靠其繁殖優勢和擴展能力,迅速增加種群數量,很快在入侵成功的生態系統中占據重要生態位,對原生態系統造成破壞[5]。研究表明鳳眼蓮可以在水面擴散形成致密層,降低水下植物的可利用光資源,占據生態位和生境,使本地種失去競爭能力[6—7],進而導致沉水植物種間關系改變,甚至會引發沉水植物的死亡。沉水植物的死亡又使水體中溶解氧含量降低,導致其他魚類死亡,使生態鏈失去平衡[8]。
沉水植物是河流、湖泊等濕地生態系統中的重要初級生產者,為濕地動物提供豐富的食物,還可以為它們提供棲息環境和繁殖場所[9],對生態系統的健康和穩定有著重要作用[10—11]。沉水植物在水體中占據的生態位相同,生長期也大致相近,必然存在種間競爭關系[12]。植物間的相互作用直接影響植物個體的生長發育、形態特征,同時也影響植物群落的分布乃至整個生態系統的結構和功能[13—14]。并且,植物間相互作用的關系和強度也會隨著環境因子的變化而變化[15—16]。其中濕地植物入侵也是影響沉水植物生長和種間關系變化的環境因子之一。外來植物入侵過程通常被分為引入階段、建群階段和擴散階段,但在實際調查中很難界定這不同階段的界限和范疇。故此,研究中更多使用入侵種覆蓋范圍、植株密度等指標來實現入侵程度的衡量[17],入侵植物重要值涵蓋入侵植物的數量、高度、蓋度,眾多研究采用此指標來分析入侵程度對本地物種的影響[18—19]。而在外來植物入侵過程中對本地種的影響并非單純的抑制作用[20—21],李安定等人對空心蓮子草(Alternantheraphiloxeroides)的研究發現本地種多樣性指數和均勻度指數均表現為隨空心蓮子草重要值先增加后減少,并且在水生和濕生環境中蓮子草的臨界重要值也不同[22]。而關于鳳眼蓮入侵程度對沉水植物生長和種間關系的研究鮮見報道。
目前針對外來植物鳳眼蓮的入侵機制,以及入侵植物對本地單一沉水植物的影響有一定的報道,而關于鳳眼蓮入侵程度對沉水植物生長及種間關系影響的研究相對缺乏。本文通過控制實驗,設計入侵植物鳳眼蓮不同入侵程度(以鳳眼蓮不同株數形成水面覆蓋程度設定),定量分析研究鳳眼蓮入侵程度對沉水植物生長和種間關系的影響。試圖解決以下問題:(1)鳳眼蓮不同入侵程度對沉水植物生長指標的影響如何變化?(2)鳳眼蓮不同入侵程度對沉水植物種間關系有什么影響?(3)鳳眼蓮不同入侵程度對水質如何影響?(4)鳳眼蓮入侵、沉水植物生長及水體理化性質之間如何相互影響?
1 材料與方法
1.1 實驗材料
金魚藻(Ceratophyllumdemersum)是金魚藻科金魚藻屬的沉水性多年生水草,花期6—7月,果期8—9月。金魚藻多生長于小湖泊靜水處,于池塘、水溝等處常見,生命力較強,適溫性較廣,在水溫低至4℃時也能生長良好。為魚類的餌料,也可作為豬的飼料[23]。
黑藻(Hydrillaverticillata)是水鱉科黑藻屬的單子葉多年生沉水植物,花果期5—10月。廣布于池塘、湖泊和水溝中。喜陽光充足的環境,環境蔭蔽則植株生長受阻,在15—30℃的溫度范圍內生長良好,越冬不低于4℃。適宜淺水綠化、室內水體綠化,是良好的沉水觀賞植物[24]。
鳳眼蓮(Eichhorniacrassipes)是雨久花科鳳眼藍屬的漂浮性水生植物,被列入世界百大外來入侵種之一,廣布于中國長江、黃河流域及華南各省。喜歡溫暖濕潤、陽光充足的環境,適應性很強。適宜水溫18—23℃,超過35℃也可生長,氣溫低于10℃停止生長,具有一定耐寒性。喜歡生于淺水中,隨水漂流,繁殖迅速。生命力頑強,在干旱環境下種子也可保存15—20a[25]。
1.2 實驗設計
實驗所需要的沉水植物及相關實驗材料均取自北京奧林匹克森林公園,并協同水體,帶回實驗區域進行培養。
實驗采用雙因素設計,包括鳳眼蓮入侵程度3種模式和鄰株狀態3種定植模式。設置鳳眼蓮入侵程度3種模式,包括無入侵,入侵程度蓋度約25%(以鳳眼蓮入侵植物1株),入侵程度蓋度約75%(以鳳眼蓮入侵植物3株)。鄰株狀態3種定植模式,分別為沉水植物金魚藻無種間競爭模式、沉水植物黑藻無種間競爭模式、兩個物種混種存在種間競爭模式。實驗共9種處理,分別是:(1)無入侵金魚藻單種模式;(2)無入侵黑藻單種模式;(3)無入侵金魚藻和黑藻混種模式;(4) 鳳眼蓮蓋度約25%金魚藻單種模式;(5) 鳳眼蓮入侵蓋度約25%黑藻單種模式;(6) 鳳眼蓮入侵蓋度約25%金魚藻和黑藻混種模式;(7) 鳳眼蓮蓋度約75%金魚藻單種模式;(8) 鳳眼蓮入侵蓋度約75%黑藻單種模式;(9) 鳳眼蓮入侵蓋度約75%金魚藻和黑藻混種模式(圖1)。
2018年8月20日,實驗植株均取自北京奧林匹克森林公園。將生長健壯,長勢均一的金魚藻與黑藻分別剪取約8 cm長的帶芽莖段作為實驗材料,定植在的實驗容器(上徑29 cm,下徑27 cm,高38 cm)中,實驗基質底泥采自北京市野鴨湖濕地公園。基質深度約為10 cm,有機質含量為 (0.797±0.050) g/kg,總氮含量為 (0.105±0.030) g/kg,總磷含量為 (0.146±0.060) g/kg。植株種植完畢在桶內水體表面放置0株、1株或3株入侵植物鳳眼蓮,分別模擬鳳眼蓮入侵程度為0、25%和75%的沉水植物生長環境(圖1)。實驗設置5組重復。每2—3天澆水一次,水體為未經處理的普通自來水。
實驗于北京市海淀區北京林業大學三頃園苗圃內進行,實驗時間為2018年8月27日至10月13日,共48天。
實驗結束收獲前首先用取水管于每個處理取15 mL水樣用于水質檢測,然后小心取出植物進行生長指標測定。
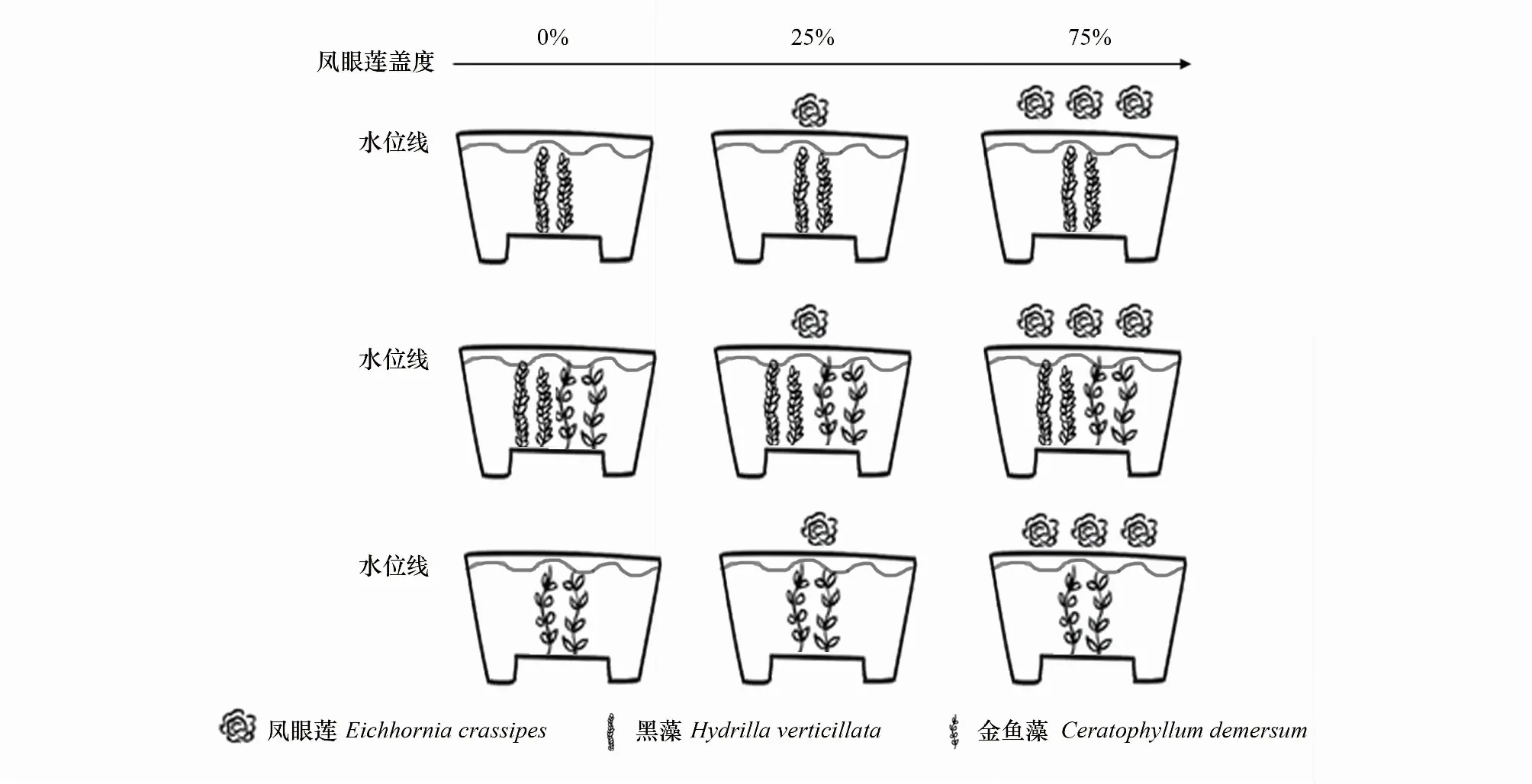
圖1 實驗設計圖Fig.1 Experimental design 實驗設計9個處理,每個處理設置5組重復,共45組樣品
1.3 數據分析
實驗結束后,分別收獲入侵植物鳳眼蓮,及各個處理容器中的沉水植物。沉水植物測量生長指標包括株高,分節數和分枝數。將收獲的沉水植物整株放入信封內,于烘箱中70゜C烘至恒重(超過48 h),分別稱量各部分生物量并計算總生物量。由于入侵植物鳳眼蓮主要作為處理環境所用,沒有進行細分各個指標。
對于各個實驗容器的水體,進行測定水體中的總氮、總磷和水體透明度。其中利用堿性過硫酸鉀消解紫外分光光度法測定水體中的總氮含量、利用鉬酸銨分光光度法測定水體中總磷含量,利用塞氏盤法測定水體中的透明度。
根據烘干稱重得到的單株生物量并采用對數響應比lnRR(ln response ratio)來計算沉水植物的種間競爭系數[26],采用公式如下:
lnRR=ln(B+/B0)
式中,B+為有競爭關系時平均單株生物量,B0為無競爭關系時平均單株生物量。
相對鄰體效應指數的數值范圍為-1—1,其中值小于0時,表示種間關系中鄰株對植物的生長產生抑制作用,值越大表示鄰體競爭效應越小;當RNE數值大于0時,則表示種間關系中鄰株對植物的生長產生促進作用,值越大表示鄰體促進效應越大。
采用雙因素方差分析(Two-Way ANOVA)分別檢驗鳳眼蓮入侵程度與種植模式及兩者交互作用對金魚藻和黑藻生物量、株高、分枝數和分節數的影響,以及水體中總氮、總磷和透明度的影響。在進行方差分析前,對數據進行正態性檢驗和方差齊性檢驗。用單因素方差分析不同處理下相對鄰體效應指數處理間是否存在顯著性差異。數據統計分析用SPSS 20.0統計軟件進行。使用Duncan方法進行顯著性檢驗(P<0.05)。使用結構方程模型(structural equation model, SEM)分析鳳眼蓮入侵強度以及水體理化性質對沉水植物生長的影響大小,利用SEM的標準回歸化系數,即指標間的路徑系數量化鳳眼蓮入侵或水體性質對沉水植物生長的影響程度。在構建初始模型時,假設沉水植物群落、入侵植物群落、水體理化性質任意兩個變量之間,及變量的指標之間都會產生相互作用,這樣的模型為飽和模型。通過刪除不顯著的變量關系(P>0.05的路徑),或者基于不同變量之間的殘差協方差來增加變量關系,對初始模型進行修改。利用χ2(當P>0.05模型擬合較好),擬合指數(CFI; 當CFI>0.95模型擬合較好)和近似的均方根誤差(RMSEA; 當RMSEA<0模型擬合較好)評估整體模型。在Sigma Plot 14.0中進行作圖。
2 結果與分析
2.1 鳳眼蓮入侵程度和定植模式對沉水植物金魚藻生長繁殖的影響
鳳眼蓮入侵程度對沉水植物金魚藻生物量、分枝數具有顯著影響,定植模式對金魚藻生物量也具有顯著影響,同時兩者的交互作用對金魚藻所有的生長繁殖指標如生物量、株高、分枝數和分節數均具有顯著影響(表1)。
鳳眼蓮不存在時,金魚藻生物量、株高、分枝數和分節數均為單種模式高于混合模式(圖2);隨鳳眼蓮蓋度增加,金魚藻生物量顯著減少,株高有所降低,但變化不顯著,金魚藻分枝數則顯著增加。同時,隨鳳眼蓮蓋度增加,金魚藻各生長指標如株高、分枝數和分節數先是混合模式高于單種模式,之后趨勢相反(圖2)。
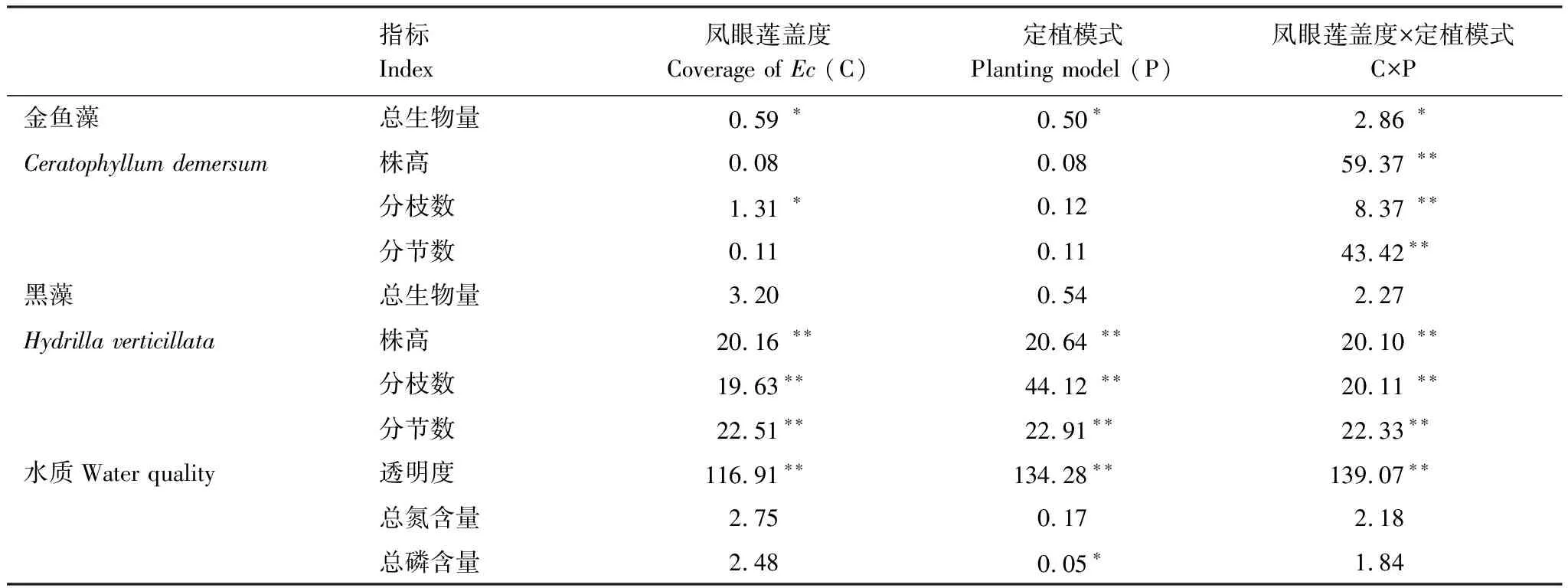
表1 鳳眼蓮入侵程度和定植模式對沉水植物生長指標及水質影響的方差分析表
2.2 鳳眼蓮入侵程度和定植模式對沉水植物黑藻生長繁殖的影響
鳳眼蓮入侵程度、定植模式和兩者的交互作用對沉水植物黑藻株高、分枝數和分節數均顯著影響,而對生物量無顯著影響(表1)。
鳳眼蓮不存在時,黑藻生物量、株高、分枝數和分節數均為單種模式高于混合模式(圖3);隨著鳳眼蓮入侵,但是其蓋度較小時,黑藻生物量有所降低,其生物量和株高單種模式小于混合模式,而分節數卻趨勢相反;隨著鳳眼蓮蓋度增加,黑藻的各項生長繁殖指標均降低,且均為單種模式高于混合模式(圖3)。
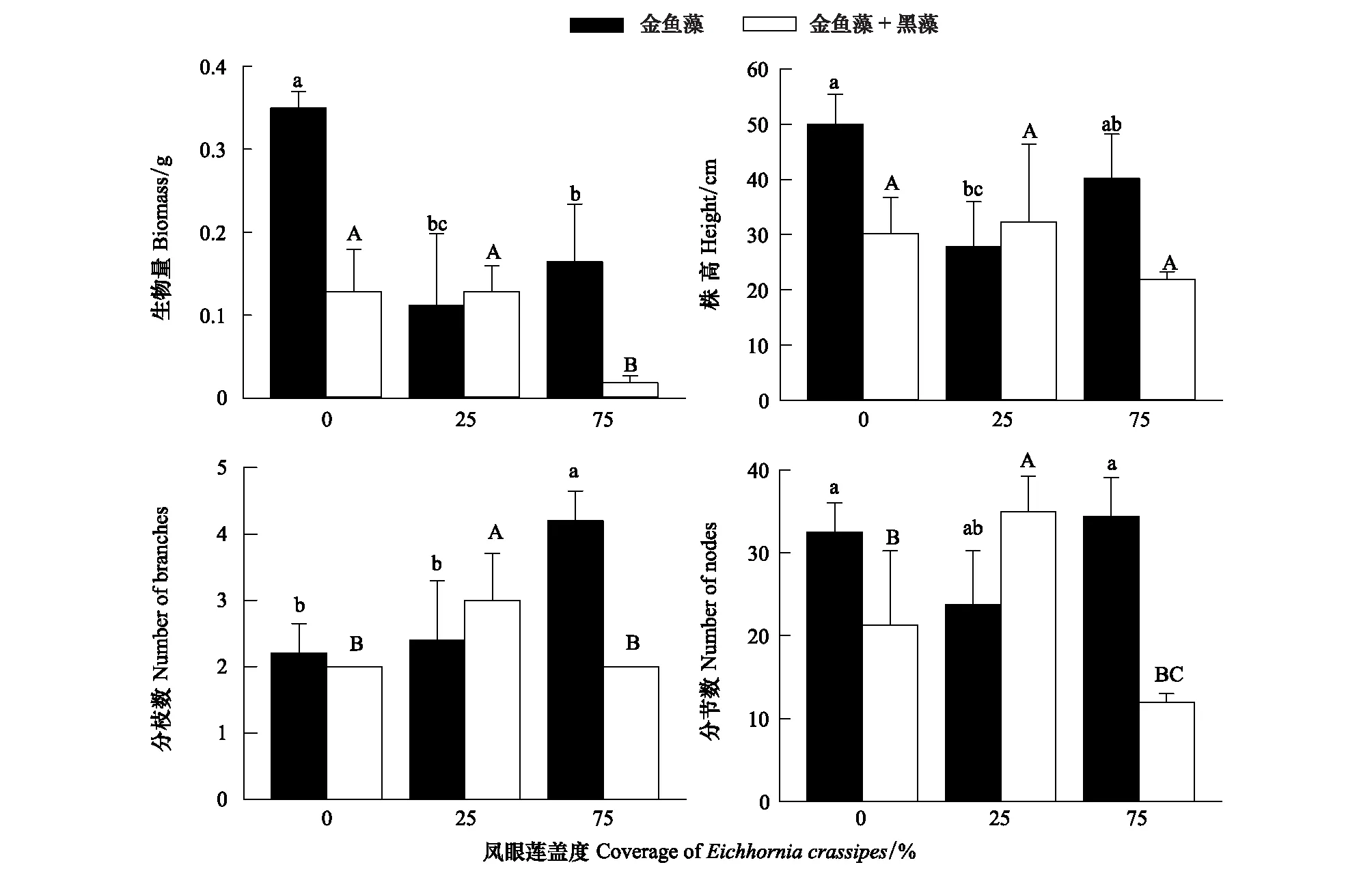
圖2 鳳眼蓮入侵程度和定植方式對金魚藻生長繁殖指標的影響 (平均值+標準誤)Fig.2 Effects of different invasive degree of Eichhornia crassipes on the growth index of Ceratophyllum demersum (Mean+SE) 不同小寫或大寫字母表示不同入侵程度同一模式下差異顯著(P<0.05)
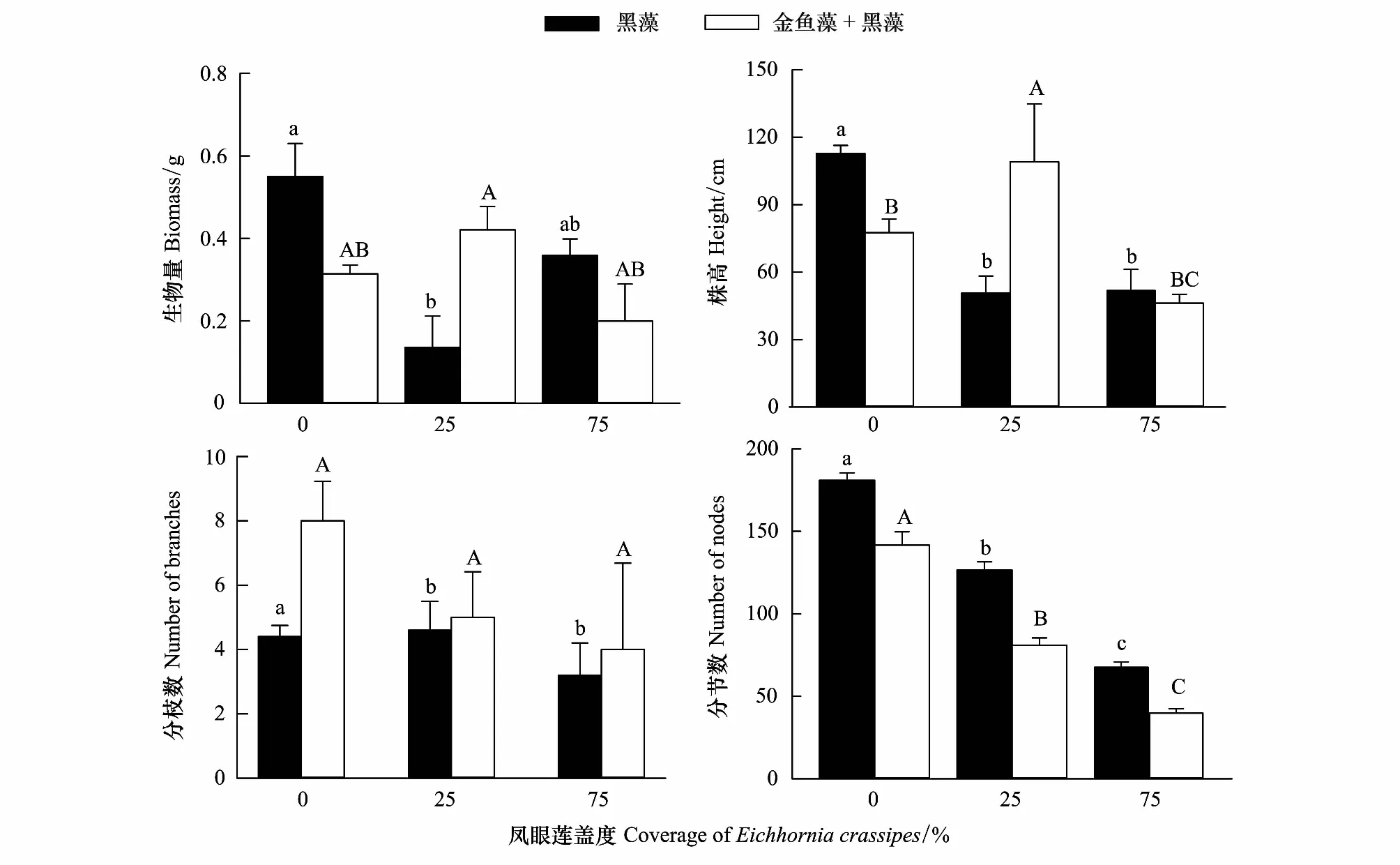
圖3 鳳眼蓮入侵程度和定植方式對黑藻生長繁殖的影響 (平均值+標準誤)Fig.3 Effects of different coverage of Eichhornia crassipes on the growth index of Hydrilla verticillate (Mean+SE) 不同小寫或大寫字母表示不同入侵程度同一模式下差異顯著(P<0.05)
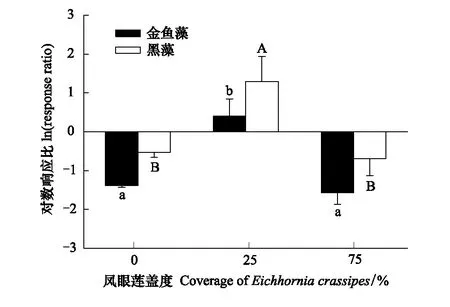
圖4 鳳眼蓮入侵程度對沉水植物種間關系的影響 (平均值+標準誤)Fig.4 Effects of different coverage of Eichhornia crassipes on the interspecies relationship of submerged plants (Mean+SE) 不同小寫或大寫字母表示不同入侵程度同一模式下差異顯著(P<0.05)
2.3 鳳眼蓮入侵程度對金魚藻和黑藻種間關系的影響
當鳳眼蓮不存在時,兩種沉水植物金魚藻和黑藻種間關系指數均小于0,說明此時兩種沉水植物單種生物量高于混種,種間關系表現為競爭作用;隨著鳳眼蓮入侵,兩種沉水植物種間關系指數均大于0,表明其沉水植物單種模式生物量小于混種模式,種間關系表現為促進作用,說明隨著入侵脅迫變化,物種間的競爭關系削弱,而轉變為促進作用共同抵御其脅迫生境。但隨著其鳳眼蓮入侵程度增加,兩者的種間關系指數又變回小于0,表明兩者又呈現了競爭作用(圖4) 。
2.4 鳳眼蓮入侵程度和定植模式對水質的影響
鳳眼蓮入侵程度、定植方式和兩者的交互作用均對水體的透明度顯著影響,同時定植模式對水體的總磷含量具有顯著影響,其余均無顯著影響(表1)。
隨鳳眼蓮蓋度增加,水體中總氮TN含量無顯著變化,總磷TP含量先略微上升后下降,但受鳳眼蓮入侵程度影響不顯著。當鳳眼蓮不存在時,水體透明度較低;隨鳳眼蓮蓋度增加,水體透明度先增加后略微降低,但仍高于沒有鳳眼蓮入侵的對照組,說明鳳眼蓮對水體透明度有輕度凈化作用(圖5)。
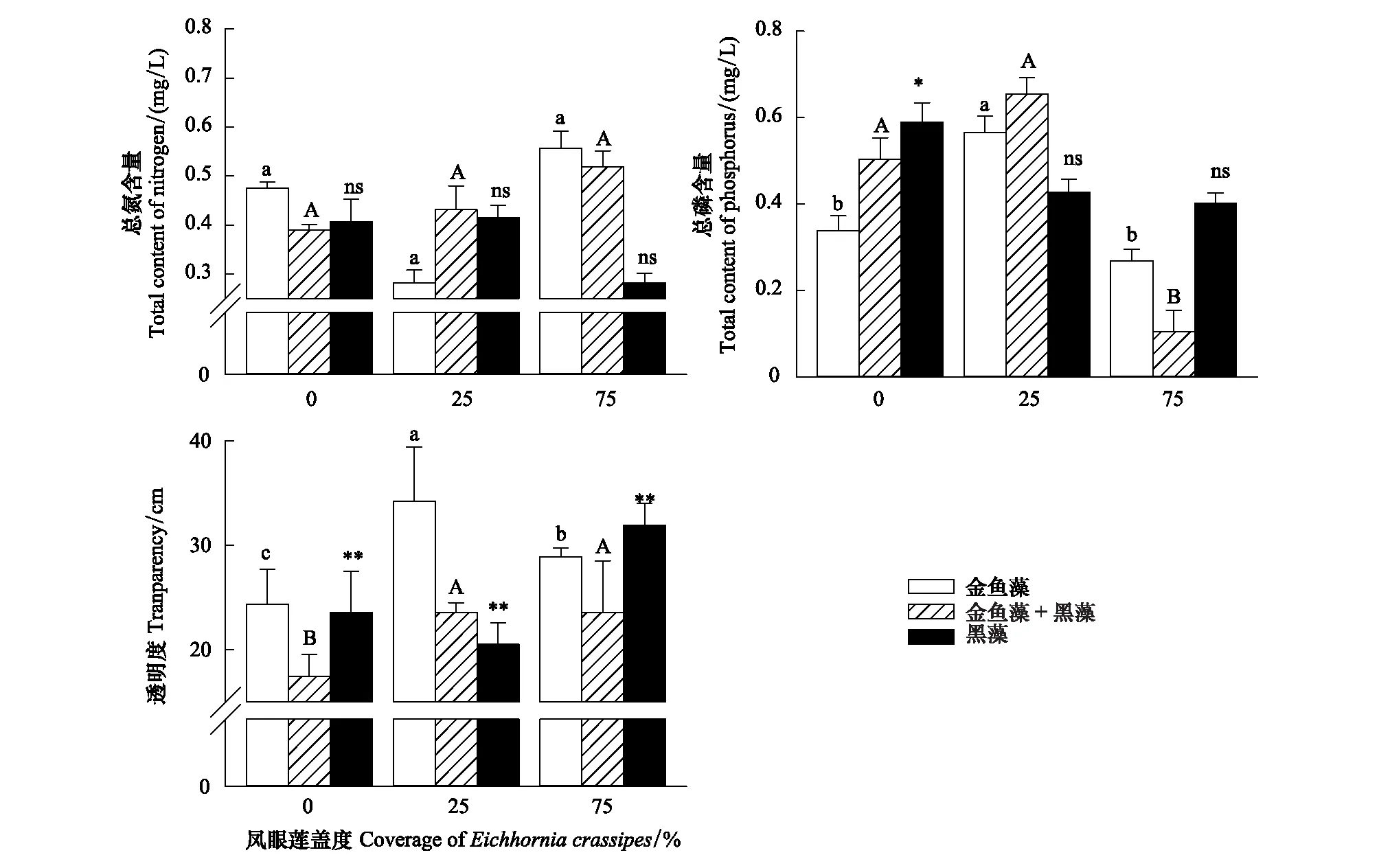
圖5 鳳眼蓮入侵程度和定植方式對水質的影響 (平均值+標準誤)Fig.5 Effects of different coverage of Eichhornia crassipes on water quality (Mean+SE) 不同小寫或大寫字母表示不同入侵程度下同一定植模式差異性顯著(P<0.05);ns代表無顯著性差異,*,**分別表示顯著性差異P<0.05、P<0.01
2.5 鳳眼蓮入侵和水環境因子對沉水植物生長影響的結構方程模型分析
在相關性分析基礎上,選取影響沉水植物群落特征的因素,通過建立模型、擬合模型、評價和修正模型后得到了沉水植物群落與入侵植物群落、水體理化性質相互關系的結構方程模型(圖6)(χ2=4.512,CFI=0.98,RMSEA=0.002)。沉水植物群落特征與株高、生物量、分枝數、分節數呈顯著正相關,其中與生物量相關系數最高,為0.439;其次為株高,相關系數為0.318。入侵植物群落特征與鳳眼蓮生物量呈顯著正相關,相關系數為0.708。水體理化性質與透明度、TN、TP呈顯著正相關,其中與TN相關系數最高,為0.625。植物入侵和TN等水體理化性質增加對沉水植物群落特征均有顯著負向影響(P<0.05),路徑系數分別為-0.487和-0.657。同時,入侵植物群落與水體理化性質也具有顯著的相關關系,相關系數為0.246,說明入侵植物群落與水體理化性質相互影響,并且水體理化性質較入侵植物對沉水植物影響更大。
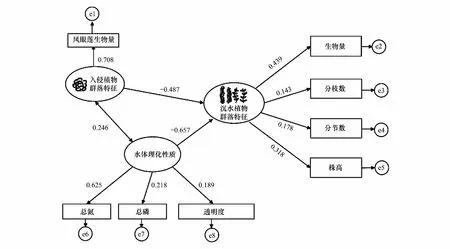
圖6 入侵植物群落、沉水植物群落與水體理化性質相互關系的結構方程模型Fig.6 SEM of the relationship between invasive plant community, submerged plant community and the physical and chemical properties of water 橢圓代表潛在變量,長方形代表可測變量,圓代表殘差,雙箭頭表示相關系數,單箭頭表示路徑系數
3 討論
3.1 鳳眼蓮入侵程度和定植模式對沉水植物生長繁殖指標的影響
植物入侵程度的影響主要與植物本身入侵性,及本地物種群落的脆弱性,群落的空間尺度大小等密切相關。研究表明黃頂菊(Flaveriabidentis)的入侵顯著降低了植物群落的物種多樣性指數和豐富度指數[27],其它研究表明隨著反枝莧(Amaranthusretroflexus)入侵程度的增加,本地群落植物的物種數、株高及生物量均顯著降低[28];而在加拿大一枝黃花(Solidagocanadensis)中度和重度入侵區域群落生物多樣性顯著下降,而輕度入侵區域群落多樣性則無顯著變化[20]。本研究結果與以往研究類似。
入侵植物鳳眼蓮具有較大的生態位寬度及較強的繁殖能力,對環境的適應能力極強,大量繁殖后可占據極大的空間。較強的資源利用能力使得鳳眼蓮可以成功入侵本地沉水植物群落并進一步擴散造成危害。夏秋季是鳳眼蓮和沉水植物的生長旺盛期,在此時段外來物種入侵對本地物種多樣性的負作用也相對顯著[29]。本實驗中鳳眼蓮的入侵使沉水植物的總生物量及株高明顯降低,同時對不同種類的沉水植物影響效果也不同。鳳眼蓮入侵使金魚藻生物量降低了約2.4倍,黑藻的生物量則降低了約1.5倍,對金魚藻生長的抑制作用更加明顯。說明黑藻對鳳眼蓮入侵的耐受能力更強。隨著鳳眼蓮入侵程度增加,金魚藻的分枝數也隨之顯著增加,分節數則沒有顯著變化,說明當入侵威脅加大時,金魚藻更傾向于橫向擴展,以尋求光照適應不利環境(圖2)。而黑藻的分枝隨鳳眼蓮蓋度的增加略微下降,而分節數則呈現顯著下降,說明雖然黑藻對鳳眼蓮有一定的耐受性,但隨著鳳眼蓮入侵程度增強,鳳眼蓮入侵將削弱了黑藻的生長繁殖。
比較鳳眼蓮相同入侵程度下兩種沉水植物的生長指標,可以明顯看出黑藻的生物量及株高均顯著高于金魚藻,黑藻在群落生長中占據競爭優勢。黑藻和金魚藻均屬于冠層型沉水植物,在水體中處于同一生態高度,不存在相互遮陰的可能,出現此現象的原因可能是由于黑藻強大的繁殖能力[30]。黑藻擁有斷裂生殖、塊莖、具鱗根出條以及種子繁殖四種繁殖方式,其中斷裂生殖由于黑藻質地清脆易斷,會產生大量斷枝,可使黑藻大量繁殖。并且,黑藻無性繁殖體的生根能力也強于金魚藻[31]。同時,黑藻在地下部分也有一定優勢。黑藻的不定根能夠迅速扎根在土壤底質中吸收營養,從而影響金魚藻生長,因此黑藻在競爭中占據優勢。
3.2 鳳眼蓮入侵程度對沉水植物種間關系的影響
在自然界中,物種不是孤立存在的,彼此間的相互作用普遍存在。種間關系是影響植物形態、生長、存活以及物種分布的重要因子,且在具有相似生態幅的物種間往往更容易發生激烈的競爭現象,植株的特征、資源水平和環境條件都會影響其競爭能力。在水生環境中,有效光合輻射和氮磷等營養物質也是影響植物競爭的重要環境因子[32]。而對于鳳眼蓮入侵會導致其光資源及營養物質等環境條件的變化,進而影響沉水植物的群落生產力,也可能導致其沉水植物的種間關系發生變化。研究表明當鳳眼蓮入侵顯著降低了沉水植物群落的生產力,主要由于其降低了優勢種黑藻的生物量[10],進而可能會導致沉水植物種間關系的改變。
無鳳眼蓮入侵時,金魚藻與黑藻之間的種間關系均小于0,表現為競爭關系,且金魚藻受到黑藻的競爭作用更強;隨鳳眼蓮入侵蓋度增加,兩者的種間關系由先促進作用后又變回競爭作用,表明在受到輕微脅迫時,兩者共同協助抵御其不良環境,但是隨著脅迫程度加重,兩者又呈現競爭關系,黑藻仍呈現較強的競爭作用,符合“脅迫梯度假說 (stress gradient hypothesis, SGH)”[33]。其他研究也表明當處于非脅迫條件時,辣蓼與苔草的地上部分對光的競爭顯著;但在脅迫水位條件下,植物間的競爭作用則非常微弱[34]。可能是因為相比于種間或種內的競爭作用,外界的環境脅迫更能影響植物的生長狀況[35],進而引起種間關系變化。
3.3 鳳眼蓮入侵對水質的影響
水體環境對濕地的結構和功能十分重要,其中營養物濃度是重要的環境控制因子,為植物的生長提供能量[36]。水體中營養鹽的有效性決定了植物的初級生產量,而過量的營養鹽會使水體中藻類以及其它水生生物異常繁殖,引起水體透明度和溶解氧的變化導致水質惡化和生態系統破壞。水體透明度下降時,光的通透量減少,喜光植物生長受到限制,沉水植物受到的影響則更大[37]。
本實驗中隨鳳眼蓮蓋度增加,水體中營養濃度變化不顯著。可能是鳳眼蓮對水體氮磷的去除主要依靠根系及根系微生物的吸附作用[38]、植株的吸收同化作用、微生物轉化作用及沉淀作用[39],而本實驗時間處于鳳眼蓮花期(9—11月),生長速率減緩,根系吸附作用不顯著[40]。他人研究發現, 鳳眼蓮在富營養化的水體中能夠使水體的透射光明顯下降[41]。而本實驗中鳳眼蓮入侵反而使水體透明度增加,可能是由于鳳眼蓮并未大量擴散繁殖, 沒有在水面上形成草墊遮擋陽光, 凈化作用占據主導,使水體更加清澈。植物對水體中氮磷的凈化作用是以植物為中心的植物與微生物綜合作用的結果,是一個持續進行的過程[42—43],本實驗持續時間相對于植物的生長期偏短,只能反映一段時間內鳳眼蓮對水體中的氮磷去除的作用,具有一定局限性。
3.4 鳳眼蓮入侵程度和水環境因子對沉水植物生長影響的結構方程模型分析
鳳眼蓮的入侵程度這一生物因素和總氮、總磷等水體環境非生物因素對沉水植物生長具有不可忽視的影響。多項研究表明,鳳眼蓮在適宜的生境中迅速繁殖,在水面上方形成細密的植物墊,大大抑制了水下光照,從而限制沉水植物生長[44]。水體透明度是反映水下光照的重要指標,直接影響沉水植物可接受到的光照強度,進而影響光合作用[45]。營養鹽濃度也會影響植物的生長,低濃度氮磷能促進沉水植物生長;高濃度氮磷則會導致藻類數量激增,水體透明度降低,不利于正常沉水植物生長。鳳眼蓮入侵程度和水體的理化性質對沉水植物生長均有顯著影響(P<0.05),這與多數前人研究一致[46],而水體理化性質對沉水植物生長的影響略大于入侵植物,推測是因為水體理化性質不僅可以直接影響沉水植物的生長繁殖,還會通過影響入侵植物的生長間接影響沉水植物[47]。將鳳眼蓮入侵強度對沉水植物生長及種間關系影響制成一個概念框架圖(圖7)。根據概念框架圖,無鳳眼蓮入侵時,沉水植物間表現為競爭關系;鳳眼蓮輕度入侵顯著降低了兩種沉水植物的生長繁殖,沉水植物種間關系表現促進。當鳳眼蓮重度入侵時,兩種沉水植物“各自逃逸”,重新相互競爭。
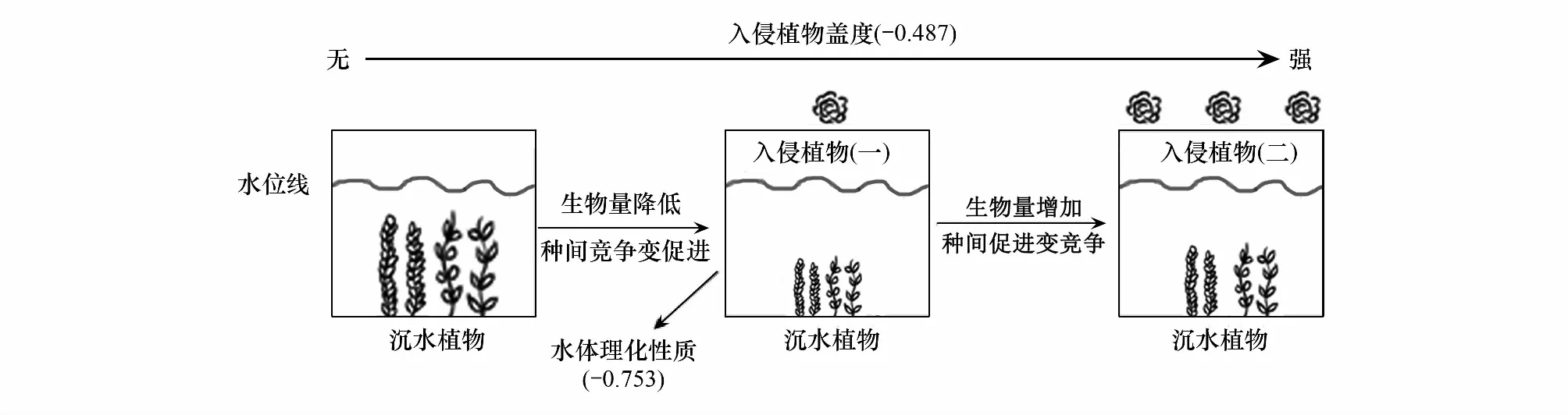
圖7 沉水植物生物量及種間關系決定因子示意圖Fig.7 Schematic diagram of determining factors of submerged plant biomass and interspecific relationships -0.487和-0.753分別為入侵植物蓋度和水體理化性質對沉水植物影響的結構方程模型系數
4 結論
綜上所述,鳳眼蓮的入侵使沉水植物的總生物量及株高均明顯降低,金魚藻生物量降低了約2.4倍,黑藻的生物量則降低了約1.5倍,對金魚藻生長的抑制作用更加明顯;當鳳眼蓮不存在時,金魚藻與黑藻之間的種間競爭關系較強;隨入侵蓋度增加,種間關系則先變為促進作用后又變回競爭作用。黑藻在沉水植物共存環境中占據了明顯的優勢地位;隨鳳眼蓮蓋度入侵程度增加,水體中TN、TP濃度均無顯著變化,水體透明度則略微降低,說明鳳眼蓮對水體透明度有輕度凈化作用。植物入侵及TN等水體理化性質增加對沉水植物生長均有顯著負向影響,且水體理化性質對沉水植物生長的影響略大于入侵植物。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
