內蒙古東北段森林衰退現狀及種群競爭對其生長的影響
2022-09-26 12:21:44田育紅劉怡青吳秀臣顏孟萱劉鳳花欒靜雅
生態學報 2022年17期
關鍵詞:生長
田育紅,劉怡青,吳秀臣,顏孟萱,劉鳳花,欒靜雅
北京師范大學地理科學學部,地表過程與資源生態國家重點實驗室, 北京 100875
森林覆蓋著全球約30%的陸地表面[1],提供了大量的木材、纖維及燃料等產物,對于人類生產生活的各個方面有不可替代的作用,森林生態系統同樣也對于調節全球碳平衡、改善大氣、水、土壤等生態環境等有重要意義,其生態系統服務功能具有很高的價值[2]。森林衰退來自于多種脅迫的共同作用,具有隨機性與復雜性,它是指森林(樹木)在生長過程中表現出生產力降低、生理機能及生長活力下降、生長緩慢停滯或死亡以及地力衰退等狀態,主要表現為林木生長力衰減甚至出現死亡[3]。當前森林衰退現象有向全世界蔓延的趨勢,廣泛引起各國領導層的關注和生態學家們的重視。大規模的森林衰退會改變生態系統物種組成、營養物質循環和生物量的積累周期,進而影響森林的更新和結構,對森林生態系統的服務功能和生物多樣性造成難以恢復的干擾[4],一定程度上對人類社會、文化和生態功能產生影響,降低可持續發展性[5]。
種群競爭是森林衰退的內在驅動因子。植物個體間的競爭作用對植物各方面活動比如生長、存活和繁殖都有影響[6]。競爭本質是個體之間為爭奪資源而抑制對方的現象,可能會致使一方受到抑制甚至逐漸衰亡[7]。特別是當資源有限時,植物之間的競爭作用會導致植物種群存活率和生長率的降低[8]。Fernández-de-Ua等[9]對樟子松(Pinussylvestris)、比利牛斯櫟(Quercuspyrenaica)與葡萄牙櫟(Quercusfaginea)的種間競爭及其與樹木生長、氣候變化的關系進行研究后,發現樹木生長隨著競爭的加劇呈指數級下降。由于氣候變化引發的資源利用率降低、水勢下降、光合速率和氣孔導度降低,地上地下的競爭也越發激烈,最后降低了個體的徑向生長。低競爭水平下的樹木將能更好地抵御氣候變化情景下預測的變暖條件,而高競爭水平下的林木個體在持續增強的干旱事件中難以存活。因此,競爭強度影響著樹木在生長過程中對氣候響應的變異性[10]。
另外,氣候變化的背景下,物種競爭加劇,進而促使生物群落的變化,改變物種分布和森林結構。樹木死亡率的增加不一定歸因于全球變化型干旱,也可能是由于氣候影響樹木競爭所致[11]。相關研究指出森林組分變動主要通過兩種途徑實現:一是原樹種和更適應于氣候變化的樹種(或入侵種)之間產生競爭而后被逐漸替代,導致森林結構發生變化[12];二是原樹種通過干早引起的枯死或野火燒毀發生死亡并被替代[13]。在歐洲西北部、南美等地的森林,入侵的喜溫植物取代了原有物種而引發了森林逐步退化的現象[14—16]。劉丹等[17]的研究結果顯示,分布在大興安嶺的興安落葉松(Larixgmelinii)及小興安嶺和東部山地的云杉(Piceaasperata)、冷杉(Abiesfabri)和紅杉(Larixpotaninii)等樹種的最適分布范圍均發生了北移[18]。Luo[19]通過鄰域分析對中西部加拿大北方森林進行研究發現:區域變暖加劇了樹木的競爭。區域變暖可能通過延長生長季節和增加土壤養分利用率來刺激寒冷地區的樹木生長[20—21],這反過來可能增加了樹木的競爭。總而言之,在全球氣候變化包括CO2的增加導致的升溫等環境因素的影響下,森林中的樹種如果不能及時有效地改變基因和生理機制去適應變化后的氣候,就很有可能在競爭和分異中逐漸衰退[22]。廣泛的研究結果顯示,種群競爭對森林衰退的影響具有一致性。當前對于種群競爭引起森林衰退的潛在影響機制知之甚少,這是氣候學家和生態學家研究的重點方向之一[23],這也是干旱半干旱區森林衰退影響機制當前亟待解決的難題之一。
內蒙古東北段近百年來溫度增加趨勢顯著[24],氣候異常事件頻發,主要表現為升溫和降水的減少[25—26]。同時在該區域觀測到,在低海拔區域和北方森林地區南部有大量森林衰退現象[27—28]。這些區域已成為研究森林衰退的重點區域。然而,對該區域森林種群競爭引起森林衰退的影響機制仍缺乏足夠的研究。本研究選取中國半干旱區東段內蒙古大興安嶺的典型森林樣地為研究對象,選擇合適的競爭指數模型并結合樹木年輪法分析森林衰退特征,從種群競爭(內因)方面研究中國半干旱區東段的森林衰退影響因素,為森林衰退機理研究提供依據,為半干旱區森林資源動態監測提供參考,對相關區域森林管理提供建議。
1 研究區概括
研究區位于中國干旱半干旱區東段內蒙古東北段地區,地理坐標為118°42′34″—123°42′14″ E,44°11′33″—50°56′40″ N(圖1),屬于大陸性季風氣候,冬季寒冷夏季溫暖。該區是亞洲內陸最大的生態過渡帶,在氣候上屬于干濕過渡帶,在植被分布上屬于林草過渡帶,是森林分布的干旱極限[29],種群競爭激烈,植被對氣候變化非常敏感[30]。在過去40年中該區域的年均降水為330 mm。夏季(6月—8月)平均氣溫和降水分別為16.5℃和232 mm,冬季(12月—次年2月)平均氣溫和降水分別為-24℃和8 mm[27—28]。研究區土壤類型主要為巖灰壤,地下水位大約為幾十米。植被主要是以興安落葉松(Larixgmelinii)天然林和人工林以及白樺(Betulaplatyphylla)天然林為主,伴有少量黑樺(Betuladahurica)、白樺(Betulaplatyphylla)、山楊(Populusdavidiana)、華北落葉松(Larixprincipis-rupprechtii)等。由于當地森林保護的各項政策,人為干擾措施主要表現為移除枯木而進行的擇伐,對興安落葉松人工林的種群競爭作用不顯著,因此在這里人為因素不作為考慮因素。
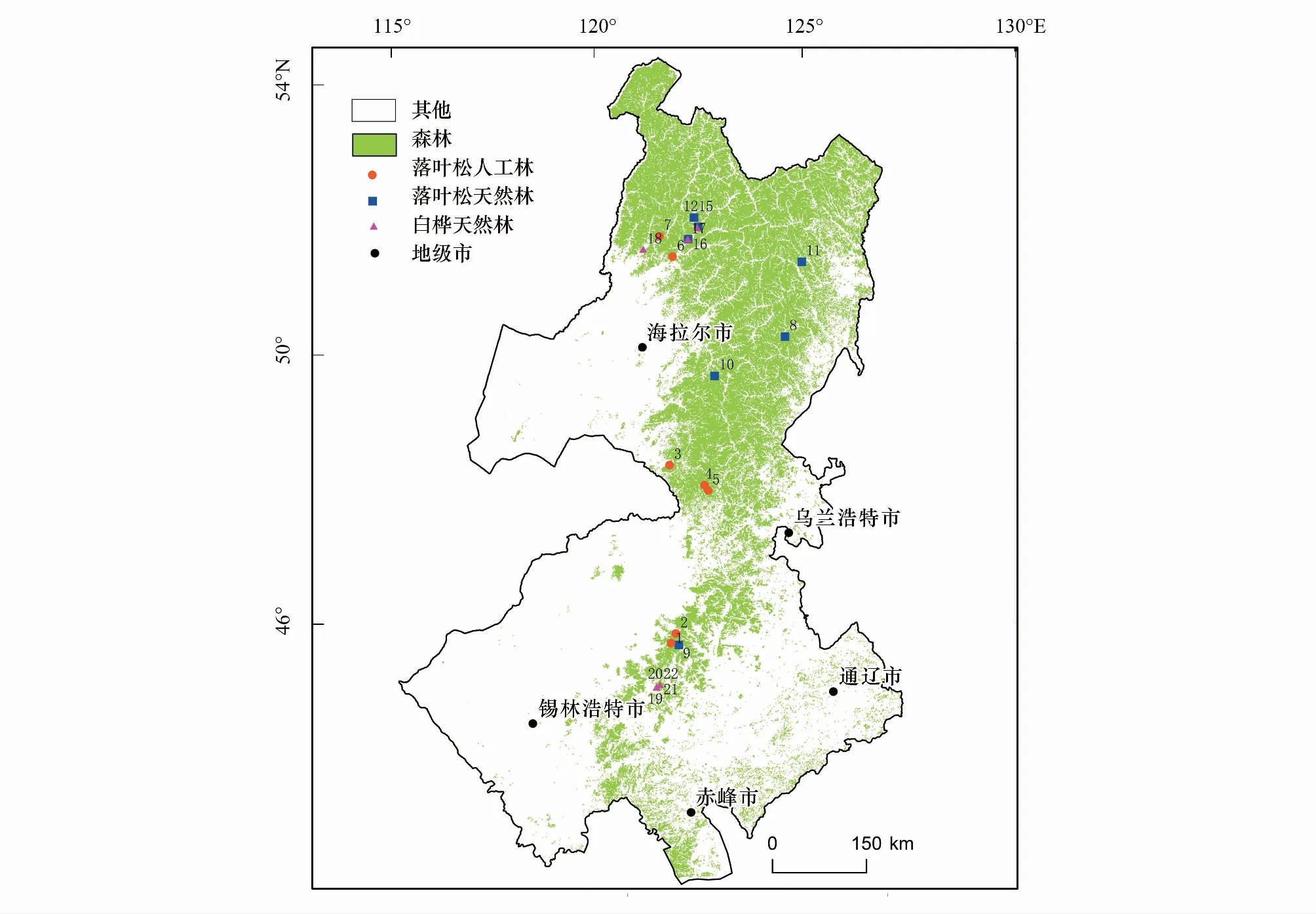
圖1 研究區采樣分布點Fig.1 Sampling distribution in the study area
2 研究方法
2.1 樣地調查
本項目于2016年7月—2017年8月進行兩次野外實地調查。樣地所在區域群落包括落葉松人工林、落葉松天然林、白樺天然林等(圖1)。選擇興安落葉松人工純林作為此次的研究對象。在研究區內自南往北的方向上設立盡可能遠離人為干擾且具有明顯森林衰退現象的典型樣地8個(圖1中紅色圓點標注),設置樣方面積30 m×30 m,在每個樣方內,記錄每棵樹的位置(X、Y坐標軸)用于進行種群競爭的分析,同時調查樹木的胸徑、基徑、樹高、冠幅、枝下高以及樣地的經緯度、海拔、坡度、坡向等因子。然后使用取樣直徑為5.15 mm、長度為400 mm的植物生長錐,在樹木位于平胸高度的部分順著自北向南和自西向東兩個不同方向采集兩個樹芯,每個樣地的采樣樹木至少30棵以上,以保證樣品量充足。長在坡上的樹木,取樣避免在向坡/背坡方向,在平行于坡的兩側取樣。
從生長錐取出樹芯保存于改良的吸管內(兩側開口,便于裝樣和通風干燥),同時用白色布基膠布封口,在吸管一頭同油性筆標注樣品號。本研究共采集318棵樹,一共636條樹芯,群落類型為興安落葉松人工純林,樣品信息見表1。
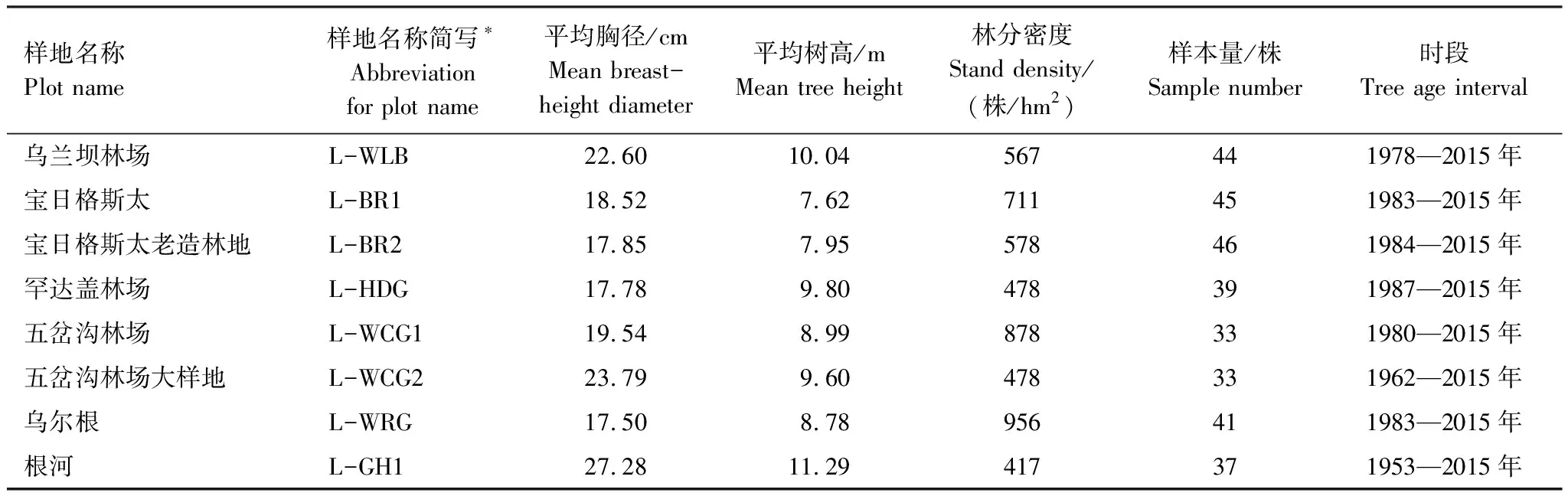
表1 年輪樣品信息
2.2 競爭指數選擇和競爭范圍的確定方法
本文根據桌面地理信息系統(Mapinfo)軟件的相鄰多邊形地理分析功能確定競爭范圍和競爭木。用胸徑倒數作為權重因子對泰森多邊形(Voronoi)圖進行加權,利用ArcGIS軟件中的加權泰森多邊形工具得到加權后的Voronoi圖。然后依據1974年Hegyi提出的單木競爭指數模型計算個體水平上和樣地水平上的競爭指數(具體內容見前期研究)[31]。由于國家對森林的政策保護措施,研究區近十年內未進行采伐作業,且林木自然更新周期較長。由于樣地內未發現明顯的樹樁、倒木及人工干擾現象,表明近十年內,樣地內很可能較少出現樹木個體的減少導致的種內和喬木種間競爭變化。且落葉松林胸徑,樹高等樹形因子在18年左右達到高峰后基本定型[32—33],而研究區用于分析樹輪的8個樣地都達到了20年以上的平均樹齡,因此使用采樣時觀測到的競爭指數可以近似反映近十年來幾個樣地的種群競爭狀況相對特征。
2.3 樹木年輪方法
樹木年輪的處理方法主要基于以下幾個步驟:
第一,依據國際通用規范,將樹輪樣品進行預處理。先將保存在吸管的樣品在實驗室陰涼通風處平鋪展開,陰干15 d左右。待樣品干燥后,用白色乳膠將年輪樣芯粘貼在定制的木槽之中,輔以棉線或者透明膠帶捆綁加固固定,以防樣品在其干燥時發生變形,繼續平鋪陰干大約兩周時間。等乳膠干透之后卸下棉線或膠帶進行樣品打磨。按順序使用280—600目的粒度從粗到細的砂紙打磨樣品,再用800目砂紙對樣品進行拋光,直至樣品表面平滑,年輪邊界清晰。
第二,采用顯微鏡目視定年的方法進行交叉定年,具體步驟如下:(1)選取四至六根生長狀況良好,年輪完整的樹芯先做處理;(2)根據采樣年份斷定靠近樹皮的那一年,繼而推測其他年份;(3)利用顯微鏡從樹皮端向內數樹木年輪,由于人工林的樹齡在40年左右,每數到年份為5年時在樣芯上做標記,整10年做兩個標記;(4)定年時將幾個樣本中多次同時出現的窄輪對應的年份做好標記,作為特征年份;(5)重復以上步驟,對所有樹輪樣本進行定年,定年時可參照之前選取的年輪樣本;(6)結合特征年輪和參照處理好的年輪樣品,對有偽輪、缺失年輪的樹芯進行定年。
第三,利用0.001 mm測量精度的LINTAB 5輪寬測量儀完成所有年輪樣品的寬度測量,將得到的每條樹芯的輪寬序列保存為raw文件。再使用COFECHA程序對raw文件的輪寬數值進行檢驗。其檢測原理是將每個輪寬序列與主序列進行相關分析,依據其相關系數檢驗是否存在測量錯誤或者定年錯誤的時段。再通過檢驗結果對存在錯誤的樹芯再次定年與測量,以保障年輪序列的正確性,同時剔除異常的樣本[34]。
第四,利用負指數或者線性函數消除大部分低頻變化,擬合出每個年輪的生長序列[35],再利用寬度的實際測量數值除以擬合值進行標準化,得到了無量綱序列,從而消除速生時期的樹木對年輪年際間變化的影響[36]。將每棵樹的年輪寬度分別用ARSTAN軟件處理得到樹木個體的生長年表[37],將樣地中所有的樹木年輪共同處理以建立樣地年表,數據形式為樹輪指數(TRI,Tree ring index)。
2.4 計算年輪指標
(1)個體生長衰退指標
當個體同時滿足以下3個標準時,可以定義該樹木個體發生了生長衰退[37]:第一,樹輪指數(TRI)持續4年以上低于1(由于此次調查的樣地樹齡偏小,衰退年數定為4年以上);第二,該時期年輪指數(TRI)的平均值低于0.75;第三,該時期存在兩年及兩年上的樹輪指數(TRI)最小值低于0.6。
(2)樣地生長衰退指標(胸高斷面積增量BAI計算)
為了保留TRI年表建立過程中可能被消除的重要變化趨勢,在此利用raw原始文件計算BAI,計算公式[38]如下:
式中,BAIt表示t年的胸高斷面積增量,單位為mm2;Rt表示t年的年增量結束時的樹木半徑,單位為mm;Rt-1是t年增量開始時的樹木半徑即前一年的樹木半徑,單位為mm。
又由于每年的年輪寬度(wt)相當于樹木半徑的年增量。因此,將wt代入,該公式可以表示為[39]:
BAIt=π (wt2+ 2wtRt-1)
求出每棵樹每年的胸高斷面積增量(BAI)后,將樣地中同一年所有樹木的BAI取平均值,得到樣地平均BAI序列,用比值法將每棵樹的BAI序列標準化即每棵樹每年的BAI實際數值除以每年樣地平均BAI,得到無量綱的數列也就是每棵樹的相對BAI序列(稱為rBAI)。
3 結果分析
3.1 個體水平上的森林生長(衰退)現狀分析
表2顯示了各樣地的樹間相關系數(即樣地內不同樹之間的年輪序列的相關性)、樹內相關系數(即樣地內同一棵樹的兩條樹芯的年輪序列的相關性)以及所有序列相關系數(即樣地內所有序列的相關性),綜合衡量樹木個體之間的生長趨勢的相似性[37]。從表2可以看出,樹間相關系數變化范圍為0.449—0.760,相對偏低;樹內相關系數相對較高,最低0.682,最高達到0.973;所有序列相關系數居中,變化幅度從0.460到0.762。所有樣地中,樣地L-WLB和樣地L-WCG2的所有序列相關系數都偏低,小于0.5,其他樣地所有序列相關系數都在0.5以上。各樣地的平均樹齡最小為樣地L-HDG的23年,最大為樣地L-GH1的48年。
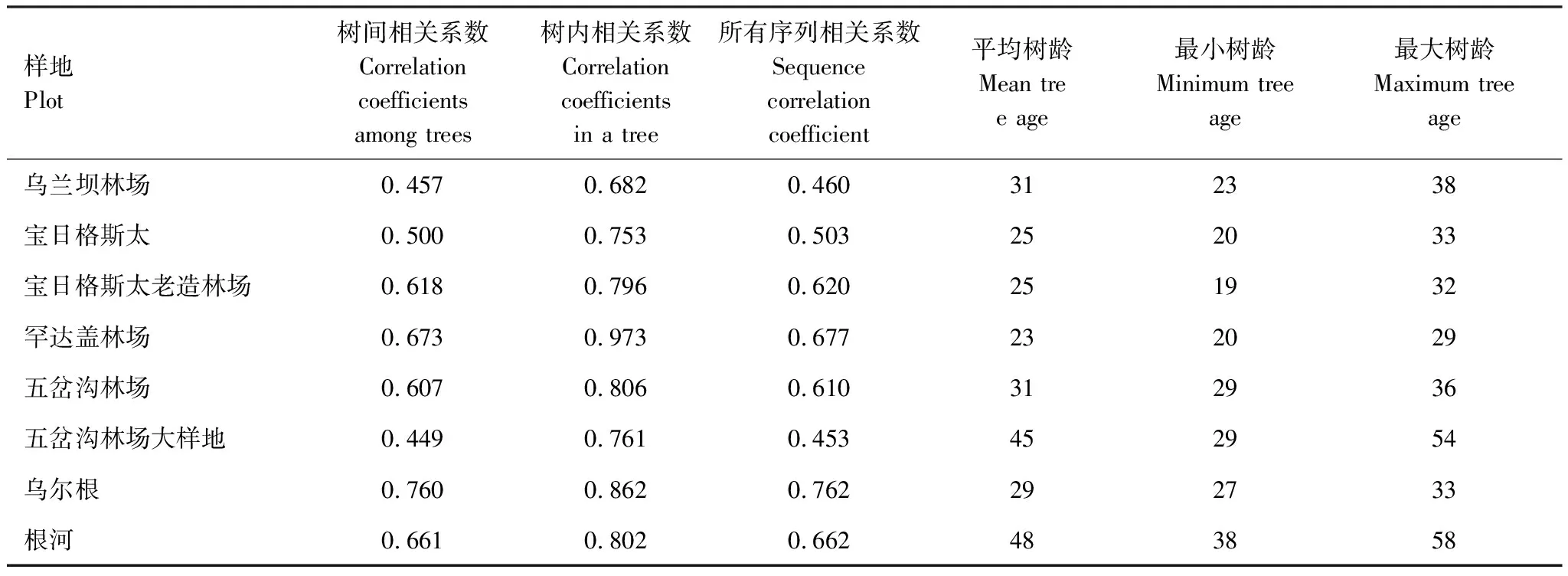
表2 各樣地的序列相關性系數及樹齡
通過3個衰退標準條件的篩選,得到發生生長衰退的樹木及對應的衰退時段,再將每一年發生衰退的樹木個數匯總,除以這一年的樹木總數,得到該年樹木衰退百分比,結果見圖2。對單個樹木個體生長軌跡的研究表明,在其樹輪年表中,出現生長下降期(> 4年)的樹木普遍存在(陰影面積)。設置樹木衰退百分比50%為閾值,即認為發生嚴重衰退事件。
圖2顯示各樣地的衰退樹木所占百分比及樣本數量信息,得出樣地L-WLB的嚴重衰退時期發生在2001—2005年,影響25棵樹木即占樣地樹木的57%,不存在衰退現象的樹木達到23%;樣地L-BR1的衰退事件發生在1983—1985年間,由于該時段實測到的樹木個數只有1—5棵,不能真實反映出衰退樹木與當年所有樹木的比例信息,同樣情況發生也在樣地L-BR2,它的嚴重衰退期在1985—1986年期間,樹木個體數量只有3、4棵,存在衰退現象的樹木個數為2,兩個樣地不存在衰退現象的樹木比例分別是36%和48%;樣地L-HDG發生嚴重衰退的時間介于2000—2005年間,出現衰退現象的樹木數量達到23—36棵,占比同年樹木數量的59%—92%,不存在衰退現象的樹木比例為3%;樣地L-WCG2的嚴重衰退時期為2000—2005年,影響樹木達到19—23棵,占比同時期樹木數量的58%—70%,不存在衰退現象的樹木比例為12%;樣地L-WRG于2000—2007年期間存在嚴重衰退現象,受影響樹木個體多達35棵,所占同時期樹木總數的51%—85%,不存在衰退現象的樹木比例為5%;而只有樣地L-WCG1與樣地L-GH1在樹木個體生長TRI年表上無嚴重衰退時段,但從整體上看,兩個樣地所存在的衰退事件的陰影面積都占據一定比例,不存在衰退現象的樹木占比分別為30%和27%。
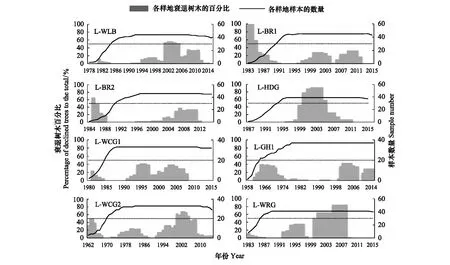
圖2 各樣地樣本數量及衰退樹木所占百分比Fig.2 Sample number and the percentage of degraded trees in each plotL-WLB:烏蘭壩林場;L-BR1:寶日格斯太;L-BR2:寶日格斯太老造林地;L-HDG:罕達蓋林場;L-WCG1:五岔溝林場;L-GH1:根河;L-WCG2:五岔溝林場大樣地;L-WRG:烏爾根
將所有樣地的衰退現象綜合分析,發現樣地L-WLB、L-HDG、L-WCG2和L-WRG發生嚴重衰退的時段重合期在2000—2005年間。
3.2 樣地水平上的森林生長(衰退)現狀分析
將樣地中所有的樹木年輪共同用ARSTAN軟件去趨勢和標準化處理得到樣地TRI年表,具體信息如圖3。圖3中顯示的各樣地的樹輪指數序列中,可以看出,位于五岔溝的樣地L-WCG1和L-WCG2,以及位于根河的樣地L-GH1的生長序列即TRI年表長度相對其他樣地都比較長,其中樣地L-GH1的TRI年表最長,一共為58年,時段為1953—2015年;樣地L-HDG的TRI年表最短,共29年,時段為1987—2015年。從波動幅度來看,樣地L-WRG的年輪指數(TRI)上下波動范圍最大,差值達到2.224,樣地L-BR1的TRI波動幅度最小,差值為1.405。
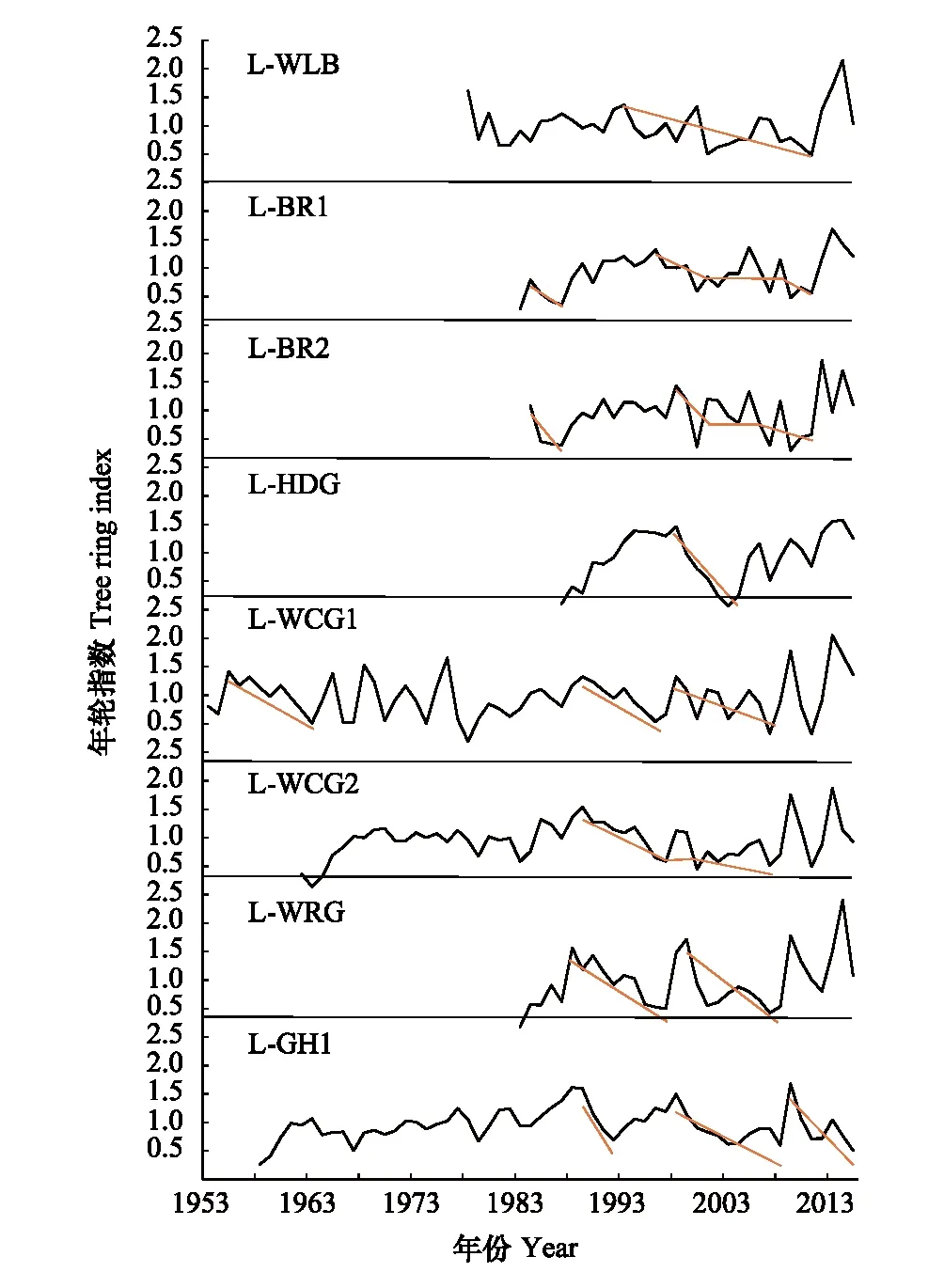
圖3 各樣地的年輪指數序列 (紅色線條代表年輪指數下降即森林衰退的趨勢)Fig.3 Annual ring index series of various plots (red lines represent the trends of forest decline)
結合趨勢線和年表序列來看,樣地L-WLB的TRI年表在2000年前的波動幅度較小,1993—2011年間TRI指數整體呈現下降趨勢,期間在2000年出現生長釋放。樣地L-BR1的TRI指數在1984—1997年間,1996—2011年間出現下降趨勢,期間在2005年、2008年TRI出現回升。樣地L-BR2的TRI指數分別在1984—1987年、1998—2011年總體呈現下降的趨勢,其中在2000年TRI跌至12.86。樣地L-HDG中,TRI數值在1998—2004年內持續下降,在2003年出現TRI年表的最低點即0.17。樣地L-WCG1的TRI年表總體上波動幅度相對比較劇烈,TRI指數在1955—1963年、1989—1995年持續下降,1998—2007年在幾次波動中呈現下降趨勢。樣地L-WCG2的TRI在1989—2008年間呈現下降趨勢,期間在1998年出現生長釋放的TRI回升現象。樣地L-WRG的TRI指數在1988—1997年間的幾次小波動中持續下降,之后快速上升,并在1999年開始下降直到2008年。樣地L-GH1的TRI指數在1998—2008年持續下降,而后徒增至2009年的1.63,之后至2015年都持續下降。
可以看出,樣地TRI年表與樹木個體TRI年表所指示的衰退時段基本一致,即圖2的陰影面積對應的年份和圖3紅線部分的對應年份有很高的重疊性。由此可以看出,樣地L-WLB的衰退年份是1993—2011年;樣地L-BR1的衰退年份是1984—1987年、1997—2011年;樣地L-BR2的衰退年份是1985—1987年、2000—2011年;樣地L-HDG的衰退年份是1999—2004年;樣地L-WCG1的衰退年份是1992—1995年、1998—2007年;樣地L-WCG2的衰退年份是1992—2008年;樣地L-WRG的衰退年份是1993—1997年、2000—2008年;樣地L-GH1的衰退年份是2000—2008年、2010—2015年。
將樣地間進行對比,發現存在不同樣地間生長衰退時段(即年表中TRI指數持續下降時期)重合的現象:樣地L-WCG1、L-WCG2和L-WRG存在1989—1997年的衰退重合時期,樣地L-HDG、L-WCG1、L-WCG2、L-WRG和L-GH1的衰退時段都在1998—2003年前后出現。
3.3 個體水平上競爭指數與樹木生長(衰退)的關系分析
由于該年采集的競爭指數可以相對反映近十年樣地樹木的競爭近似狀況,結合近幾年的生長趨勢進行分析樹木個體的競爭狀況與個體衰退的關系,本研究分別選取近2年(生長年份為2014—2015年)、近5年(生長年份為2011—2015年)以及近10年(生長年份為2006—2015年)的樹木個體相對胸高斷面積增量rBAI2、rBAI5、rBAI10的平均值,與對應的樹木個體的競爭指數做相關性分析。結果如表3。
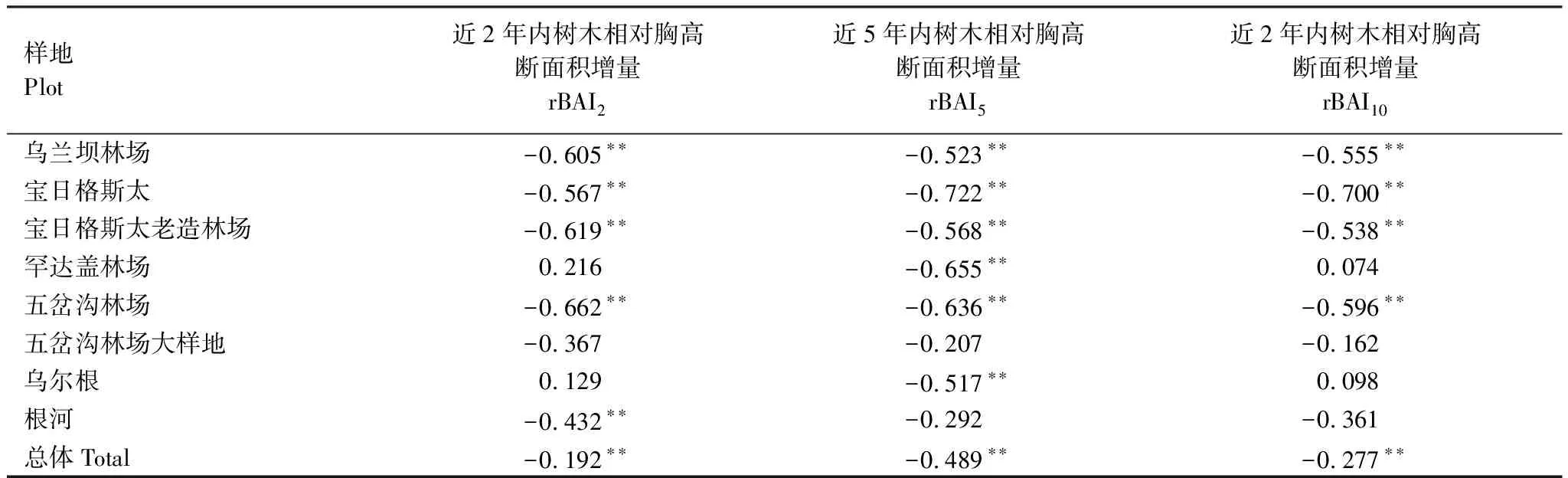
表3 各樣地樹木個體競爭指數與rBAI的相關性分析
從表3可以得出,總體樣本量上,個體水平的競爭指數與rBAI2、rBAI5、rBAI10的3個指標都呈現出極顯著的負相關關系(P<0.01)。其中樣地L-WLB、L-BR1、L-BR2、L-HDG、L-WCG1和L-WRG的6個樣地的樹木個體競爭指數與rBAI5呈現極顯著的負相關關系,且相關系數絕對值都在0.5以上。在rBAI2與個體競爭指數相關性分析中,樣地L-WLB、L-BR1、L-BR2、L-WCG1和L-WRG的5個樣地中兩者呈現顯著性相關。在rBAI10與個體競爭指數相關性分析中,共有4個樣地(樣地L-WLB、L-BR1、L-BR2和L-WCG1)的相關性顯著,且絕對值也都在0.5以上。其他樣地的個體競爭指數與相關指標之間的關系不顯著。
在樣地的橫向比較中,樣地L-WCG2的rBAI2、rBAI5、rBAI10的3個指標與其個體競爭指數的相關性都不顯著,可能的原因是該樣地的樣本量最少,只有19棵,導致其相關關系不顯著;樣地L-HDG與L-WRG的rBAI2、rBAI10都與個體競爭指數呈現不顯著正相關,而rBAI5卻與個體競爭指數呈現顯著性負相關。所有樣地的個體競爭指數與3個指標之間的顯著關系均為負相關,因此,個體競爭指數越高,近十年內的相對胸高斷面積增量越低,這也說明了種群競爭易造成樹木生長量的下降。
在3個指數之間,rBAI5與各樣地的個體競爭指數相關性程度較高,因此將rBAI5與個體競爭指數做一個擬合分析得到圖4。
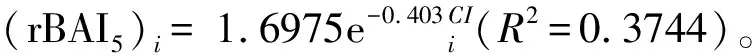
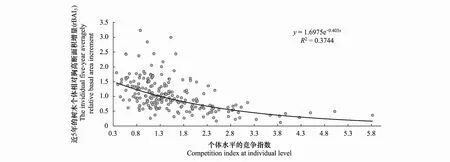
圖4 樹木個體的rBAI5與競爭指數的變化關系Fig.4 The relationship between individual tree rBAI5 and competition index
3.4 樣地水平上競爭指數和樣地生長(衰退)的關系分析
分別選取近2年(生長年份為2014—2015年)、近5年(生長年份為2011—2015年)以及近10年(生長年份為2006—2015年)的樣地胸高斷面積增量BAI2、BAI5、BAI10的平均值代表樣地生長的近期狀況,與對應的樣地競爭指數進行比較,結果如圖5。
可以看出,各樣地的3個胸高斷面積增量指標BAI2、BAI5、BAI10相差并不大,變化趨勢也相對一致,多數樣地的大小規律是:BAI2>BAI5>BAI10。在樣地L-WCG1中,BAI5最大;在樣地L-GH1中BAI10最大,BAI5居中,BAI2最小。
結合樣地競爭指數進行對比分析,發現樣地競爭指數最高(124.91)的樣地L-WRG的胸高斷面積增量最小,樣地L-GH1、L-WLB、L-WCG2的樣地競爭指數在28.65—36之間,胸高斷面積增量也在6.5—9 mm2左右,相差不大;樣地L-HDG、L-BR2的樣地競爭指數在44—48的區間,兩個樣地的三種胸高斷面積增量指標兩兩之間相差不大。因此,說明各樣地的樣地競爭指數與胸高斷面積增量的相對關系比較穩定。
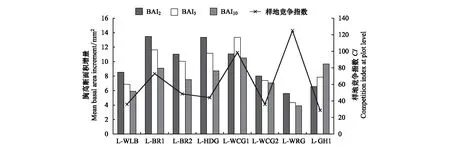
圖5 樣地競爭指數與樣地胸高斷面積增量比較Fig.5 Plot competition index and basal area increment in each plotBAI2:近兩年內樣地平均胸高斷面積增量Mean basal area increment in two years;BAI5:近五年內樣地平均胸高斷面積增量Mean basal area increment in five years;BAI10:近十年內樣地平均胸高斷面積增量Mean basal area increment in ten years;CI:樣地競爭指數Competition index at plot level
4 討論
4.1 個體水平的樹木衰退狀況分析
在個體水平上,根據衰退的定義標準,利用年輪指數(TRI)篩選出生長衰退的樹木并計算其所占百分比,發現生長衰退的現象在各樣地的樹木個體間普遍存在。從衰退百分比所占的陰影面積來看,衰退現象最為嚴重的是樣地L-WRG2,其次是樣地L-WRG,兩者衰退嚴重時期存在衰退現象的樹木所占百分比高達70%以上。通過樣地間衰退時期的對比,發現各樣地超過閾值50%的嚴重衰退時期年份基本在2001—2005年間。這是由于該時期這一地區發生了大規模的干旱事件,尤其是2000年和2001年干旱程度最嚴重[40—41],導致不同地點的樹木生長都受到了抑制。
4.2 樣地水平的森林衰退狀況分析
在樣地水平上,通過建立八個樣地樹輪指數序列組成的TRI年表,并擬合各樣地的生長趨勢,顯示各樣地都存在TRI指數持續減小的生長衰退時段,結合樹木個體TRI年表,發現兩者指示的衰退時段有共同性,結果較為一致。通過樣地間對比發現,不同樣地間的生長衰退時段有重合的現象,比如大部分樣地的衰退時段都在1998—2003年前后出現。從地理位置上可以發現樣地L-HDG、L-WCG1和L-WCG2的地理坐標相近,樣地L-WRG和L-GH1的位置也比較接近,這與前人對該區域干旱分析得出的嚴重干旱發生的年份相一致[41],說明干旱的發生直接導致了森林的衰退。從整體年表上來看,樣地L-GH1的生長趨勢出現衰退現象,其他樣地的TRI整體上有所增長。
4.3 種群競爭與樹木生長的關系分析
樹木個體的三種平均相對胸高斷面積增量rBAI2、rBAI5、rBAI10與個體水平競爭指數CIi都呈現顯著性負相關,各樣地中rBAI5與個體競爭指數相關的顯著性最高。將兩者通過趨勢擬合分析,發現兩者的關系可用指數函數方程表達,即個體的五年平均相對胸高斷面積增量隨著個體水平競爭指數的上升而下降。這說明了競爭指數對于樹木生長存在顯著的影響。樹木受到相鄰木的競爭壓力越大,其生長越受到抑制,可表示為胸徑的徑向生長量減少,特別是當資源有限時,個體間的競爭會加劇,造成有些樹木因獲取資源受到限制而逐步地形成生長緩慢甚至是生長停滯期,增大其死亡的風險性[42]。另一方面,樹木胸徑越大,所占據的空間越大,其競爭能力就越強,徑向生長的增量就相對更高。有研究表明,樹木間的競爭關系和競爭強度很可能比外部環境因素更能致使樹木徑向生長的增量減少,生長趨勢降低[42]。樣地競爭指數與近年樣地胸高斷面積均值的關系并沒有明顯的升降變化趨勢,但同一水平的樣地競爭指數所對應的近年樣地胸高斷面積均值的差異性并不大,說明兩者之間可能存在內在聯系。然而由于樣地水平上的生長狀況和競爭指數變化是由多種因素共同作用的結果,不同因素對兩個指標的影響程度也不同。
5 結論
TRI年表顯示出現生長下降期(> 4年)的樹木普遍存在。根據個體TRI年表所指示的衰退情況發現,樣地L-WLB、L-HDG、L-WCG2和L-WRG發生嚴重衰退的時段重合期在2000—2005年間。只有樣地L-WCG1與樣地L-GH1在樹木個體生長TRI年表上無嚴重衰退時段。
樣地TRI年表與樹木個體TRI年表所指示的衰退時段基本一致,即存在不同樣地間生長衰退時段(即年表中TRI指數持續下降時期)重合的現象:樣地L-WCG1、L-WCG2和L-WRG存在1989—1997年的衰退重合時期,樣地L-HDG、L-WCG1、L-WCG2、L-WRG和L-GH1的衰退時段都在1998—2003年前后出現。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
