虎奶菇菌絲體抗菌肽提取工藝優化及活性研究
2022-09-15 03:33:22張書會孫雪言馬愛民
食品與機械 2022年8期
張書會 羅 璐 孫雪言 馬愛民
(華中農業大學食品科學技術學院,湖北 武漢 430070)
抗菌肽(Antimicrobial peptides, AMPs)是含有100個以內氨基酸殘基的多肽抗菌物質,具有廣譜抑菌效果,對細菌、真菌、病毒以及寄生蟲等均有抑制作用[1],被廣泛應用于生物醫藥、食品及動物飼料加工領域。抗菌肽能夠通過破壞微生物的細胞膜或靶向細胞內成分產生抑菌作用[2]。截至2022年5月,已有3 000多種抗菌肽被收錄至抗菌肽數據庫(https://aps.unmc.edu)中。然而這些抗菌肽主要來源于動物、植物和細菌,抗菌肽數據庫中真菌來源的抗菌肽<1%。
虎奶菇(Pleurotustuber-regium)是一種既能形成菌核又能形成子實體的高溫食用菌,廣泛分布于熱帶和亞熱帶地區[3]。然而虎奶菇菌核和子實體的生長周期長,生物轉化率低,因此常用液體深層發酵培養的菌絲體作為試驗材料[4]。近年來,有關虎奶菇生物活性的研究主要集中于抗氧化活性[5]、抗腫瘤活性[6]、免疫調節活性[7]、抗炎活性[8]、抑菌活性[9]以及降血糖和降血脂活性[10]等方面。虎奶菇菌絲體中含有豐富的蛋白質和氨基酸[11],是天然抗菌肽的優質來源。但目前尚未見從虎奶菇菌絲體中提取抗菌肽的相關報道。
常見的抗菌肽制備方法主要包括酶解法、基因工程法、化學合成法以及微生物發酵法[12]。由于專一性強、反應條件溫和以及過程易于控制等優點,酶解法成為目前制備抗菌肽最常用的方法[13]。因此,研究擬以虎奶菇菌絲體為原料,通過酶解法提取抗菌肽。在單因素試驗的基礎上結合響應面試驗優化提取工藝,并對其抑菌活性進行研究,以期提高虎奶菇菌絲體抗菌肽的提取率和抑菌率,從而為虎奶菇菌絲體抗菌肽的進一步開發與利用提供依據。
1 材料與方法
1.1 供試菌株與主要試劑
虎奶菇菌株、金黃色葡萄球菌(ATCC25923)菌株、大腸桿菌(ATCC25922)菌株:華中農業大學食品科學技術學院食品微生物實驗室;
木瓜蛋白酶、中性蛋白酶、堿性蛋白酶、風味蛋白酶、復合蛋白酶:上海源葉生物科技有限公司;
胰蛋白酶、牛血清白蛋白:德國Biofroxx公司;
福林酚:北京索萊寶科技有限公司;
乙醇、氫氧化鈉、鹽酸、四水合酒石酸鉀鈉:分析純,國藥集團化學試劑有限公司。
1.2 主要儀器與設備
真空冷凍干燥機:Beta2-8LD型,德國Christ公司;
臺式電熱干燥箱:202-00AB型,天津市泰斯特儀器有限公司;
數顯恒溫水浴鍋:HH-4型,上海力辰邦西儀器科技有限公司;
臺式pH計:ST 2100型,美國奧豪斯儀器有限公司;
超速冷凍離心機:Avanti J-E型,美國Beckman公司;
超凈工作臺:VD-1320型,北京東聯哈爾儀器制造有限公司;
恒溫培養搖床:KYC-1102C型,金壇市盛藍儀器制造有限公司;
全自動酶標儀:Multiskan Skyhigh型,美國Thermo公司。
1.3 培養基
(1) PDA培養基:馬鈴薯200 g,葡萄糖20 g,瓊脂粉15 g,加蒸餾水至1 L,121 ℃高壓滅菌20 min。
(2) 發酵培養基:葡萄糖20 g,酵母提取物5 g,磷酸二氫鉀1 g,無水硫酸鎂0.5 g,加蒸餾水至1 L,121 ℃高壓滅菌20 min。
(3) LB培養基:胰蛋白胨10 g,NaCl 10 g、酵母提取物5 g,加蒸餾水至1 L,121 ℃高壓滅菌20 min。
1.4 試驗方法
1.4.1 液體深層發酵培養虎奶菇菌絲體 用孔徑為0.5 cm的打孔器將PDA培養基上生長的虎奶菇菌絲體打孔后,用滅菌的鑷子取8個菌絲塊接種至100 mL液體發酵培養基中,32 ℃、120 r/min培養8 d。培養結束后,將其作為液體菌種,用滅菌的玻璃勻漿器磨勻,以5%的接種量接種至新的發酵培養基中。相同條件下培養8 d,過濾洗滌,收集菌絲體,并冷凍干燥,獲得干燥的虎奶菇菌絲體。
1.4.2 樣品脫脂 將凍干后的虎奶菇菌絲體研磨成粉末,于索氏提取器中用無水乙醇浸提6 h脫脂。脫脂后的菌絲于50 ℃烘干,粉碎過篩,即得到脫脂虎奶菇菌絲體粉末。
1.4.3 酶解法制備虎奶菇菌絲體抗菌肽 參照文獻[14-15]并修改。稱取1 g脫脂菌粉,以料液比(m菌粉∶V水)為1∶20 (g/mL)溶解后,添加蛋白酶于最適pH及溫度下酶解3 h,加酶量為8 000 U/g。每隔1 h使用1 mol/L NaOH和1 mol/L HCl溶液維持體系中的pH恒定。反應結束,將虎奶菇菌絲體酶解液于沸水中水浴10 min,11 000 r/min離心30 min,收集上清液。
1.4.4 虎奶菇菌絲體抗菌肽的初步分離 根據文獻[15-16]并修改。向酶解上清液中加入4倍體積的無水乙醇,使乙醇體積分數為80%,以除去酶解上清液中的多糖。混合液于4 ℃靜置24 h,8 000 r/min離心15 min。將上清液旋轉蒸發去除乙醇,加入一定體積的蒸餾水,-20 ℃放置過夜,真空冷凍干燥,獲得虎奶菇菌絲體多肽粉末。
1.4.5 最適蛋白酶的篩選 分別用木瓜蛋白酶、中性蛋白酶、堿性蛋白酶、風味蛋白酶、復合蛋白酶和胰蛋白酶于其最適pH及溫度下酶解制備虎奶菇菌絲體多肽(表1)。將各蛋白酶酶解制備的虎奶菇菌絲體多肽粉末于干燥皿中貯藏用于后續多肽得率及抑菌率的測定。

表1 不同蛋白酶的理論最適酶活條件Table 1 Theoretical optimal conditions for enzyme activity of different proteases
1.4.6 單因素試驗 在蛋白酶篩選的基礎上,根據其最適蛋白酶的理論最適酶活范圍,分別考察酶解溫度(30,35,40,45,50 ℃)、酶添加量(6 000,8 000,10 000,12 000,14 000 U/g)、pH(9.5,10.0,10.5,11.0,11.5)、酶解時間(1,2,3,4,5 h)和料液比(V水∶m菌粉)[1∶10,1∶15,1∶20,1∶25,1∶30 (g/mL)]對抗菌肽抑菌活性及得率的影響。
1.4.7 響應面試驗 在單因素試驗的基礎上,確定響應面試驗的三因素和三水平。并根據Design-Expert 10.0.7軟件中的Box-Behnken中心組合設計原理設計響應面試驗,優化虎奶菇菌絲體抗菌肽的最佳酶解條件。
1.4.8 多肽得率測定 參照文獻[15]并修改。以牛血清白蛋白標準品濃度為橫坐標,吸光度值OD500 nm為縱坐標繪制標準曲線,得線性回歸方程y=0.769x+0.053 2(R2=0.998 2)。將多肽粉末加蒸餾水復溶,經50倍稀釋后取100 μL于1.5 mL離心管中,加入500 μL堿性銅試液,25 ℃反應10 min,隨后加入50 μL福林酚試劑混勻后繼續反應30 min,用酶標儀測定500 nm 處吸光值。空白組用超純水代替多肽溶液,按式(1)計算多肽得率。
(1)
式中:
R1——虎奶菇菌絲體多肽得率,%;
C——稀釋后多肽溶液的質量濃度,mg/mL;
n——多肽的稀釋倍數;
V——虎奶菇菌絲體酶解后獲得上清液的體積,mL;
m——脫脂菌粉樣品的質量,mg。
1.4.9 總糖含量測定 采用苯酚—硫酸法[17]。以葡萄糖含量為橫坐標,OD490 nm為縱坐標,繪制總糖含量測定的標準曲線為y=7.007 4x+0.067 8(R2=0.999 7)。并按式(2)計算總糖含量。
(2)
式中:
R2——總糖含量,%;
C——稀釋后酶解液中糖質量濃度,mg/mL;
n——酶解液的稀釋倍數;
V——酶解液體積,mL;
m——脫脂菌粉樣品的質量,mg。
1.4.10 抑菌活性測定 參照文獻[18]的方法,按式(3)計算抑菌率。
(3)
式中:
Y——抗菌肽抑菌率,%;
A0——培養12 h后對照組混合液在OD600 nm處的吸光度;
A1——培養12 h后樣品組混合液在OD600 nm處的吸光度。
1.4.11 最小抑菌濃度(MIC)測定 采用微量稀釋法[19]。
1.4.12 數據處理 每組試驗重復3次,采用Origin 2021軟件繪制圖形,Design Expert 10.0.7軟件對響應面試驗結果進行分析。
2 結果與分析
2.1 虎奶菇菌絲體抗菌肽的初步分離
由圖1可知,醇沉法脫糖后酶解液中總糖含量僅為2.95%,遠低于多肽含量,從而保證了多肽作為酶解液中抑菌的主要活性成分。此外,脫糖處理對酶解液中多肽含量的影響較小,脫糖處理前后酶解液中多肽含量分別為21.55%和20.43%,損失量較低。
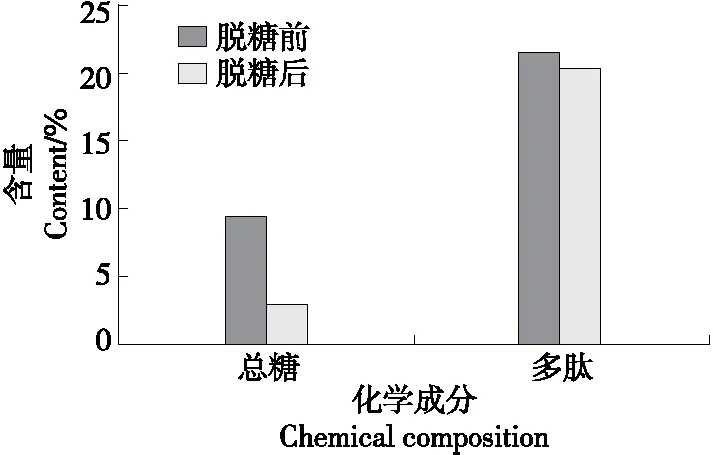
圖1 脫糖前后酶解液中多肽及總糖含量變化Figure 1 Changes of polypeptide and total sugar contents in enzymatic hydrolysate before and after desugarization
2.2 最適蛋白酶的篩選
由圖2可知,相比于其他蛋白酶,風味蛋白酶酶解制備的多肽得率最高,為18.59%。推測是因為風味蛋白酶是經復配而成的一種復合型蛋白酶,含有內切蛋白酶和外切肽酶,具有廣泛的作用位點,水解更徹底,因此多肽得率較高[20]。其次對虎奶菇菌絲體多肽得率影響較大的是堿性蛋白酶(17.05%),是因為堿性蛋白酶主要作用于C-端的某些疏水性氨基酸殘基,導致蛋白質結構疏松,能夠獲得較多的多肽[21]。林冰潔等[22]研究發現,相比于中性、復合和風味蛋白酶,堿性蛋白酶酶解制備的抗菌肽得率最高為88.88%,與試驗結果相似。
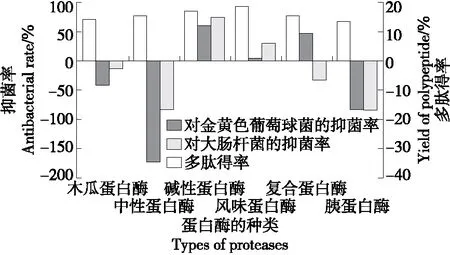
圖2 不同蛋白酶對虎奶菇菌絲體多肽得率及 抑菌率的影響Figure 2 Effects of different proteases on the yield and antibacterial rate of PTRM-AMPs
堿性蛋白酶酶解產生的多肽對金黃色葡萄球菌和大腸桿菌均具有抑制作用,且抑菌活性最強,分別為61.10% 和75.30%,與馬尚玄等[23]的結論類似。利用木瓜蛋白酶、中性蛋白酶和胰蛋白酶制備的多肽對金黃色葡萄球菌和大腸桿菌的生長存在不同程度的促進作用,可能是這些蛋白酶制備的多肽粉末中存在的含碳物質和含氮物質能夠促進金黃色葡萄球菌和大腸桿菌的進一步生長發育[18-19]。宋偉光[24]研究發現,堿性蛋白酶酶解產物中分子量<1 kD的多肽比例最高,為86.5%。而木瓜蛋白酶和中性蛋白酶中分子量<1 kD的多肽比例相對較低,分別為73.1%和72.4%。林賀等[25]發現<1 kD的多肽組分對大腸桿菌和金黃色葡萄球菌的抑菌作用最顯著。
綜上,以多肽得率及多肽對金黃色葡萄球菌和大腸桿菌的抑菌率為評價指標,與其他5種蛋白酶相比,堿性蛋白酶酶解虎奶菇菌絲體得到的多肽不僅得率較高,其抑菌活性也最強。因此,選取堿性蛋白酶用于制備虎奶菇菌絲體抗菌肽。
2.3 單因素試驗
2.3.1 酶解溫度 由圖3可知,當酶解溫度為30~50 ℃時,抗菌肽得率和抑菌率呈先增加后降低的趨勢,并在40 ℃時達最高,是因為過低的溫度不利于蛋白酶與底物的結合,酶促反應延緩,因此得到的抗菌肽片段也相對較少,抑菌活性降低。隨著酶解溫度的增加,蛋白酶分子與底物分子之間自由碰撞概率增加,酶的催化效率也隨之增加,能夠促使底物產生更多的抗菌肽。酶解溫度過高,蛋白酶的結構可能被破壞,酶活性降低,甚至失活,不利于抗菌肽的生成。因此選擇40 ℃為最佳酶解溫度。
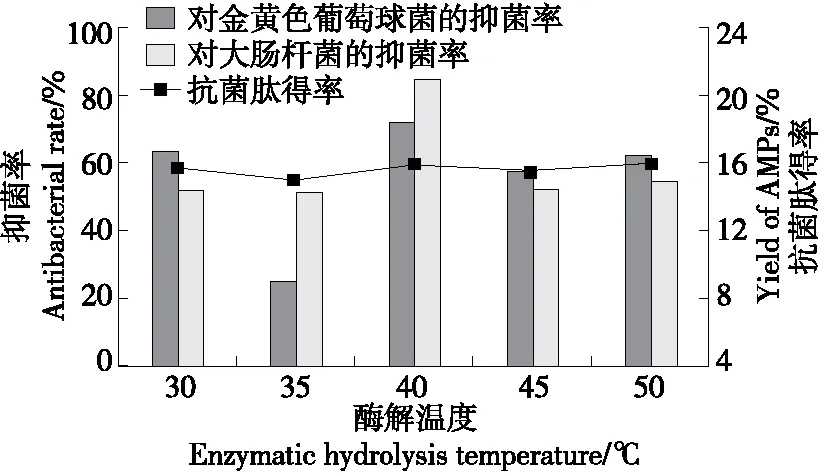
圖3 酶解溫度對虎奶菇菌絲體抗菌肽抑菌率及 得率的影響Figure 3 Effects of enzymatic hydrolysis temperature on antibacterial rate and yield of PTRM-AMPs
2.3.2 酶解時間 由圖4可知,當酶解時間為1~3 h時,虎奶菇菌絲體抗菌肽對金黃色葡萄球菌的抑菌率逐漸增加,在3~5 h酶解范圍內變化趨勢平緩。抗菌肽得率及對大腸桿菌的抑菌率在酶解1~5 h時先上升后下降,且在酶解3 h時達最大值,分別為17.67%和84.75%。繼續增加酶解時間,抗菌肽得率和抑菌率反而降低。這可能是因為更長的酶解時間使得具有抑菌活性的多肽被繼續降解為氨基酸,進而導致抗菌肽減少,抑菌率下降。故選擇3 h為最佳酶解時間。
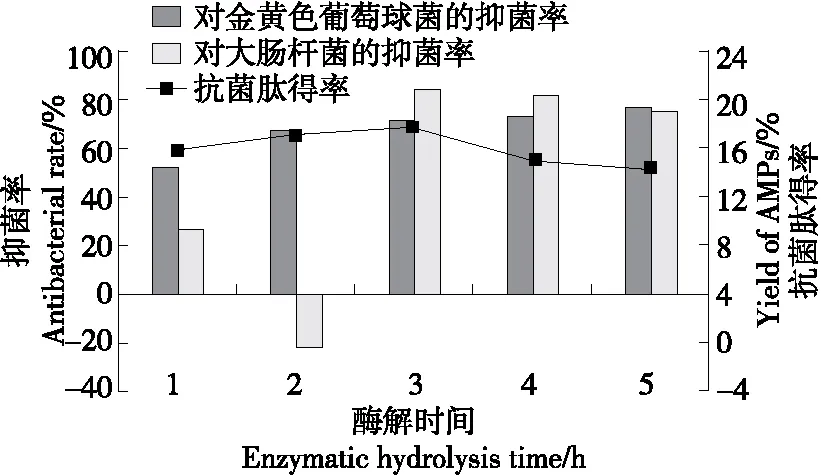
圖4 酶解時間對虎奶菇菌絲抗菌肽抑菌率及 得率的影響Figure 4 Effects of enzymatic hydrolysis time on antibacterial rate and yield of PTRM-AMPs
2.3.3 pH 由圖5可知,當pH為9.5~10.5時,抗菌肽得率無明顯變化,繼續增大pH,抗菌肽得率變大。這是因為堿性蛋白酶在堿性較高的環境下能夠發揮更好的作用,從而促進抗菌肽的生成。當pH為9.5~11.0時,抗菌肽對金黃色葡萄球菌的抑菌率隨pH的增大逐漸增加,并在pH為11.0時達最大值(74.03%),繼續增大pH,抑菌率反而下降,是因為過高的pH易導致蛋白酶活性降低,甚至失活。虎奶菇菌絲體抗菌肽對大腸桿菌抑菌率在pH為9.5~10.5時隨pH的增加逐漸增強,當pH為10.5,11.0時,抑菌率差異不顯著,分別為84.75%和84.54%。當pH>11.0時,抑菌率下降。因此選擇11.0為最佳酶解pH。

圖5 pH對虎奶菇菌絲體抗菌肽抑菌率及得率的影響Figure 5 Effects of pH on antibacterial rate and yield of PTRM-AMPs
2.3.4 料液比 由圖6可知,當料液比為1∶10~1∶20 (g/mL)時,虎奶菇菌絲體抗菌肽得率逐漸增加,抗菌肽對金黃色葡萄球菌和大腸桿菌的抑菌率也呈上升趨勢。這是因為料液比較低時,底物黏稠,不能充分接觸蛋白酶,不利于抗菌肽的提取。在一定水分含量范圍內,隨著料液比的增加,溶劑完全浸潤樣品,底物與蛋白酶充分接觸,產生更多具有抗菌活性的多肽片段[26]。當料液比從1∶20 (g/mL)繼續增加至1∶30 (g/mL)時,抗菌肽得率雖有所增加,但抑菌率降低,可能是隨著料液比的進一步增加,樣品中其他水溶性物質溶解度增加,抗菌肽抑菌率下降,甚至促進大腸桿菌的生長。因此,綜合考慮,選擇1∶20 (g/mL)為最佳料液比。
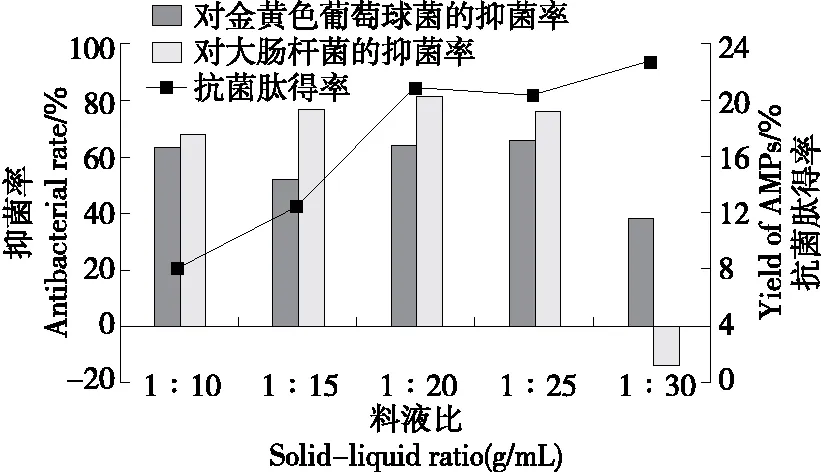
圖6 料液比對虎奶菇菌絲體抗菌肽抑菌率及 得率的影響Figure 6 Effects of solid-liquid ratio on antibacterial rate and yield of PTRM-AMPs
2.3.5 酶添加量 由圖7可知,抗菌肽對金黃色葡萄球菌的抑菌率在加酶量為6 000~8 000 U/g范圍內平緩增加,在8 000~14 000 U/g范圍內呈下降趨勢。而抗菌肽得率和對大腸桿菌的抑菌率在加酶量為6 000~14 000 U/g時持續下降,是因為蛋白酶和底物的比例達到平衡時,能夠生成較多的抗菌肽。蛋白酶添加量繼續增加,底物質量保持不變,則會使得生成的抗菌肽進一步水解成活性較低的小分子肽或者無活性的氨基酸,抗菌肽含量及抑菌率下降。為節約成本,選擇6 000 U/g為最佳酶添加量。
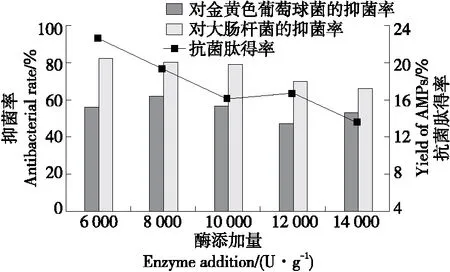
圖7 酶添加量對虎奶菇菌絲體抗菌肽抑菌率及 得率的影響Figure 7 Effects of enzyme addition on antibacterial rate and yield of PTRM-AMPs
2.4 響應面優化
2.4.1 響應面試驗設計與結果 根據單因素試驗結果,固定料液比為1∶20 (g/mL),酶添加量為6 000 U/g,以酶解時間、pH和酶解溫度為自變量,以抗菌肽對金黃色葡萄球菌和大腸桿菌的抑菌率為響應值,根據Design-Expert 10.0.7軟件中的Box-Behnken中心組合設計原理進行響應面設計,因素水平見表2,試驗設計及結果見表3。
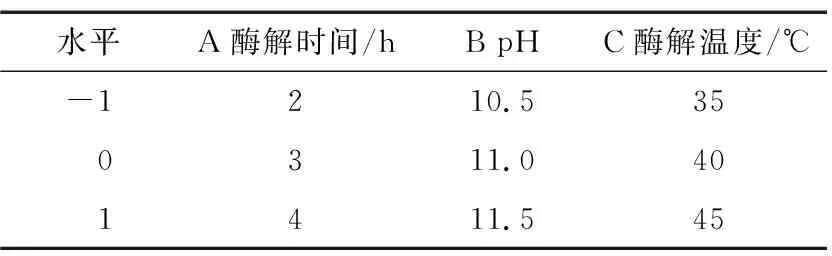
表2 響應面試驗的因素和水平Table 2 Factors and levels of response surface test
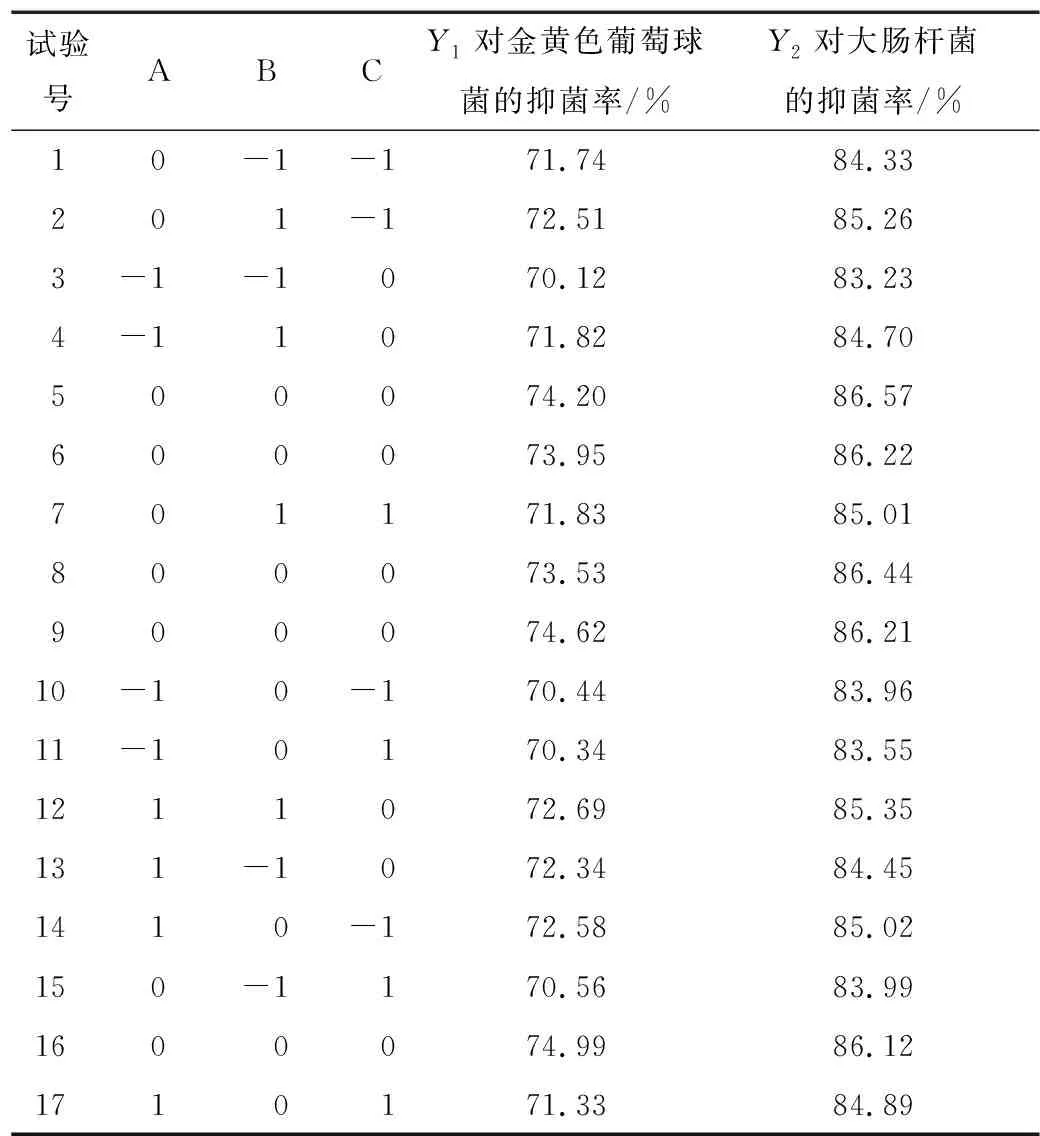
表3 響應面試驗設計及結果Table 3 Experimental design and results for response surface methodology
2.4.2 響應面模型的建立及方差分析 通過Design-Expert 10.0.7軟件對試驗結果進行多元回歸擬合,得回歸模型方程:
Y1=74.26+0.78A+0.51B-0.40C-0.34AB-
0.29AC+0.12BC-1.50A2-1.01B2-1.58C2,
(3)
Y2=86.31+0.53A+0.54B-0.14C-0.14AB+0.07AC+0.023BC-1.09A2-0.79B2-0.87C2。
(4)


表4 回歸模型方差分析(Y1)?Table 4 Variance analysis of regression model (Y1)
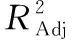
2.4.3 響應面交互作用分析 由圖8、圖9可知,酶解時間和pH的交互作用對虎奶菇菌絲體抗菌肽抑菌率的影響最顯著,酶解時間、pH和酶解溫度3個因素中任意兩個因素之間的交互作用均不明顯,與表4、表5中方差分析結果相吻合。
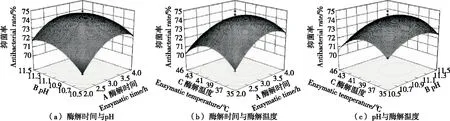
圖8 各因素交互作用對虎奶菇菌絲體抗菌肽抑菌率的影響(金黃色葡萄球菌)Figure 8 Effects of the interaction of various factors on antibacterial rate of PTRM-AMPs (Against S.aureus)

圖9 各因素交互作用對虎奶菇菌絲體抗菌肽抑菌率的影響(大腸桿菌)Figure 9 Effects of the interaction of various factors on antibacterial rate of PTRM-AMPs (Against E.coli)

表5 回歸模型方差分析 (Y2)?Table 5 Variance analysis of regression model (Y2)
2.4.4 雙指數最優提取條件的確定及驗證 根據模型分析結果及試驗操作的可行性,將最佳酶解條件確定為酶解時間3.2 h、pH值11.1、酶解溫度39.5 ℃,此時虎奶菇菌絲體抗菌肽對金黃色葡萄球菌和大腸桿菌抑菌率的預測值分別為74.43%和86.46%。為了驗證該模型的有效性,連續進行3組重復實驗,測得該條件下制備的虎奶菇菌絲體抗菌肽對金黃色葡萄球菌和大腸桿菌的抑菌率平均值分別為74.17%和86.22%,與模型預測結果接近。因此,此模型準確可行。此時,抗菌肽得率為22.67%,比優化前提高了5.62%。
2.5 最小抑菌濃度(MIC)
由圖10可知,抗菌肽對金黃色葡萄球菌和大腸桿菌的MIC值分別為5.0,2.5 mg/mL。馬尚玄等[23]采用堿性蛋白酶酶解制備的澳洲堅果多肽MNP-8對金黃色葡萄球球菌和大腸桿菌的MIC分別為4.0,4.5 mg/mL,與試驗結論相似。因此天然虎奶菇菌絲體抗菌肽具有廣譜抑菌活性,且抑菌作用較強,有望成為抗生素和食品防腐劑的替代品。
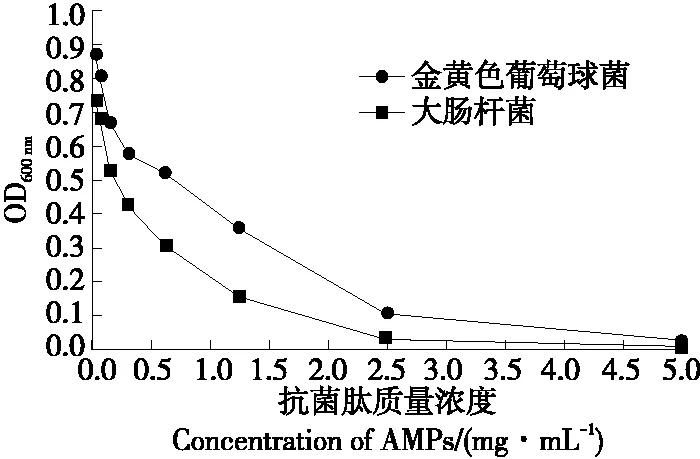
圖10 虎奶菇菌絲體抗菌肽的最小抑菌濃度Figure 10 Minimum inhibitory concentration of PTRM-AMPs
3 結論
堿性蛋白酶酶解制備虎奶菇菌絲體抗菌肽的最佳工藝條件為酶解時間3.2 h、酶解溫度39.5 ℃、pH值11.1,此時虎奶菇菌絲體抗菌肽對金黃色葡萄球菌和大腸桿菌的抑菌率分別為74.17%和86.22%,抗菌肽得率為22.67%。抗菌肽對金黃色葡萄球菌和大腸桿菌的最低抑菌濃度分別為5.0,2.5 mg/mL。后續可對虎奶菇菌絲體抗菌肽進行分離純化,并研究其作為抗生素及食品防腐劑的潛在可能,從而提高虎奶菇菌絲體抗菌肽的利用價值。
