黃花蒿Dof基因家族鑒定及在GA-UV處理下對青蒿素生物合成的影響*
2022-09-29 15:19:38王星文馬婷玉尹青崗師玉華
世界科學技術-中醫藥現代化 2022年5期
關鍵詞:分析
王星文,鄔 蘭,馬婷玉,尹青崗,師玉華,向 麗
(中藥鑒定與安全性評估重點實驗室/中國中醫科學院中藥研究所 北京 100700)
Dof(DNA binding with one finger)家族是一類植物特有的轉錄因子家族,可通過與啟動子結合或者與特定蛋白相互作用來調控基因表達,參與多種植物生理過程的調控[1]。Dof蛋白N-末端共有一個高度保守的,由52 個氨基酸組成的Dof 結構域,其中核心基序CX2CX21CX2C與Zn2+共價結合形成的單鋅指結構,可以特異性的識別并結合下游基因啟動子序列中核心基序為“T/AAAAG”的元件,但也有如南瓜AOBP基因,僅可以與“AGTA”核心的基序結合;Dof蛋白C-端的轉錄調控結構域保守性低,氨基酸序列多變,主要作用是蛋白-蛋白結合互作,因此Dof家族在參與調控的途徑上存在較大差異,功能也具有多樣性[2-4]。自第一個Dof 蛋白ZmDof1 在玉米中被鑒定以來,Dof 家族已在擬南芥、水稻、番茄、白菜、菊花等[5-9]多種植物中被鑒定,主要參與光響應調控[10-11]、光周期調控[12-13]、植物激素和非生物脅迫信號通路傳導[14-15]、糖代謝[16]、氮代謝[17]、種子發育[18]和細胞周期調控[19]等復雜植物生理過程。如Xu 等研究發現擬南芥Dof 家族基因AtOBP4(OBF BINDING PROTEIN 4)參與調控細胞周期進程,可以抑制細胞的生長和增殖,致使植株生長出現矮化、少花等明顯缺陷[19]。另有一類Dof 家族基因AtCDFs(CYCLING DOF FACTOR)廣泛參與到光周期調控中[13],過表達AtCDF1基因的擬南芥植株表現為早花表型[20];而在番茄中AtCDFs的同源基因SlCDFs不僅參與光周期開花調控,對植株在干旱、高鹽等非生物脅迫下的耐受性也有增強作用[12]。AtDAG1(DOF AFFECTING GERMINATION 1)突變的擬南芥種子對紅光敏感,受光敏色素B(phytochrome B,phyB)基因的調控,可降低種子萌發過程中對紅光與赤霉素(Gibberellic acid,GA)的需求量,并參與負調控光介導的下胚軸生長過程[18,21]。AtOBP3(OBF4 BINDING PROTEIN 3)受水楊酸(Salicylic acid, SA)與生長素的影響,同時也是phyB下游調控因子,且受到隱花色素1調控,其突變體在藍光下子葉生長明顯大于野生型[10]。綜上可見,Dof 家族轉錄因子是響應光和激素脅迫的重要調控因子,極大的參與到植物生理活動中。而植物代謝產物,尤其是作為藥用活性成分的次生代謝產物也多是因響應外界逆境信號而累積[22]。因此,探究轉錄水平上植物光和激素的響應調控網絡,對理解和進一步深入研究植物次生代謝產物生物合成機制尤為重要。
青蒿來源于菊科植物黃花蒿(Artemisia annua)的干燥地上部分,是我國的傳統中藥,具有清虛熱、除骨蒸等功效,其主要活性成分青蒿素及其衍生物是聯合國推薦的抗瘧一線藥物。而除治療瘧疾外,青蒿素及其衍生物在治療紅斑狼瘡等免疫性疾病方面也有突出作用[23]。黃花蒿是青蒿素提取的主要原料藥材,目前對于青蒿素的生物合成途徑也已有了許多研究,其中非生物脅迫及生物脅迫均對其具有促進作用[24-25]。課題組前期研究發現在紅光和藍光處理后,黃花蒿中青蒿素的含量明顯提升[26]。GA 和紫外線B(UV-B)處理也對青蒿素生物合成具有正向影響作用,協同促進了植株體內青蒿素的積累[22]。Dof 家族蛋白廣泛參與植物響應非生物脅迫的復雜調控過程,但在黃花蒿中還未被系統鑒定與分析。基于其家族功能進行推測,AaDof 家族極有可能通過光與激素等響應途徑影響黃花蒿體內青蒿素的生物合成。因此,本研究基于黃花蒿基因組及轉錄組數據,對黃花蒿中Dof 家族進行鑒定,并對其理化性質、系統進化發育、保守結構、GA 和UV-B 脅迫下的基因表達模式等進行分析,以期為深入探索Dof 家族功能、進一步解析青蒿素生物合成調控網絡提供參考依據。
1 材料與方法
1.1 材料
本研究所用黃花蒿種質為課題組于海南收集的野生黃花蒿品種。種子播種后在光周期16/8 h,溫度25°C 的溫室中培養4 周。以100 μmol·L-1GA 和2 W/m2UV-B 燈(275-320 nm,Huaqiang)進行脅迫處理,對照組不進行任何處理。GA、UV-B 處理及GA+UV-B協同處理6 h 后采集幼苗的所有地上部分,每10 株混合為一個重復,每個處理取三個生物學重復。材料經液氮速凍后保存于-80℃備用。
1.2 黃花蒿Dof家族鑒定及序列分析
本研究基于黃花蒿(Artemisia annua)基因組數據(PRJNA416223)。經在線網站iTAK(http://itak.feilab.net/)檢索后獲得候選AaDof 蛋白序列,通過Pfam 數據庫(http://pfam.sanger.ac.uk/search)鑒定各序列結構域模型完整性,去除重復及結構不完整序列。然后利用ExPASy Proteomic Server (http://expasy. org/tools/protparam.html)對序列蛋白質理化性質進行預測,并通過在線軟件ProtComp 9.0(http://linux1.softberry.com/berry. phtml? topic=protcomppl&group=programs&subgroup=proloc)進行亞細胞定位分析。
1.3 多序列比對及進化分析
將鑒定的AaDof 序列與擬南芥全部36 個AtDof 序列及水稻和番茄中部分同源Dof蛋白序列進行聚類分析[5,8],采用MEGA X 中CLUSTALW 工具進行多重序列比對分析,利用鄰接法(Neighbor-Joining,NJ)構建系統發育樹,校驗參數Bootstrap 重復次數設置為1000次。AaDof 序列自身以同樣方法建樹分析。以EvolView(https://www. evolgenius. info/evolview/#login)進行可視化修飾。
1.4 基因結構及蛋白保守結構分析
通過DNAMAN 對AaDof 蛋白進行序列比對分析,并 通 過MEME(http://meme-suite.org/tools/meme)和NCBI-CDD(https://www. ncbi. nlm. nih. gov/Structure/bwrpsb/bwrpsb.cgi)對AaDof 家族成員蛋白保守結構域進行檢索。使用TBtools提取基因結構數據,并進行可視化分析。
1.5 啟動子序列結合元件分析
提取基因組中AaDof基因上游2000bp 序列,提交至 PlantCARE (http://bioinformatics. psb. ugent. be/webtools/plantcare/html/)進行分析,篩選并合并響應元件標簽后應用TBtools軟件進行可視化。
1.6 基因表達模式分析與實時熒光定量PCR驗證
基因表達模式分析所用數據為課題組前期獲得的GA 和UV-B 處理前后黃花蒿轉錄組數據[22]。采用TBtools 軟件對AaDof基因在GA、UV-B 及GA+UV-B協同脅迫下的表達模式進行熱圖分析,并以GA+UVB 組和對照組的AaDof基因表達量(FPKM 值)數據差異倍數≥2.0為閾值篩選差異表達基因。
以AaActin[26]為內參基因進行實時熒光定量PCR(qRT-PCR)驗證。樣品總RNA 提取方法參照EASYspin Plus Plant RNA Kit(Aidlab Biotechnologies)試劑盒步驟,總RNA 反轉錄第一鏈cDNA 使用TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix(TransGen Biotech)試劑盒方法。qRT-PCR 引物通過Primer Premier 3.0 軟件設計(表1),并利用Primer-BLAST(http://ncbi.nlm.nih.gov./tools/primer-BLAST)進行引物序列特異性分析。qRT-PCR體系參照TransStart Green qPCR SuperMix UDG(Transgene)體系:Mix10.0 μL,正向引物0.3 μM,反向引物0.3 μM,cDNA 1.0 μL,ddH2O 8.4 μL。qRT-PCR在Rotor-GeneQ(Qiagen)儀器上進行,程序為:94℃預變性10 min;95℃、10 s,60℃、15 s,72℃、20 s,40 個循環;熔解曲線分析程序:95℃、15 s,60℃、1 min,95℃、15 s。
2 結果
2.1 黃花蒿Dof家族鑒定及理化性質分析
本研究從黃花蒿基因組數據中提取黃花蒿CDS序列,經iTAK在線網站檢索后初步獲得91條AaDof候選序列。以Pfam 數據庫進行鑒定后去除冗余,最終共鑒定出51 個AaDof 蛋白,均具有完整的zf-Dof 結構域(表2)。特征分析發現,AaDof家族蛋白氨基酸長度在115 aa-453 aa,分子量為12.8-49.4 KDa,理論等電點在4.43-10.31,親水指數均為負數,顯示為親水性蛋白,不 穩 定 系 數 除AaDof21、AaDof24、AaDof29、AaDof38 外均大于40,表明大部分AaDof 為不穩定蛋白。亞細胞定位預測結果顯示,39個基因定位于細胞核,占大多數。12 個基因定位在細胞外,分布在除C2.1外的所有亞族中,其中C3亞族的所有序列均顯示為細胞外定位,這可能與其參與調控的功能機制密切相關[27]。

表2 黃花蒿Dof家族基本信息
2.2 黃花蒿Dof家族系統進化分析
將AaDof 蛋白序列與36 個擬南芥Dof 序列及部分水稻和番茄[5,8]中報道的Dof 序列進行同源對比,并以鄰接法構建系統發育樹。聚類結果顯示AaDof序列易與自身聚為一支,具有較高的保守性,此外較多與番茄Dof 序列聚為一支,顯示二者進化更為相似。依據Lijavetzky 等[5]分類方法將AaDof 分為A、B、C1、C2.1、C2.2、C3、D1、D2 8 個亞族,并基于亞族順序進行重命名。A 亞族成員有13 個,相比擬南芥中3 個,水稻中4個,黃花蒿中A 亞族可能發生了復制事件。B 亞族成員共有7個,C1、C2.1、C2.2和C3亞族成員分別有4個、5 個、4 個和3 個,D1 和D2 亞族成員則分別有9 個和7個。同一亞族的Dof 蛋白功能具有相似性,依據進化樹聚類可預測其功能。
2.3 黃花蒿Dof家族的結構分析
多序列對比顯示AaDof 蛋白中Dof 結構域非常保守(圖2),除固定的C2-C2結構外,結構域中其他氨基酸分布也極為相似,與前人研究結果一致[8]。經CDD數據庫檢索分析,AaDof 家族除了zf-Dof 結構域外,AaDof46與AaDof49序列N-端還分別存在鋅指超家族N-端和SRPBCC 超家族C-端的部分結構域、AaDof44序列C-端存在Dimer_Tnp_hAT 結構域,其是HAT 超家族C-端二聚化結合域,在蛋白結合互作途徑上可能具有參考意義。經MEME 進行保守motif 分析共檢索到12 個motif,其中所有AaDof 序列均含有motif1,為Dof 保守結構域,其余motif 大部分都在C-端,基序多變但在亞族間相對保守,推測為不同蛋白結合域。C2.2 亞組均含有motif5,C3 亞族均含有motif4、motif7、motif8 和motif9,C1 亞族大部分共有motif12,D1 亞族大部分共有motif2 和motif4,D2 大部分共有motif3,這些保守結構使亞族間功能趨于相似。基因結構分析顯示AaDof基因存在內含子較少,基本僅含有0-2 個,AaDof46含有3 個。其中AaDof21和AaDof4中均含有一個異常的大內含子,還需通過實驗進一步驗證(圖3)。
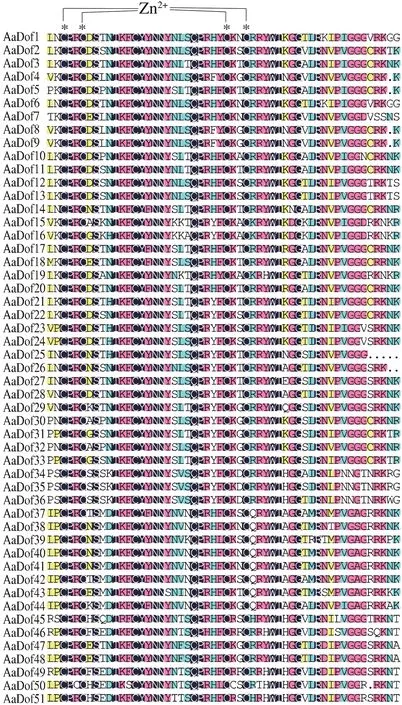
圖2 黃花蒿Dof家族保守結構域分析

圖3 黃花蒿Dof家族結構域與基因結構分析

續表

圖1 黃花蒿Dof家族蛋白系統進化分析
2.4 黃花蒿Dof家族啟動子結合元件分析
對AaDof啟動子區序列進行分析,其中AaDof12和AaDof47基因啟動子區域因基因組拼接問題不足2000 bp(圖4)。所有啟動子區域均存在SA、GA、脫落酸(Abscisic acid, ABA)、茉莉酸(Jasmonic acid, JA)等多種激素響應元件以及低溫、干旱等環境因子響應元件,尤其如D 亞族AaDof40、AaDof41等基因啟動子區域響應元件較多,可能參與到復雜的調控網絡中。A亞族、C2.1、C2.2 和C3 亞族基因的啟動子區富含如GBox(CACGTT)、GT1-motif(GGTTAA)等參與光相關調控的元件,其功能可能也與光調控相關。另外AaDof啟動子區還存在MYB轉錄因子家族結合域,細胞周期及種子發育等功能相關響應元件,有待進一步分析。本結果將為AaDof基因的功能預測提供支持。

圖4 黃花蒿Dof基因啟動子作用元件分析
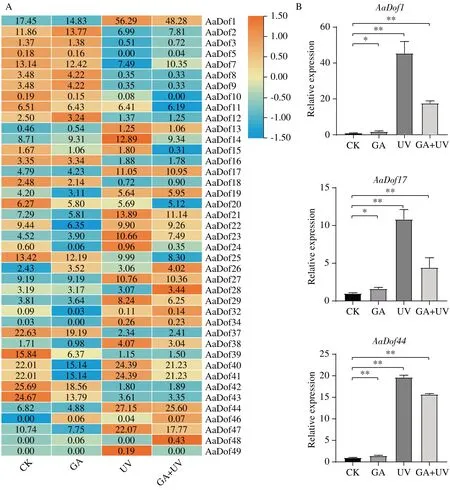
注:A:基因表達熱圖分析;B:差異基因的qRT-PCR驗證,±SD,n=3,*P<0.05,**P<0.01。
2.5 黃花蒿Dof家族基因的表達模式分析
課題組前期研究結果表明GA 與UV-B 處理都對黃花蒿中青蒿素含量具有提升作用,其中6 h 時GA+UV-B 處理作用最明顯,測得青蒿素含量最高[22]。因此基于GA、UV-B 及GA+UV-B 協同處理下6h 的轉錄組數據對AaDof的表達模式進行分析(圖5)。結果發現,41 個AaDof基因有表達且受UV-B 脅迫影響明顯。A 亞族除AaDof1和AaDof13外均在UV-B 脅迫下表達下調,C1 亞族和C2.1 亞族的大部分成員在UV-B 脅迫下表達上調。B 與D 亞族表達模式復雜,但大部分基因也都響應UV-B 調控,尤其AaDof14和AaDof49,僅在UV-B 脅迫下表達有上調。此外,部分基因也響應GA 脅迫,AaDof26、AaDof46在GA 脅迫下表達上調,AaDof15、AaDof22、AaDof40、AaDof41在GA 脅迫下表達下調,均可能密切參與到GA 激素調控網絡中。而AaDof28、AaDof48基因僅在GA+UV-B 協同處理下高表達,這些Dof基因可能參與更加復雜的調控過程。余下10 個AaDof基因未檢測到表達,其中A 亞族基因2 個,D2 亞族基因3 個。C2.2 和C3 兩亞族均只有1 個基因檢測到微量表達,分別為AaDof32和AaDof34。基于GA+UV-B 脅迫處理下表達量數據和具體FPKM 進行篩選,A 亞 族AaDof1、B2 亞 族AaDof17和D1 亞 族AaDof443 個基因表達上調倍數大于2。qRT-PCR 驗證結果發現,這3 個基因在UV-B 處理下表達量顯著上調,而在GA 處理下表達量也有上升,與轉錄組測定數據存在一定差異但趨勢一致。
3 討論
本文對黃花蒿基因組進行分析,共鑒定出51 個Dof 家族轉錄因子。研究報道擬南芥中Dof 家族基因有36 個[5]、白菜中有76 個[7]、菊花中有20 個[7],可見Dof家族在不同植物中數目差異較大。特征分析發現AaDof 家族蛋白大部分為親水不穩定蛋白。亞細胞定位預測結果則發現有23%的AaDof基因未定位在細胞核,而是定位分泌在細胞外,或可能于過氧化物酶體、葉綠體、高爾基體等細胞器中表達,這些基因的功能或許也存在特異性[27]。對Dof 家族蛋白保守結構和基因結構的分析顯示,AaDof 家族進化過程非常保守,Dof 結構域核心基序在不同物種間也極為相似。蛋白保守基序具有多樣性,但在亞族間相對一致,這也是亞族中基因功能具有相似性的基礎。此外,一些AaDof 蛋白如AaDof1、AaDof6 雖然含有保守的Dof 結構域,但是氨基酸長度小于200aa,仍需要進一步克隆驗證。根據Lijavetzky 等[5]分類方法將AaDof 家族分為A、B、C1、C2.1、C2.2、C3、D1、D2 8 個亞族,其中A 亞族與B 亞族均有一支被分出且沒有與擬南芥Dof 序列聚類,而是與水稻和番茄Dof序列相似度更高,這些基因顯示出不同的進化趨勢,在功能上可能也與原亞族有一定區別。進一步根據蛋白序列同源性對其功能進行預測,發現AaDof家族基因可能參與細胞周期、光和激素等多種調控過程中。光敏色素A(phytochrome A,phyA)和phyB基因受紅光和遠紅光誘導表達,參與調控幼苗的去黃化以及種子中GA和ABA激素調控等生理過程。擬南芥中AT1G29160.1(AtCOG1)參與phyA和phyB信號通路下游調控。D1 亞族中AaDof44 與其聚為一支,可能為黃花蒿中COG1 的同源基因并具有類似功能[11]。其余D1 亞族成員則與AtCDFs 聚為一支,可能參與到光周期及非生物脅迫相關調控中[12]。AT2G46590.1(DAG1)也為phyB下游調控因子,在種子萌發過程中起重要作用,C2.1 亞族AaDof29 與其聚為一支,因此AaDof29 功能可能與紅光響應和種子發育相 關 調 控 有 關[18,21]。 而AaDof18 與AT3G55370.1(AtOBP3)聚為一支,推測其功能與AtOBP3相似,可能也響應SA 和紅藍光,參與植株下胚軸與子葉的生長調節過程中[10]。啟動子區的響應元件分析也是預測基因功能的重要手段。通過對AaDof序列上游2000 bp的啟動子區域進行分析,發現AaDof基因可能廣泛參與到植物激素和光等調控網絡中。其中,A、C2.1、C2.2和C3亞族基因啟動子序列富含光調控元件,其亞族功能可能也與光密切相關。
植物響應外界環境信號進而調控體內次生代謝產物合成積累,Dof 家族作為光與植物激素響應調控途徑上重要轉錄因子家族,也極有可能參與到黃花蒿體內青蒿素等次生代謝產物的合成調控中。目前已證實水稻OsDof3基因參與赤霉素靶基因的調控,Yamamoto 等進一步將其鑒定為Dof蛋白P-box 結合因子(PBF),其可與bZIP 轉錄因子協同互作,調控水稻種子中蛋白儲存[28-29]。草莓中FaDOF2通過與MYB 轉錄因子FaEOBII結合參與果實中苯丙烷途徑中丁香酚含量的調控,并受到果實中ABA 和生長素水平的調節[30]。本研究通過對黃花蒿GA 和UV-B 及二者協同處理下轉錄組數據進行分析,發現大部分AaDof基因都在脅迫下表達,尤其對UV-B 處理敏感,在GA+UVB 協同處理時UV-B 處理對基因表達影響占主導。A亞族大部分成員表達量響應UV-B 脅迫而下調,C1 亞族和C2.1亞族大部分成員表達量響應UV-B脅迫而上調,這些亞族基因均參與到UV-B響應調控過程中,與啟動子序列分析結果較一致。同時也有部分基因僅響應GA 脅迫。此外,C2.2 和C3 亞族基本無表達,目前研究表明C3 亞族基因功能與植物分枝和種皮發育等有關,同時參與到擬南芥苯丙烷代謝途徑中,可以沉默類黃酮成分生物合成相關基因表達,并促進下胚軸和花蕾中羥基肉桂酸的積累[31-32],本文分析可為黃花蒿中相關通路后續研究提供參考依據。對差異基因進行qRT-PCR 驗證結果顯示AaDof1、AaDof17、AaDof44均響 應GA 和UV-B 處理,并在UV-B 脅迫下相對表達量顯著上升。綜合化學含量變化分析,這三個基因可能與黃花蒿中青蒿素生物合成有關,并起到正向調節作用。其中AaDof1 在進化分析中與AtOBP4同聚在A 亞族,功能可能與細胞周期調控相關,并可能參與到ABA 應答網絡中[33]。B 亞族AaDof17 與擬南芥PEAR(PHLOEM EARLY DOF)類蛋白序列較為相似,該類蛋白可在胞間短距離移動,功能特異性調控細胞平周分裂并參與植物形成層發育[34]。D1 亞族AaDof44 與AtCOG 聚為一支,其響應GA 和UV-B 脅迫的表達模式與AtCOG 報道功能有一定相關性。這些基因序列均與擬南芥等模式植物基因存在較大差異,推測在黃花蒿中功能更具有特異性,并可能會更多參與到黃花蒿體內萜類代謝的復雜調控中[35-36]。而在GA+UV-B 協同處理時相對表達量較UV-B 處理時有所下降的情況,可能與UV-B和GA間復雜的調控抑制關系有關[37]。
青蒿素的生物合成途徑涉及復雜的調控過程,是不同光質、JA、SA、ABA、GA 等激素共同作用的結果[38]。基于模式植物研究對一些基因在黃花蒿中同源序列功能的研究,也可能為青蒿素合成途徑的進一步解析提供理論依據。如Zhuo 等[15]研究表明AtDof2.1(AT2G28510.1)是JA 激素調控途徑的上游調控因子,可以直接結合MYC2的啟動子,影響下游基因表達。Ma 等[39]發現了黃花蒿中涉及ABA 與JA 激素途徑的兩條轉錄因子調控網絡,其下游調控因子AaTCP14/15對黃花蒿中青蒿素合成具有促進作用,而擬南芥中AtDof6被驗證可與黃花蒿AaTCP14的同源基因AtTCP14互作[40]。本文從基因組水平系統鑒定了黃花蒿Dof 家族并對其特征和表達模式進行分析,為黃花蒿Dof家族基因功能與相關轉錄調控網絡的進一步研究提供支持,并為轉錄水平上黃花蒿中青蒿素及其他次生代謝產物生物合成調控機制的解析提供參考。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06