鳶尾糖基轉移酶ItUGT797基因的克隆與表達分析*
2022-09-29 15:19:18黃梓璐陶無恙陳嘉杰段禮新季愛加
世界科學技術-中醫藥現代化 2022年5期
關鍵詞:植物
姚 薇,李 靜,黃梓璐,陶無恙,陳嘉杰,段禮新,季愛加
(廣州中醫藥大學中藥學院國際中醫藥轉化醫學研究所 廣州 510000)
鳶尾(Iris tectorumMaxim.)別名紫蝴蝶、蛤蟆七,性寒味辛、苦,是鳶尾屬植物,遍布于各個國家,植株可入藥也可觀賞[1]。在中國,鳶尾較多分布于陜西、甘肅、青海等西北地區以及四川、貴州、云南等西南地區[2]。鳶尾的干燥根莖可加工為中藥川射干,可活血祛瘀以及治療咽喉腫痛等,因此被稱為“治療喉痹咽痛之要藥”。川射干主要藥效成分為異黃酮及其糖苷類化合物[3],此類化合物在解除疼痛、治療炎癥、過敏以及癌癥等方面效果良好[4-6],其生物合成更是備受關注。
糖基化反應是糖苷類化合物生成途徑上涉及到的重要修飾步驟,該反應依賴于糖基轉移酶[7]。其過程主要涉及的供體分子是活化后的糖,它在轉移酶的催化作用下轉移到特定的受體上,從而形成不同的糖苷類化合物,同時苷元的穩定性、親水性以及生物活性都能得到改善[8-10]。據報道糖基轉移酶依據序列相似度和催化特異性可分為99 個家族,其中尿苷二磷酸-糖基轉移酶(UGT)家族與植物有效活性成分的合成聯系最為密切。UGT 家族作為參與植物發育以及代謝調控的重要糖基轉移酶家族,廣泛分布于各個物種中,模式作物擬南芥中(120)[11]、水稻(180)[12]以及大豆(182)[13]中的UGT 家族成員已經被鑒定出來。另外,藥用植物柴胡、地黃中均有10 個UGT 基因片段被發現[14],有待克隆和功能驗證。三七中被鑒別出來的PnUGT82 可以催化黃酮類產物的生成[15],來自紅花的CtGT3 可催化7 種化合物進行糖基化[16];黃芩中也有3種新穎的O-糖基轉移酶被鑒定出來,既可催化形成黃芩的主要糖苷類成分,也能夠催化外源的黃酮底物發生O-糖基化反應[17];金花茶中的CnUFGT14 則有促進多酚合成的作用[18]。底物寬泛性是UGT 家族成員的特性之一,除此外,該家族的成員在C-末端均具有一個PSPG(Plant Secondary Product Glycosyltrans-ferase box)盒子,是包含44 個氨基酸的高度保守區域,而N端則變化多樣。UGT 家族成員的結構特征是其行使催化功能、作用底物不一的重要基礎[19]。
川射干中的主要活性物質鳶尾苷(tectoridin)是通過異黃酮代謝支路合成。在查爾酮異構酶(chalcone isomerase, CHI)的催化作用下,異甘草素(isoliquiritigenin)合成甘草素(liquiritigenin)。隨后在異黃酮合酶(isoflavone synthase,IFS)及2-羥基異黃烷酮脫水酶(2-hydroxyisoflavanone synthase, HID)的催化作用下合成異黃酮骨架,合成鳶尾苷之前還需要在其C-6 位加上甲氧基最后再糖基化。與其他植物例如大豆中異黃酮具有較為顯著的雌激素樣作用不同,射干類藥材異黃酮藥理活性更偏向于明顯的抗菌消炎等作用[20]。通過兩者結構比較發現,大豆異黃酮C-5 位多數沒有羥基而射干類藥材中的異黃酮則具有C-5 位羥基的結構,從鳶尾中異黃酮結構就可以很明顯的發現這一差別,另外鳶尾中的異黃酮多數含有甲氧基。可以推測由于兩者結構上的差別造成其在活性上的不同。而糖基化修飾作為關鍵環節參與了異黃酮結構的形成,可預測該過程對于異黃酮的藥理活性也有一定的影響。因而解析川射干中糖基轉移酶的修飾功能以及機制對其有效活性成分異黃酮類物質的合成及藥理活性具有重大意義。
目前植物中已經被表征功能的UGT 成員很多,涉及到黃酮、萜類以及多種植物激素的生物合成,能夠間接或者直接調控植物的生長發育以及品質。而鳶尾作為藥用并觀賞兩用的植物擁有較高的市場價值,其中的UGT 家族成員鮮見報道,因而本研究著重于挖掘鳶尾中的可能參與有效成分鳶尾苷生物合成的UGT 家族成員。利用已知功能的同源基因從鳶尾轉錄組數據blast 篩選并克隆出一條糖基轉移酶基因ItUGT797[21],并對其編碼蛋白進行了特征解析,通過同源重組的方法將其克隆到pET-32a 載體上并進行體外原核表達,成功表達出可溶性蛋白,實時熒光定量PCR 實驗顯示該基因在不同部位均有不同程度表達。這些結果為進一步探索ItUGT797的具體功能及其在鳶尾苷的生物合成中的作用奠定了基礎。
1 方法與試劑
1.1 鳶尾總RNA提取以及cDNA的獲得
實驗材料植物鳶尾(Iris tectorumMaxim.)于2019年6 月采自北京中國科學院植物研究所,后在廣州中醫藥大學栽種。選取生長年限2年,生長健壯,處于盛花期的鳶尾植物,洗凈,取須根、根狀莖、葉片和花等新鮮組織后立即置于液氮中速凍。參照北京華越洋生物科技有限公司RNA提取試劑盒說明書(Lot:2016#08),提取各組織部位RNA。RNA 提取液各取1μL 進行瓊脂糖凝膠電泳檢測看是否具備完整條帶,其濃度則用島津BioSpec-nano 核酸定量儀測定。確認合格的mRNA 用于合成川射干的cDNA 鏈,具體操作按照TakaRa 公 司 的 PrimeScriptTMII 1st Strand cDNA Synthesis Kit反轉錄試劑盒中的說明進行。
1.2 川射干ItUGT797基因的克隆及表達載體構建
以已知功能的類黃酮7-O-UGT 作為Query 序列,以川射干轉錄組的pep 文件為Library 進行本地BLASTp,綜合基因在根狀莖、葉子以及花中的FPKM值與系統發育進化樹結果篩選到一條潛在的糖基轉移酶基因,命名為ItUGT797。根據其CDS 序列信息,使用Vector NTI設計引物(表1),以鳶尾各部位的混合cDNA 為模板,在冰上按照表2 順序加入體系,輕彈混勻后按照表3程序進行PCR擴增反應。獲得的擴增產物參照生工生物工程(上海)股份有限公司的SanPrep柱式PCR 產物純化試劑盒說明書進行回收。原核表達載體pET32a 采用雙酶切線性化(酶切位點為EcoRI和XhoI)的手段獲得具有粘性末端的pET32a 骨架,通過南京諾唯贊生物科技有限公司的同源重組酶將線性化的載體與膠回收產物連接。同源重組的體系按照ClonExpress?ⅡOne Step Cloning Kit 的說明書進行配置。同源基因片段和線性化載體后將其轉化至大腸桿菌DH5α,涂布于含100 mg·L-1濃度的氨芐青霉素鈉抗性的平板上,置于37℃培養箱過夜培養。挑取大小適中的單克隆進行PCR 鑒定,選取擁有單一且明亮的條帶的陽性菌液送測序,測序比對正確后提取質粒,獲得pET-32a(+)-ItUGT797重組載體。

表1 引物序列
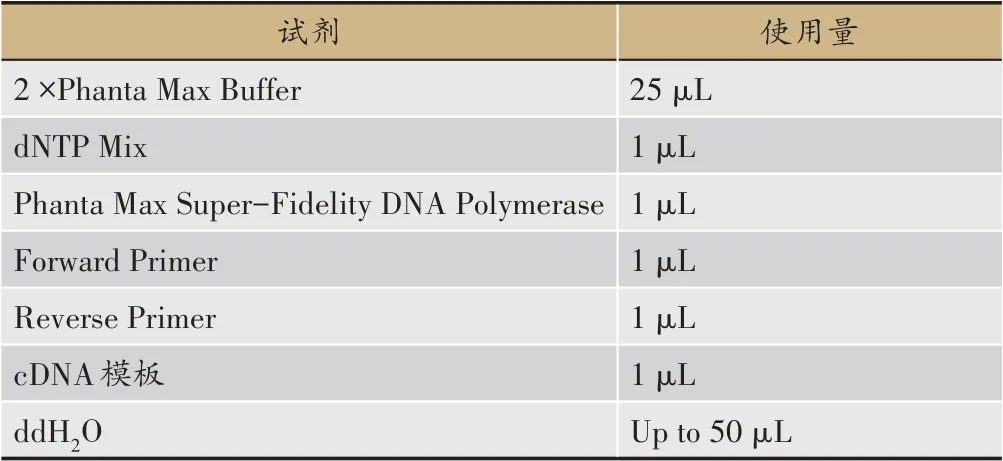
表2 高保真酶PCR 擴增反應體系
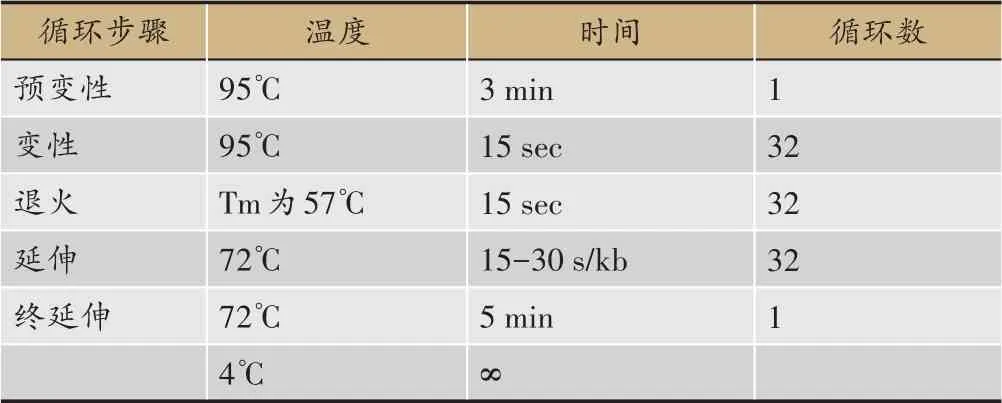
表3 高保真酶PCR 擴增反應程序
1.3 生物信息學分析
將測序得到的ItUGT797序列用各類在線工具解析(表4)。首先利用NCBI 在線工具ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder)搜尋擴增候選糖基轉移酶基因ItUGT797序列的開放閱讀框;在線工具ProtParam(https://web.expasy.org/protparam/)、CELLO v2.5(http://cello.life.nctu.edu.tw/)和SignalP-5.0(http://www.cbs.dtu.dk/services/SignalP/)被分別用來預測該蛋白的理化性質、亞細胞定位和蛋白信號肽;蛋白跨膜區和蛋白的二、三級結構則分別使用在線工具TMHMM(http://www. cbs. dtu. dk/services/TMHMM/)、SOPMA (https://npsa-prabi. ibcp. fr/cgi-bin/secpred_sopma.pl)和SWISS-MODEL(https://swissmodel.expasy.org/interactive)來預測。DNAMAN 軟件被用來進行序列比對,NCBI-cdd 和pfam 網站被用來預測蛋白的結構域范圍,最后使用MEGA 7.0 軟件中的Neighbor-Joining 法預測基因與其他已被驗證功能的糖基轉移酶基因的進化關系。

表4 功能表征UGT來源
1.4 川射干ItUGT797 基因Real-time PCR 分析
采用QuantStudio5 實時熒光定量PCR 儀對ItUGT797在各部位的表達情況進行檢測,以eEF 基因為內參[21]。鳶尾各組織部位的mRNA 則參照TakaRa公司的反轉錄試劑盒(RR047A)中的操作說明合成cDNA 鏈用作模板。按照以下體積配置10 μL 反應原液:2×SYBR Green Pro Taq HS Premix(艾科瑞公司),5.0 μL;正反向引物各0.2 μL;熒光染料0.2 μL;稀釋20 倍的cDNA 模板1 μL;超純水3.4 μL。反應條件:95℃,30 s;40個循環(95℃,5 s;64℃,34 s)。每個樣品三次重復,使用2-△△Ct法檢測ItUGT797基因在根狀莖、葉片和花等部位中的表達水平。
1.5 原核表達
將重組質粒pET-32a(+)-ItUGT797轉入宿主菌BL21(DE3)感受態,PCR 鑒定選取陽性克隆菌,接種于1 mL 帶有氨芐抗性的LB 液體培養基中,置于37℃搖床,200 rpm 轉速培養,至菌液渾濁,以1:100 的比例接種于500 mL 帶有氨芐抗性的LB液體培養基中。轉速與溫度同前,菌液培養至OD 值達到0.5-0.6 的程度即可。18℃,靜置30 min,分別加入終濃度0.1 mM、0.3 mM、0.5 mM 的IPTG 進行三個平行實驗以摸索最佳的誘導劑濃度。在18℃,180 rpm 條件下誘導培養18 h。后續則在4℃下收集菌體,棄上清。重懸菌體時加入50 mM Tris-HCl,蛋白破碎前加入1 mM PMSF。超聲破碎蛋白條件為:間隙時間5 s,超聲時間2 s,全程時間18 min,超聲功率30%,以上重懸至超聲破碎過程均在冰水浴中進行。4℃,12000 rpm,離心30 min,分離上清及沉淀。最終的蛋白表達結果使用SDS-PAGE檢測。
2 結果分析
2.1 鳶尾各組織部位RNA的獲取
經過1%的瓊脂糖凝膠電泳檢測,從鳶尾當中提取的各部位的28S 和18S 條帶均完整無彌散(圖1),凝膠條帶亮度越高表明提取RNA 的濃度越高,其中花亮度最高,表明其濃度最大,根狀莖的濃度次之,葉片濃度最低,提取的各部位RNA 質量均較好,均可用于接下來反轉錄實驗。
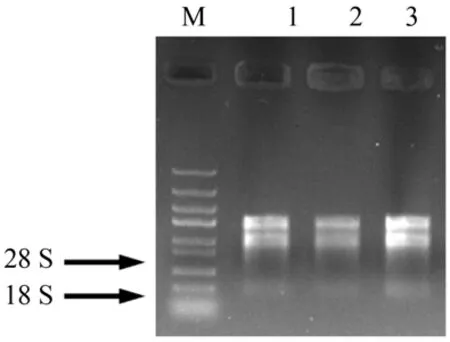
圖1 RNA提取凝膠電泳圖
2.2 鳶尾ItUGT797基因的克隆以及表達載體構建
以轉錄組中ItUGT797的CDS 序列為基準,設計帶有同源臂的特異性引物,擴增模板為鳶尾的各部位的混合cDNA。電泳結果顯示PCR 擴增出的條帶處于1500 bp左右(圖2),符合ItUGT797基因實際大小。參照生工上柱膠回收之后,通過同源重組的方法將帶有同源臂的ItUGT797基因連接到線性化的pET-32a(+)表達載體上,轉化后挑取單克隆進行驗證,后將陽性菌液測序,測序結果顯示與轉錄組序列一致。而經在線工具預測ItUGT797開放閱讀框長度為1455 bp,編碼484個氨基酸。
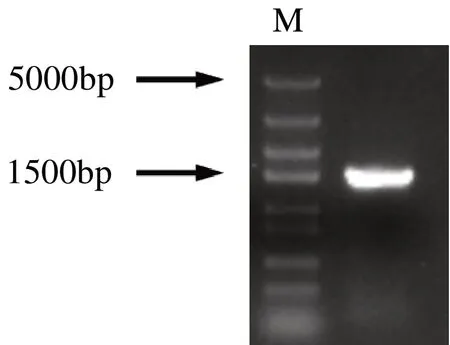
圖2 鳶尾ItUGT797基因的克隆
2.3 ItUGT797的生物信息學以及序列分析分析
2.3.1 ItUGT797的理化性質、跨膜區及信號肽預測
ItUGT797 的分子量大小(Molecular,MW)、理論等電點(Theoretical pI)和不穩定系數(Instability index,Ⅱ)等均是通過ProtParam 在線網站預測,結果分別為53.80 kDa,4.94以及52.52。不穩定系數評價結果顯示ItUGT797 屬于穩定蛋白。ItUGT797 經預測可能定位于細胞質。從在線工具TMHMM 預測的結果來看,ItUGT797 不含有跨膜螺旋區,屬于膜外蛋白(圖3)。SignalP-5.0 網站顯示ItUGT797 沒有信號肽,屬于非分泌蛋白(圖4)。推測ItUGT797 可能在細胞質中發揮功能。
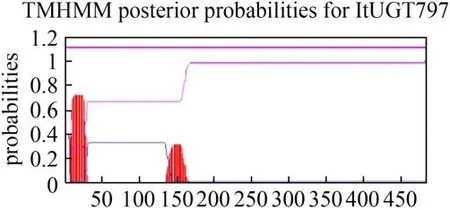
圖3 鳶尾ItUGT797的跨膜區預測
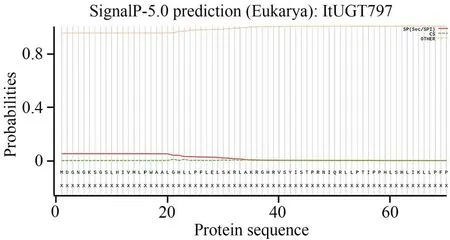
圖4 鳶尾ItUGT797的信號肽預測
2.3.2 ItUGT797蛋白的二級與三級結構預測
在線工具SOPMA 預測ItUGT797 的二級結構如下:無規則卷曲組成占比最大,為44.42%,較為均勻地分布于各處;其次是占比35.95%的α 螺旋,延伸鏈占比為13.43%,轉角占比最少為6.20%。結果表明ItUGT797 主要由無規則卷曲和α 螺旋組成(圖5)。SWISS-MODEL預測ItUGT797基因編碼蛋白的三維結構表明,該蛋白與氫醌葡萄糖基轉移酶具有26.92%的序列相似性,同源建模以該酶蛋白(2vg8.1)A 鏈為模板,建模范圍為9-470 個氨基酸,GMQE 值(全球性模型估測值)為0.59,三維建模質量良好(圖6)。

圖5 鳶尾ItUGT797基因的蛋白二級結構預測

圖6 鳶尾ItUGT797基因的蛋白三級結構預測
2.4 ItUGT797的系統發育與結構域分析
為了預測篩選出來的ItUGT797的功能,將其與已被表征過功能的擬南芥、葡萄、大豆、蒺藜苜蓿、葛根等19 種植物UGTs 進行建樹。從pfam 網站預測的結果來看,這些UGTs 包括ItUGT797均具有UDPGT 這一關鍵結構域。根據文獻報道,黃酮類化合物的UGTs具有明顯的底物偏好性,包括糖受體(黃酮醇、花色素、黃烷酮、黃酮、異黃酮和黃烷醇等苷元)和糖供體(UDP-葡萄糖、UDP-半乳糖、UDP-鼠李糖、UDP-阿拉伯糖和UDP-葡萄糖醛酸)的多樣性和特異性[8,22]。根據催化位點的不同,主要可分為O-糖基轉移酶(O-UGT)和C-糖基轉移酶(C-UGT)兩大類。黃酮類化合物的O-糖基轉移酶可進一步分為3-OUGT、5-O-UGT、7-O-UGT 和4-O-UGT。系統發育進化樹結果表明ItUGT797聚類在Cluster 1,與UGT89C1、Nt7GlcT 等具有7-O-糖基化的活性的UGTs 聚在一支上(圖7)。Cluster 2 聚類的則是有異黃酮5-O 位的糖基化活性的UGTs,Cluster 3 均是3-O-UGTs。從建樹結果可推測ItUGT797 可能是一種類黃酮7-O-UGT 糖基轉移酶。

圖7 鳶尾ItUGT797與其他物種來源UGTs的系統進化分析
2.5 ItUGT797基因的原核表達
將構建好的表達載體pET-32a(+)-ItUGT797轉化至宿主菌BL21(DE3)中,使用菌液PCR 驗證是否轉化成功。結果表明,目的條帶大小與預期一致(圖8),取陽性菌液進行IPTG誘導的原核表達。

圖8 重組子質粒轉入宿主菌BL21(DE3)菌液PCR電泳圖
經過改造的pET-32a(+)載體攜帶20.4 kDa大小的融合標簽,旨在促進蛋白的可溶性表達。IPTG 能夠誘導重組質粒在大腸桿菌體系中的異源表達,最終目的蛋白以融合蛋白的形式經由SDS-PAGE 表現。相較于空載對照組,pET-32a(+)-ItUGT797 蛋白大小理論值為75.82 kDa,實際條帶符合理論值(圖9)。改變其IPTG 蛋白誘導劑終濃度這一變量,摸索目的蛋白表達的較優條件。在誘導溫度為16℃條件下,分別加入終濃度為0.1 mM、0.3 mM、0.5 mM 的IPTG 誘導劑誘導表達12 h,最終發現誘導劑IPTG 終濃度為0.1 mM 時,上清液可溶性目的蛋白含量最高,表達情況較優(圖9)。

圖9 改變誘導劑濃度后鳶尾pET-32a-ItUGT797蛋白原核表達SDS-PAGE膠圖
2.6 ItUGT797基因在不同組織的轉錄水平分析
實時熒光定量檢測ItUGT797基因在鳶尾根狀莖、葉片和花中的表達結果以根莖為準進行歸一化(圖10)。花中ItUGT797表達最高,其次是葉片,根狀莖中水平最低,基因在植物不同部位的表達情況與其功能存在一定的聯系[23-25],本研究中ItUGT797基因在花中的高表達可能暗示其更多地在鳶尾花中發揮功能。
3 討論
類黃酮是廣泛存在于蔬菜水果以及觀賞性植物中的多酚類化合物,參與植物生長發育以及抗逆過程。類黃酮包含了黃酮、花色苷、黃烷醇、黃酮醇、黃烷酮和異黃酮等6 大類物質。據已有文獻報道,在黃酮類化合物中氧苷是最常見的糖基化類型,其中類黃酮主鏈(苷元)通過黃酮中存在的多種羥基(-OH)基團與糖基(糖供體)相連[26]。通常,黃酮、異黃酮和二氫黃酮多在7-OH 上糖基化形成糖苷鍵;二氫黃酮醇和黃酮醇則多在3-,7-,3’-,4’-OH上形成單糖苷鍵,或者會在3,7-二OH,3,4’-二OH 及7,4’-二OH 形成雙糖苷。目前在豆科植物中對異黃酮類化合物的糖基化已經研究的比較廣泛,主要集中在大豆、葛根和甘草中,大豆中GmUGT1-GmUGT7和GmUGT9均已被鑒定具有異黃酮7-O 位的糖基化活性[27]。葛根中PlUGT1、PlUGT2、PlUGT4、PlUGT13 和PlUGT15 通過克隆和功能表征,發現均可以大豆苷元及染料木素為底物進行7-O 位的糖基化。在黃蓍草中表征的PaUGT1 和PaUGT2 對多種黃酮類化合物具有7-O-糖基化的活性[28]。在植物中,UGTs 通常定位于細胞質中,具有多種生物學功能,參與植物天然產物的生物合成,如類黃酮、苯丙素、萜類和類固醇,以及植物激素的調節[29]。具備穩定天然產物(游離型苷元)、增強水溶性、改變反應性和生物活性等作用。另外還參與了生物活性天然產物的合成、貯存、轉運,植物激素和細胞穩態調節以及外源性藥物的減毒的過程,具有廣泛且重要的價值和意義[30-32]。利用糖基化可以改變和優化藥物化合物的潛在用途,豐富植物次生代謝產物的多樣性等。
鳶尾中含有豐富的異黃酮化合物及其糖苷,暗示其中存在豐富的(異)黃酮糖基轉移酶系統。鳶尾中的糖基轉移酶UGT73CD1 被報道能夠在鳶尾苷元的7-OH 位置進行糖基化產生鳶尾苷,此外該酶還能催化產生氧苷和氮苷[21]。除此之外鳶尾科植物中少有UGT 基因被研究功能。因此,本研究著重于解析挖掘到的糖基轉移酶基因ItUGT797的全貌,包括蛋白結構在內的各項生物信息學指標以及基因的進化關系均被完善分析。QPCR 結果顯示從鳶尾轉錄組中篩選出的糖基轉移酶基因ItUGT797在花中表達最高,并且可能是7-O-UGT。鳶尾的花呈現青紫色,富含花青素這一類黃酮化合物,推測ItUGT797可能參與花中包括花青素在內等類黃酮化合物的生物合成中的糖基化修飾。具體的功能驗證需要后期進行體外酶活實驗確定。且ItUGT797在根狀莖和葉中也有表達,鳶尾苷和鳶尾苷元主要分布于根狀莖部位,這暗示著它也有可能參與根狀莖中鳶尾苷的合成。已有研究表明,參與有效活性成分生物合成的基因的表達模式與次生代謝物的積累模式類似[23,33-34]。例如,常見藥用植物丹參的藥理活性成分丹參酮以及丹酚酸主要在根部積累,而許多調控這兩種化學成分生物合成的轉錄因子也在根中高表達[24-25,35]。在除此之外,對于植物體而言,異黃酮類化合物參與植物生長發育和抵抗脅迫的過程,通過提高植物對環境紫外的抵抗和對病菌和昆蟲的抵御能力來增強自身的保護[36-37]。因此,ItUGT797作為可能參與異黃酮類化合物糖基化修飾的成員,在植物生物發育、次生代謝以及防御抗逆過程中的功能值得進一步發掘研究。
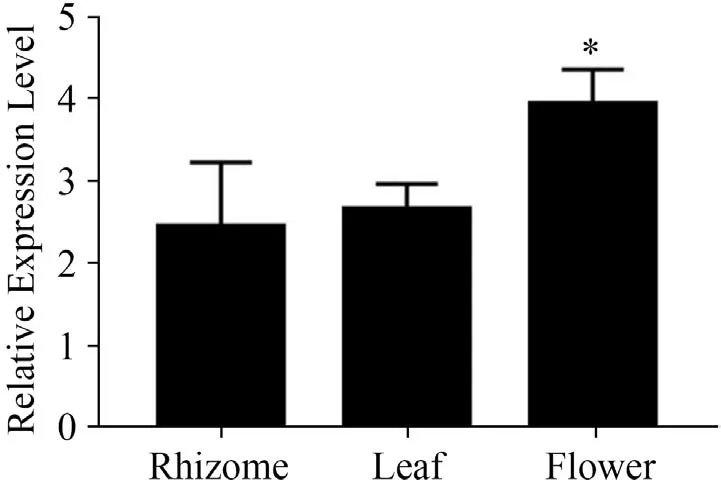
圖10 鳶尾ItUGT797基因在不同部位的表達情況
4 總結以及展望
鳶尾作為著名的藥用和園藝植物,市場需求極大。其主要活性成分為鳶尾苷、鳶尾新苷B、野鳶尾苷等多種異黃酮類化合物[3]。異黃酮糖苷類化合物的生物合成依賴于糖基轉移酶(UGTs)的修飾。本研究從鳶尾轉錄組中鑒定出ItUGT797這一糖基轉移酶基因,網站預測結果表明其為穩定的膜外非分泌蛋白,且主要由無規則卷曲組成;ItUGT797在葉片中表達最高。使用大腸桿菌為媒介的原核表達結果表明該基因可以表達出較少的可溶性蛋白。系統發育進化樹結果暗示ItUGT797 可能是一類主要以類黃酮為底物進行催化過程的糖基轉移酶。已有很多物種的糖基轉移酶被驗證可以提高植物有效活性成分含量和參與抵御生物脅迫[38-39]。然而,關于鳶尾中糖基轉移酶的研究較少,本研究后續將結合體外酶活實驗驗證ItUGT797的功能。挖掘鳶尾中的UGTs 既為其可能參與植物生長發育、抗逆等解析了機制也為實現天然藥物的生物合成提供更多工具酶,最終達到提高藥用植物品質的目的。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13