寧夏河東沙地生物土壤結皮對土壤性質及入滲過程的影響
2022-09-26 11:47:36董智今張呈春展秀麗
生態學報 2022年17期
董智今,張呈春,展秀麗
寧夏大學地理科學與規劃學院, 銀川 750021
生物土壤結皮(BSCs)是細菌,真菌,地衣,藻類和苔蘚等生物同表層土壤顆粒相互作用形成的復合體[1—2],是干旱區地表主要的覆蓋形式[3—4],其存在改變了土壤理化性質[5—7]、土壤生態[8]及水文過程[9],對荒漠生態系統的物質與能量流動產生重要影響。隨著生物土壤結皮的發育與演替,其下伏土壤對降水的再分配和儲存利用方式發生明顯改變[1,10—11],進而形成了有別于原有荒漠生態系統的獨特的生態水文過程[2—3,12]。研究沙區不同生物土壤結皮對土壤性質以及水分入滲的影響,對干旱區生態系統修復與保護意義重大。
近年來,眾多學者開展了關于生物土壤結皮對土壤影響的研究,主要包括了不同類型(發育程度)生物土壤結皮對土壤理化性質[5—6,13—17]、水分入滲[10—12,18—20]、水分動力學[18,21—22]、水汽凝結與蒸發[23]等的影響研究,以及凍融交替[24]、降水沖刷[25]、踩踏[4,20]、植物根系[21,26—27]等干擾對生物土壤結皮滲透性影響的研究,研究區重點集中在黃土高原[8, 18,21—23]、青藏高原[28—29]、騰格里沙漠[13—14,30—31]、毛烏素沙地[11,15]等生態脆弱區,其中生物土壤結皮能夠顯著影響土壤性質已成為研究共識[2,13—17],但有關生物土壤結皮對土壤入滲影響的研究結果仍不盡相同,例如由于受多種氣候以及微環境等因素的影響,生物土壤結皮具有發育復雜、空間異質性等特點,使得其對土壤水分入滲產生促進[27—28,31]、阻礙[18,30]和無顯著作用[25]這三方面的影響,因此,定量化描述不同區域生物土壤結皮對水分入滲的影響仍是目前研究的重點。本研究區固沙植被主要為灌木、生物土壤結皮與一年生草本植物,其中發育晚期的蘚類結皮與草本植物混生代表著沙區植被-土壤系統處于良好的恢復和相對穩定的階段,這增加了沙區土壤生態水文過程的多樣性與復雜性,因此研究草本與生物土壤結皮混生對土壤水分入滲過程的影響具有重要意義。
寧夏河東沙地一直是國家和寧夏生態治理的重點區域,近年來,該地區防沙治沙工作取得了重大效果,地表覆被面積增加顯著。但是目前有關該區域生態系統的深層研究較為薄弱,特別是沙漠化治理過程中生物土壤結皮的形成及覆蓋對下伏土壤性質以及對土壤水分再分配的影響研究較少。本文選擇由不同類型生物土壤結皮(藻類結皮、蘚類結皮和草本-蘚類結皮)覆蓋下的風沙土以及流動沙丘作為對照,采用雙環入滲試驗和模型模擬,對3種生物土壤結皮和裸沙的土壤性質和入滲特征展開研究,以期為該區域建立合理穩定的固沙植被,實現沙區生態系統的修復與重建提供基礎數據與理論借鑒。
1 研究地區與研究方法
1.1 研究區概況
寧夏靈武市白芨灘(37°49′05″—38°20′54″N,106°20′22″—106°37′19″E)位于毛烏素沙地西南緣,面積748.43 km2。該保護區屬于中溫帶干旱氣候區,夏季炎熱干燥,冬季寒冷,年均溫10.4 ℃,年均降水量206.2—255.2 mm,年均蒸發量1933.3 mm。研究區地帶性土壤以灰鈣土為主,非地帶性土壤主要為風沙土,土壤結構松散。植被以一年生草本植物、沙地植被和荒漠植被為主,代表性植物群落有檸條(Caraganakorshinskii)、貓頭刺(Oxytropisaciphylla)、花棒(Hedysarumscoparium)等。生物土壤結皮是該地區固定與半固定沙丘常見的土表微景觀,研究區內生物土壤結皮在固定沙丘的迎風坡上優先發育,丘間低地廣泛發育斑塊狀蘚類結皮,坡頂與迎風坡灌叢植被間有連片發育的藻類結皮。研究區藻類與蘚類結皮的優勢種為具鞘微鞘藻(Microcoleusvaginatus)、真蘚(Bryumargenteum)[32]。
1.2 試驗方案
2021年10月,在白芨灘自然保護區選擇地勢較平坦、生物土壤結皮覆蓋度廣(>90%)的區域作為試驗觀測點,分別為無結皮覆蓋的裸沙(BS)、發育早期藻類結皮覆蓋土壤(AC)、發育晚期蘚類結皮覆蓋土壤(MC)以及蘚類結皮與草本植物混生覆蓋土壤(H-MC),每個試驗點面積為25 m2(5 m×5 m)。其中位于固定沙丘頂的AC樣地中藻類結皮相對蓋度>85%,結皮厚度為2.95—3.47 cm,表面較光滑,顏色為淺灰色;位于丘間平地的MC樣地中蘚類結皮相對蓋度>80%,結皮層厚度為10.08—11.48 cm,表面較粗糙,顏色較深;H-MC樣地為檸條灌叢下高覆蓋度(>85%)的蘚類結皮,地表生長一年生雜草(覆蓋度為60%)。
土壤水分入滲采用雙環入滲儀(QT-IN12-W,北京渠道科學儀器有限公司,中國)測定,雙環入滲儀內環直徑為20 cm,外環直徑為35 cm,高度為35 cm。在試驗點內,針對AC、MC和H-MC,分別選擇藻類結皮、蘚類結皮和草本-蘚類結皮完全覆蓋的區域(結皮覆蓋面直徑> 35 cm)作為入滲試驗樣點,將雙環放置在樣點內垂直地面下埋15 cm,并使用大孔隙單層紗布覆蓋保護表層結皮。由于本研究區沙土初始入滲過快,儀器所配馬氏瓶不能滿足供水需求,因此使用2個1000 ml標準量筒分別向內環和外環同時均勻供水,當內外環中的水層高度同時達到3 cm時[33],開始計時并讀取內環相應時間內增加的水量。記錄不同時段的入滲水量:0—2 min每隔30 s記錄1次,2—12 min每隔1 min記錄1次,14—22 min每隔2 min記錄1次,27—67 min每隔5 min記錄1次,試驗過程中內外環水面固定保持在3 cm高度以此來保持水頭穩定[33]。當至少3次相同測定時間內供水量恒定時可視為達到穩定[22,33],停止試驗,測定入滲時間不少于65 min。
由于本研究區沙土入滲過快,野外試驗中初始入滲量存在誤差,參考前人研究[10—11,21—22],本文將入滲前3 min的入滲速率定義為初始入滲速率;將單位時間內的入滲量趨于穩定時的入滲速率定義為穩定入滲速率;將達到穩滲時的累積入滲量與達到穩滲所用時間的比值定義為平均入滲速率;將60 min內的累積入滲總量定義為累積入滲量[22],入滲速率計算公式如下:
(1)
式中:Vn為一定溫度下n時刻的土壤入滲速率(mm/min);Ln為n時刻的內環供水量(cm3);S為內環面積(cm3);10為cm與mm的轉換系數;Δn為觀測間隔時間差(min);T為時段內的平均水溫(10 ℃)。
1.3 樣品采集
采用烘干法測定土壤含水量[23],入滲試驗前在每個樣地用鋁盒分別采集0—1 cm、1—10和10—20 cm深處土壤樣品,每個深度重復取三次,共取鋁盒樣品36個;采用環刀法測定土壤容重[10],入滲試驗前在每個樣地分別取表層,10 cm和20 cm處取環刀樣品,每個深度重復取三次,共取環刀樣品24個;采用游標卡尺測量生物土壤結皮厚度(取3—5次重復)[21]。
入滲試驗前在樣地內選取3—5個采樣點進行分層采樣,分層為生物土壤結皮層(0—1 cm)和結皮覆蓋下土壤(1—10和10—20 cm),同層樣品進行混合并裝袋后帶回室內備用。
1.4 實驗室分析
土壤飽和含水量與土壤孔隙度在室內測定[34];采用重鉻酸鉀外加熱法測定土壤有機質含量[16];馬爾文激光粒度儀(Mastersizer 3000,Malvern Instruments Ltd.,UK)測定土壤機械組成。
1.5 土壤水分入滲模型
模型模擬公式選取Philip、Kostiakov和Horton 3種常用的入滲模型[35],對入滲過程模擬并評價其適用性。
Philip模型:
f(t)=0.5s/t1/2+A
(2)
Kostiakov模型:
f(t)=b/ta
(3)
Horton模型:
f(t)=fc+(f0+fc)/ekt
(4)
式中:f(t)為入滲速率(mm/min);f0為初滲速率(mm/min);A和fc為穩滲速率(mm/min);t為入滲時間(min);s為吸濕率(mm/min1/2),表征土壤入滲能力;a,b為常數,分別表征第一個單位時間段內的平均入滲速率與入滲速率隨時間減小的程度;k為參數,表征入滲速率隨時間減少的程度。
1.6 數據處理
使用Microsoft Excel 2019對數據進行處理與分析,IBM SPSS 24. 0進行單因素方差分析(one-way ANOVA)、Pearson相關分析(α=0.05)和逐步回歸分析(α=0.1),通過Origin 2018作圖。
2 結果與分析
2.1 不同生物土壤結皮對土壤性質的影響
從表1可知,研究區0—20 cm藻類、蘚類、草本-蘚類3種類型生物土壤結皮覆蓋下土壤以及裸沙的機械組成均以砂粒為主(>90%)、粉粒含量較少(<9%)、黏粒含量極少或無(<0.02%)。生物土壤結皮覆蓋的表層土壤與裸沙相比,粉粒含量增多(3.6%—5.8%);極細砂粒含量增多(0.4%—1.7%);細砂含量減少(7.8%—33.7%)且蘚類結皮的細砂粒含量顯著減少;中砂含量增多(2.6%—20.9%);粗砂含量增多(1.2%—8.1%)且草本-蘚類結皮的粗砂含量顯著增加。生物土壤結皮的存在,使得土壤粒徑分布更加均勻:粉粒、極細砂、中砂和粗砂含量均有所增加,細砂含量減少。
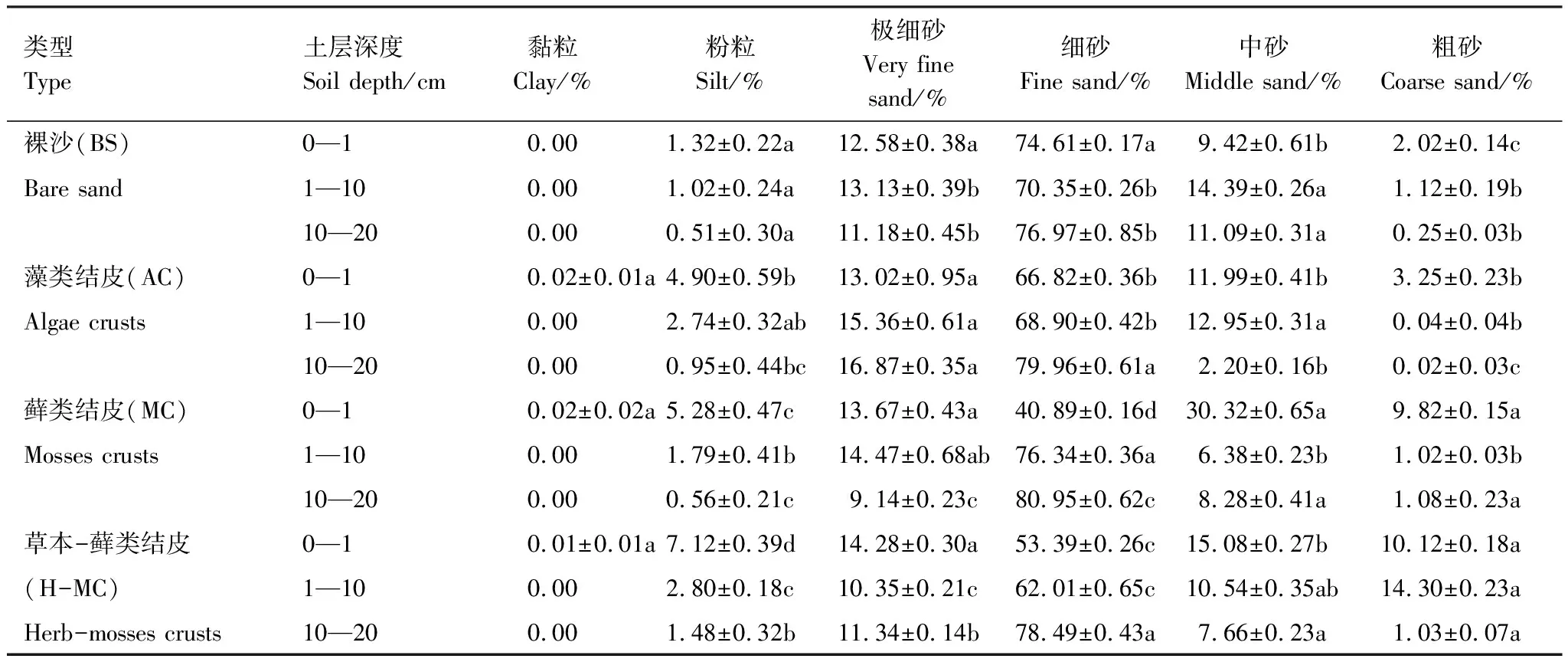
表1 不同發育程度生物土壤結皮覆蓋下土壤的顆粒組成
如表2所示,隨著生物土壤結皮的發育,結皮厚度顯著增加。與裸沙相比,藻類與蘚類結皮覆蓋下土壤容重變化不顯著,蘚類結皮初始含水量增加了2.4倍,藻類結皮初始含水量變化不顯著,土壤總孔隙度分別減少了4.0%和6.5%,飽和含水量分別減少了6.8%和11.1%,土壤有機質含量是裸沙的5—6倍。草本-蘚類結皮覆蓋下土壤與裸沙相比有機質增加了6倍,總孔隙度增加了9.1%,飽和含水量增加了12.6%,初始含水量變化不顯著。
2.2 不同生物土壤結皮覆蓋下土壤入滲特征
由圖1可知,不同生物土壤結皮覆蓋下土壤水分入滲速率均呈現出先降低再波動最終趨于穩定。4種樣地入滲速率之間差異顯著(P=0.036<0.05),累計入滲量表現為H-MC>BS>AC>MC,表明發育程度更高的蘚類結皮能夠顯著降低土壤入滲特性,而草本與蘚類結皮混生的復合表層,由于植物根系的存在入滲速率與累計入滲量顯著增加。
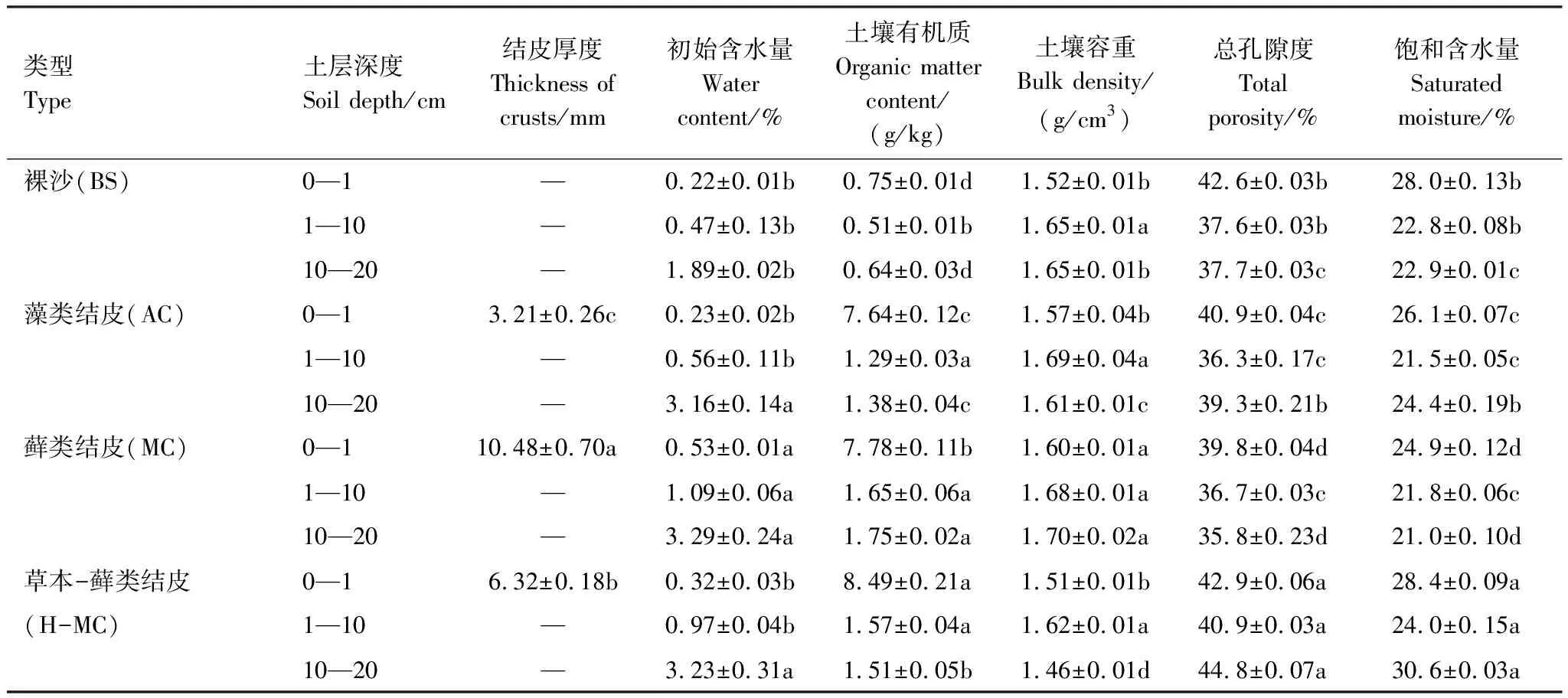
表2 不同發育程度生物土壤結皮覆蓋下的土壤性質
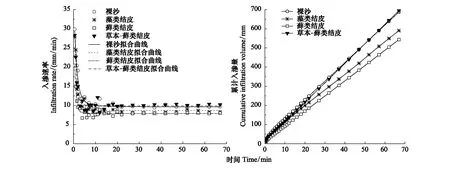
圖1 不同發育程度生物土壤結皮覆蓋下土壤的水分入滲過程與累計入滲量Fig.1 Infiltration rate /cumulative infiltration volume of the biocrusts in different developmental degrees
由圖1可知,土壤入滲可分為初期瞬減、中期波動和后期穩定三個階段。如表3所示,第一階段,裸沙初滲速率顯著高于有結皮覆蓋的土壤 (P<0.05),表明藻類、蘚類和草本-蘚類結皮的存在明顯降低了初滲速率,降低幅度依次為14.3%、37.2%、11.8%;第二階段入滲速率變化劇烈(圖1),裸沙的入滲速率仍然顯著高于發育程度較高的蘚類結皮;第三階段,由于此時土壤孔隙被水分充滿達到飽和進入穩滲階段,與裸沙相比,藻類和蘚類結皮覆蓋下土壤達到穩定入滲的時間更長,表明藻類和蘚類結皮能明顯降低土壤穩滲速率,降低幅度分別為14.4%和18.3%,草本-蘚類結皮覆蓋下土壤入滲達穩定的時間則變短,穩滲速率較裸沙增加了4.5%。表明生物土壤結皮的發育程度越高,阻止土壤水分的入滲能力越強,而草本-蘚類結皮復合體則會促進土壤水分入滲。
2.3 影響土壤入滲性能的因素
如表4所示,土壤入滲性能與土壤理化性質的相關性表現為:初滲速率與結皮厚度和初始含水量呈顯著負相關,與其余因素相關性不顯著;穩滲速率、累計入滲量均與土壤容重呈現極顯著負相關,與土壤總孔隙度以及飽和含水量呈極顯著正相關,與結皮層厚度、土壤機械組成和有機質相關性不顯著;平均入滲速率與土壤容重呈顯著負相關,與總孔隙度與飽和含水量呈顯著正相關,與其他因素相關性不顯著。
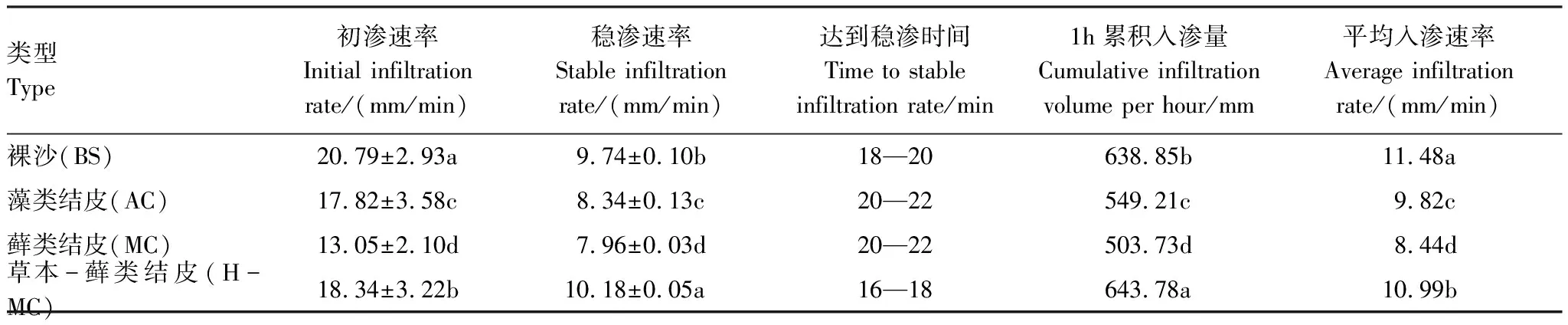
表3 不同發育程度生物土壤結皮覆蓋下土壤的水分入滲參數
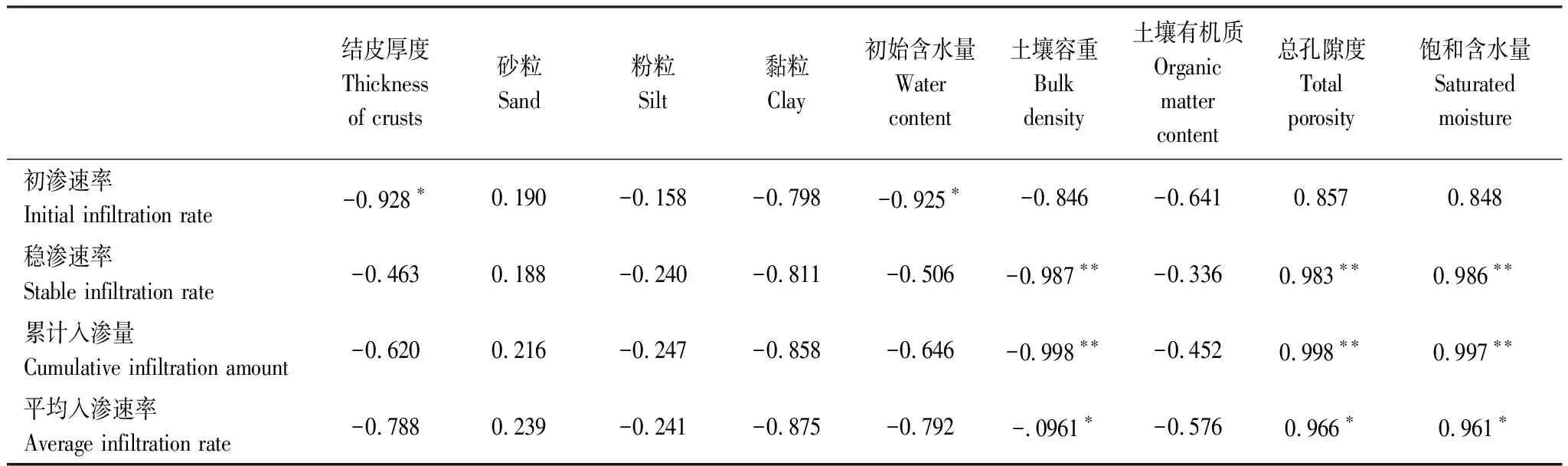
表4 土壤入滲性能與影響因素的相關性
在此基礎上分別對初滲速率與穩滲速率進行逐步回歸分析可得出:
Ii= -2.066 TOC+14.197SM+12.676BD+17.500 (R2=0.999)
(5)
Si= -1.202BD+0.225WC+ 0.00125C+9.055 (R2=0.999)
(6)
式中:Ii為初滲速率(mm/min);Si為穩滲速率(mm/min);TOC為結皮厚度(mm);SM為飽和含水量(%);BD為土壤容重(g/cm3);WC為初始含水量(%);C為黏粒含量(%)。
由方程(5)可知,初滲速率主要受結皮厚度、飽和含水量和土壤容重影響,結皮層越厚、飽和含水量越低與土壤容重越大均會阻礙土壤的初始入滲能力;由方程(6)可知,穩滲速率主要受土壤容重、初始含水量和黏粒含量影響,土壤容重越大、初始含水量越低、黏粒含量越少則會降低土壤的穩定入滲能力。
2.4 不同生物土壤結皮覆蓋下土壤入滲過程模擬
由表5可以看出,不同模型對于土壤水分入滲過程的擬合效果不同。Philip模型擬合結果顯示,擬合A(穩滲速率)值為4.86—6.65 mm/min與實際的7.96—10.18 mm/min相差較大,R2均小于0.89,說明Philip模型擬合效果較差。Horton模型擬合結果顯示,fc(穩滲速率)值與實測值相差很小(0.05—0.315 mm/min),k值在0.79—1.54之間,最大值出現在裸沙,最小值在蘚類結皮,反映了入滲速率遞減情況,與實測結果基本相符,R2均大于0.94,模型擬合效果較好。Kostiakov模型擬合結果顯示,參數b值在7.74—11.72 mm/min之間,b值越大,入滲曲線的斜率越大,瞬時入滲率衰減越快,擬合出衰減快慢順序為:BS>H-MC>AC>MC,與實測結果一致,R2均大于0.96,此模型擬合效果較好。
3 討論
3.1 生物土壤結皮覆蓋對土壤性質的影響
生物土壤結皮通過改變地表微形態使其下伏土壤的性質發生變化[5]。首先,不同發育程度的生物土壤結皮對下伏土壤機械組成影響不同。本研究發現,與藻類結皮相比,蘚類結皮表層土壤砂粒含量降低,粉粒含量增加,土壤結構細化,顆粒物分布更均勻,這是因為在蘚類結皮以及草本-蘚類結皮混生覆蓋下,蘚類與草本使得土壤表面粗糙度加大,對于細顆粒物的攔截作用明顯提高,這與高廣磊等[15]在寧夏鹽池縣以及孫福海等[18]在黃土高原的研究結果一致。
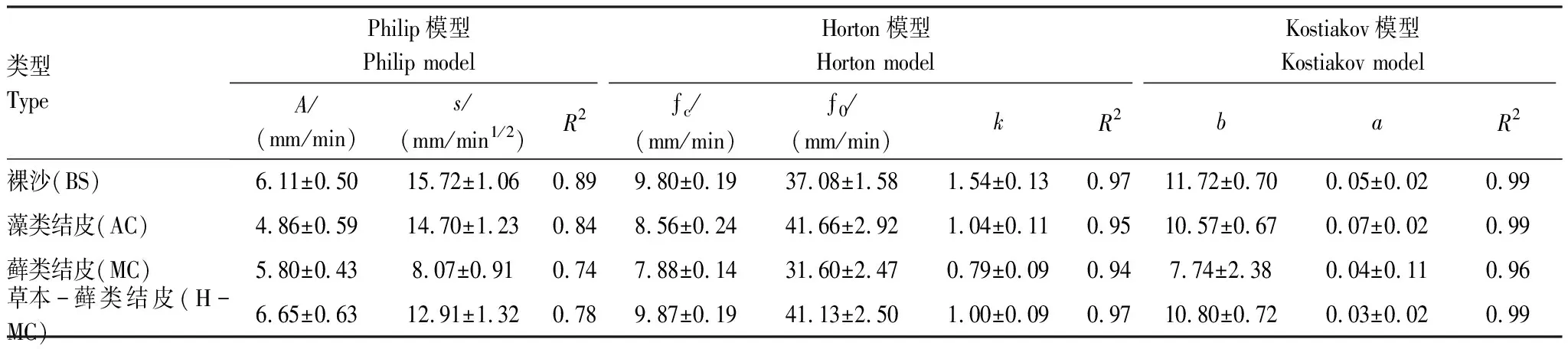
表5 不同發育程度生物土壤結皮覆蓋下土壤入滲過程的模型模擬結果
其次,不同發育程度的生物土壤結皮對下伏土壤總孔隙度和土壤含水量影響不同。本研究發現草本-蘚類結皮覆蓋下土壤總孔隙度較高,說明草本植物根系對結皮下伏土壤的影響顯著。蘚類結皮覆蓋下土壤表層含水量最高,這是因為在本研究區蘚類結皮廣泛分布于丘間低地,灰黑色藻類結皮分布于固定沙丘坡面-丘頂,蒸發較強,而草本-蘚類結皮則是由于生長季植物需水,使得下伏土壤表層含水量與裸沙相比差異較小。
最后,不同發育程度的生物土壤結皮對下伏土壤有機質影響不同。本研究發現藻類和蘚類結皮的覆蓋使得表層土壤的有機質含量增加,一方面歸因于生物土壤結皮的覆蓋使表層土壤結構優化,利于土壤團聚體形成,增加有機質含量[5];另一方面則由于生物土壤結皮可以為微生物的生長提供環境,進而提升下伏土壤的有機質含量[13]。
此外,生物土壤結皮對土壤性質的影響還具有“表層性”[36],即生物土壤結皮和下伏土壤的雙層結構,使得結皮對土壤性質的影響主要集中在表層[17,21],這在本研究中也得到了證實。
3.2 生物土壤結皮覆蓋對土壤水分入滲的影響
本研究發現藻類和蘚類結皮明顯阻礙了土壤水分入滲過程,且生物土壤結皮發育程度越高,土壤滲透性能越低。這是因為,第一,生物土壤結皮截留的細粒物質填充土壤孔隙,降低了孔隙間的連通性[37—38];第二,本研究區內高蓋度藻類和蘚類結皮中的藻絲體、真菌菌絲、蘚類假根與砂粒纏繞膠結增加了結皮層厚度[22],結皮層吸水性增強進而阻礙水分的入滲[21],這與Xiao等[37]研究結論一致,并且與裸沙相比,蘚類結皮覆蓋下表層土壤初始含水量明顯增加,降低了沙土的土壤水力梯度,進而減緩了初滲速率[19];第三,生物土壤結皮覆蓋增加了表層土壤有機質,且蘚類植物體分泌的胞外多糖等化學物質堵塞土壤孔隙進而降低入滲速率[11]。最后,本研究中蘚類較藻類結皮表現出更高的阻滲能力,這與Zhang等[14]研究結果不同,因為本研究選取地勢平坦區域作為觀測試驗點,無地形因素對結皮層的影響,而且本研究發現蘚類結皮能夠完全覆蓋表土層孔隙,蘚類植物的毛尖、莖葉微結構使得土壤更封閉疏水性更強,這對入滲的阻礙作用更顯著[26]。
本研究中還發現草本-蘚類結皮復合體能夠有效的促進土壤水分入滲,這是因為,草本植物為發育在檸條灌叢下一年生的雜草,受到雜草和檸條根系的影響,表層結皮層被破壞,水分運移方式被改變[39],同時,植物根系在土壤中交錯生長分布,形成土壤大孔隙,提高了土壤的滲透性能[40]。李淵博等[21]在黃土高原的研究也表明草本植物和蘚類結皮混生覆蓋下會對水分入滲有一定的促進作用,與本研究結果一致。
3.3 土壤入滲性能差異的因素分析
本研究發現,不同發育程度的生物土壤結皮入滲性能的差異與土壤性質密切相關,并且在不同的入滲階段,土壤性質對其影響也各不相同。在入滲的初始階段,結皮層越厚,其吸水性越強,入滲速率也越低,而且初始階段土壤初始含水量越多,水力梯度越小入滲速率也越低[19]。在本研究中蘚類結皮的覆蓋,明顯提升了結皮層厚度與土壤初始含水量,從而減小初滲速率,這與王閃閃等[41]研究結果一致。達到穩滲階段時,入滲速率與土壤容重、總孔隙度與飽和含水量呈現極顯著相關性,在該階段,生物土壤結皮對入滲的影響減弱,而下伏風沙土的性質以及植物根系產生的裂隙則成為影響入滲的主要因素[39,42]。
本研究進行逐步回歸分析后發現,結皮層厚度是影響初滲速率最重要因素,而土壤容重則是影響穩滲速率最重要因素,這進一步說明,入滲時間越長,生物土壤結皮對入滲的影響越小,而土壤性質以及植物生長產生的土壤裂隙對入滲的影響越大。
3.4 入滲模型適用性評價
本研究中采用了三種經典的入滲模型來模擬生物土壤結皮覆蓋下水分入滲規律,結果發現,Kostiakov模型對不同樣地入滲過程的模擬效果最好(R2>0.96),擬合出入滲速率的快慢為:BS>H-MC>AC>MC,與實測結果一致。而Philip模型是物理模型,與有結皮覆蓋的土壤相比,裸沙的水分入滲過程擬合效果更好(R2=0.89),說明該模型不適用于模擬生物土壤結皮覆蓋下的非均質土壤,更適用于均質土壤的一維入滲[10]。 Horton模型屬于純經驗公式[15],對4種樣地類型擬合效果均較好(R2>0.94),對初始入滲速率值擬合精度高,但是對穩定入滲速率的擬合精度不夠。
綜上所述,研究區內生物土壤結皮覆蓋改變了下伏土壤的性質,進而影響了土壤水分入滲過程。在對該區域生物土壤結皮的研究中,未來還需要深入探討固沙植物根系對水分入滲過程的影響,進一步開展生物土壤結皮對該區域土壤水文過程的研究。
4 結論
(1)生物土壤結皮對淺層土壤性質影響明顯。相較裸沙,生物土壤結皮覆蓋下的土壤砂粒含量減少,粉粒含量增加,土壤有機質含量增加;蘚類結皮土壤初始含水量最高;藻類和蘚類結皮覆蓋下土壤總孔隙度與飽和含水量降低,草本-蘚類結皮其下伏土壤總孔隙度與飽和含水量增加。
(2)生物土壤結皮改變下伏土壤性質進而對風沙土的入滲性能夠產生顯著影響。生物土壤結皮發育程度越高,阻止土壤水分的入滲能力越強,而草本-蘚類結皮復合體則促進土壤水分入滲;在入滲初期,結皮層厚度、土壤初始含水量是影響入滲的主要因素,而在穩滲階段,土壤容重則是影響入滲特征的主要因素,且植物根系的存在減小了生物土壤結皮的阻滲作用。
(3)Kostiakov模型最適用于模擬本研究區不同發育程度生物土壤結皮覆蓋下土壤水分入滲過程,Horton模型次之,Philip模型較差。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
科技傳播(2019年22期)2020-01-14 03:06:54
